










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkArchivos Venezolanos de Farmacología y Terapéutica
versión impresa ISSN 0798-0264
AVFT v.26 n.2 Caracas 2007
Biología molecular de los transportadores de glucosa: clasificación, estructura y distribución
Valmore Bermúdez, Fernando Bermúdez, Nailet Arraiz, Elliuz Leal, Sergia Linares, Edgardo Mengual, Lisney Valdelamar, Moisés Rodríguez, Hamid Seyfi, Anilsa Amell, Marisol Carrillo, Carlos Silva, Alejandro Acosta, Johnny Añez, Carla Andara, Verónica Angulo, Gabriela Martins.
Universidad del Zulia. Facultad de Medicina. Centro de Investigaciones Endocrino–Metabólicas "Dr. Félix Gómez". Maracaibo, Estado Zulia. e-mail: fago@medscape.com
Recibido: 28/06/2007 Aceptado: 03/08/2007
Resumen
Hexosas como la glucosa, galactosa y fructosa cumplen funciones importantes en las células eucarióticas. Estas moléculas son incapaces de difundir directamente a través de las membranas celulares por lo que requieren proteínas transportadoras especializadas para entrar al interior celular. Dichas biomoléculas pertenecen a un grupo de transportadores constituida por 2 familias de proteínas: la familia de los Glut´s (del inglés Glucose Transporters) y la familia de los co-transportadores de sodio y glucosa.
Según la información obtenida de la secuencia de aminoácidos por medio de librerías de cADN todos poseen una estructura básica similar: 12 (Gluts) o 14 (SGLT) dominios trasmembrana. Igualmente todos parecen estar glicosilados en alguna de sus asas extracelulares.
En los últimos siete años ha habido un explosivo incremento en la información sobre estos transportadores, de hecho, hasta hace diez años solo se conocían 6 transportadores pero esta familia ha crecido rápidamente hasta llegar a 14 miembros para los Gluts y 6 miembros para los SGLT´s. El impacto de estos descubrimientos se hace notar cuando se analizan los procesos en los que se involucran estas proteínas: Control de la glicemia basal y post-prandial; mecanismos de absorción de la glucosa y fructosa en el intestino delgado; absorción de fructosa en los espermatozoides; reabsorción de glucosa a nivel tubular renal y yeyuno; maduración de la expresión de Glut´s en la mama en lactación; incorporación de glucosa al músculo durante el ejercicio; mecanismo sensor en la secreción de insulina y respuestas adaptativa del metabolismo energético durante estados de estrés, etc.
Palabras clave: Glut, Glucosa, transportador, difusión facilitada, SGLT-1, SGLT-2, Co-transportador.
Abstract
Hexoses like glucose, galactose and fructose serve as basic fuel molecules for eucaryotic cells. These molecules are unable to diffuse across cellular membranes, and require transporter proteins for entry into and exit from cells. Three distinct groups of hexose transporters have been identified and classified based on their dependence on cellular energy and its chemistry structure. Each of the transporters has different affinities for glucose and the other hexoses, which largely dictates their function. The hexose transporters are large integral membrane proteins. Based on the deduced amino acid sequences of their cloned cDNAs, they have similar structures, consisting of 12 or 14 membrane-spanning regions with cytoplasmic C-terminal and N-terminal tails. Also, they all appear to be glycosylated on one of the extracellular loops. Transport of sugars across membranes appears to result from a series of conformational changes which "flips" the transporter between alternate states with the substrate binding site either facing the extracellular or cytoplasmic side of the membrane. Transport in either direction is thus possible, depending on relative substrate concentrations on either side of the membrane. The original protein, GLUT1, was identified in molecular terms 12 years ago. In the subsequent 15 years, a family of related transporters was identified (GLUTs 1-14). The impact of these discoveries is better realized when we list a sample of the processes that utilize different members of the GLUT family: control of glycemia; insulin dependent glucose utilization; transport pathways in brain neurons and glia; mechanisms of glucose and fructose uptake in the intestinal track; reabsorption of glucose in kidney tubules and jejunum; maturation of transporters during lactation and weaning; sensing of glucose levels by the pancreas and the liver; control of glucose uptake in high fat feeding; glucose uptake in response to exercise, adaptive response of energy metabolism to cellular stress.
Key words: Glut, Glucose, transporter, facilitative diffusion, SGLT-1, SGLT-2, Co-transporter.
Introducción
Los carbohidratos son sustancias químicas orgánicas de amplia distribución en la naturaleza. En las plantas son producidos por el proceso de la fotosíntesis e incluyen a la celulosa como ejemplo de un carbohidrato estructural y al almidón como carbohidrato de almacén. En las células animales los carbohidratos - en forma de glucosa o en su polímero de almacén, el glucógeno – sirven como fuente importante de energía para las actividades vitales de la mayoría de las especies vivientes que habitan el planeta.
Para poder llevar a cabo importantes funciones como la oxidación y el almacenaje, la glucosa debe entrar al interior de la célula para incorporarse a la vía metabólica que predomine según las condiciones hormonales y energéticas del momento1. Una célula puede sobrevivir si evita que su medio interno se mezcle y establezca el equilibrio con el medio ambiente. Este semi-aislamiento es proporcionado por la membrana celular, de naturaleza lipídica y relativamente impermeable a moléculas polares como la glucosa. Es por esta característica que moléculas como los carbohidratos requieren algún elemento transportador que les permita cruzar el escollo que significa la membrana; así, la naturaleza, a través de millones de años de evolución biológica desarrolló moléculas proteicas que cruzan completamente la membrana y que son capaces de formar "poros" que comunican el exterior con el interior celular, permitiendo el flujo de moléculas como los aminoácidos, vitaminas y carbohidratos en un sentido u otro. En este orden de ideas, los transportadores para glucosa trabajan de manera coordinada con factores hormonales, receptores, y segundos mensajeros para mantener el flujo de este metabolito en condiciones normales.
Los transportadores de glucosa presentes en las células de los organismos superiores se clasifican en dos grandes familias, la Familia de los Gluts y la familia de los Co-transportadores de Sodio y Glucosa2. La familia de los Gluts (aunque quizás sea mejor referirlos como superfamilia) es muy extensa ya que hasta la fecha se conocen más de 140 miembros distribuidos entre diferentes especies de animales, plantas, bacterias y hongos, por lo que sería un enfoque simplista circunscribir estas proteínas solo a organismos superiores como nosotros.
Todos los transportadores de membrana que median la transferencia de solutos desde el espacio extracelular hacia el citosol (o viceversa) comparten varias características:
Especificidad: en general cada transportador es específico para una sola sustancia o un grupo muy emparentado de éstas.
Saturación: La capacidad de transporte puede alcanzar un máximo cuando todos los sitios de unión para el soluto a transportar están ocupados.
Competición: ocurre cuando más de una sustancia es capaz de ser transportada por el sistema transportador. En este caso, tal como ocurre en la inhibición competitiva de enzimas, el soluto que se encuentre a mayor concentración será preferentemente transportado2,3,4.
Familia de los co-transportadores de Na+/Glucosa (SGLT)
En el epitelio intestinal y epitelio de los túbulos contorneados proximal y distal existen sistemas de co-transporte de glucosa acoplados a Na+ que permiten la absorción rápida de esta molécula desde el íleo hacia el sistema portal y además de la reabsorción de la glucosa filtrada en el glomérulo nuevamente al torrente circulatorio. Este sistema se denomina SGLT (Sodium/Glucose Transporters), del cual se conocen 6 isoformas (SGTL1-6) que aprovechan el transporte del Na+ a favor de su gradiente de concentración para generar una corriente electroquímica que produce los cambios conformacionales necesarios para la traslocación de la glucosa a través de la membrana plasmática5,6,7,8.
Al comparar las estructuras primarias de estos transportadores se encuentra que el SGLT-2, SGLT-3 y SGLT-4 tienen un 70, 59 y 54 % de homología con el SGLT-1 respectivamente.
SGLT-1 (SLC5A1)
El gen del SGLT-1 (Fig. 1) se denomina SLC5-A1 y fue aislado a partir de librerías de cDNA de intestino delgado de conejo. Con una extensión de 80 Kb y 15 exones se ubica en el cromosoma 22 en la región q13.1. Su trascripto es una proteína de 664 aminoácidos y 73KDa con una estructura secundaria formada por 14 α-hélices cuyos extremos amino y carboxilo terminales se encuentran en el espacio extracelular.

Figura 1. El principal transportador de monosacáridos (glucosa, galactosa y manosa) en el intestino delgado es el SGLT-1 o Sodium-Glucose Transporter – 1. Esta es un proteína que co-transporta 2 iones Na+ y una molécula de glucosa a razón de unas 1.000 moléculas/segundo. El SGLT-1 se organiza en forma de 14 alfa-hélices que cruzan la membrana plasmática la misma cantidad de veces.
En el ser humano este transportador se expresa primariamente a nivel del íleon, el sitio fundamental de absorción de monosacáridos como la glucosa, galactosa, manosa y fructosa. Este transportador es específico para la absorción de glucosa y galactosa en las células epiteliales del ribete en cepillo6.
Las células absortivas tienen una naturaleza polar, esto es, que la expresión de las proteínas transportadoras de membrana es diferente en el polo apical (luminal) que en el polo basolateral. Este hecho permite la transferencia eficiente de los azúcares simples (monosacáridos) desde la luz intestinal hacia los capilares. Igualmente, existen diferencias importantes entre las células que se ubican hacia las criptas, ya que en la parte baja de la microvellosidad (células muy jóvenes) y aquellas que se ubican hacia el 1/3 superior de la misma (células maduras). Las células epiteliales intestinales tienen una vida corta, originándose de células madre en las criptas y madurando en la medida que migran hacia el tope de la microvellosidad, desde donde se exfolian de 2 – 5 días después de su nacimiento. Así, los niveles mas altos de proteínas transportadoras de membrana se encuentran en las células epiteliales maduras del 1/3 superior de la microvellosidad5.
En este momento se conoce bastante bien el proceso de absorción de la glucosa, galactosa y fructosa en el borde en cepillo intestinal. De hecho, gracias a la aplicación de técnicas de biología molecular y biofísica, así como por modelaje por computadoras se ha obtenido información valiosa que apoya la tesis de que la absorción de monosacáridos ocurre en 4 fases:
1. Unión de dos iones Na+ a la cara externa del transportador, lo que produce un cambio conformacional que permite el acoplamiento de 1 molécula de glucosa o galactosa.
2. Transferencia del Na+ y del monosacárido hacia la cara citoplasmática del transportador gracias a un segundo cambio conformacional ocasionado por la glucosa y que involucra la rotación y el re-arreglo de la estructura α-helicoidal del SGLT-1.
3. Una vez en la cara interna del transportador, la glucosa se disocia del mismo y pasa al citosol para luego expulsar los 2 iones Na+, restituyendo al transportador a su forma libre de ligando. La baja afinidad del sitio de unión intracelular por la glucosa y el sodio, junto con la baja concentración de intracelular de Na+ en comparación con la extracelular y luminal (5-10 meq vs. 140-145 meq) promueve esta disociación, permitiendo que el proceso de transporte ocurra 1.000 veces por segundo a 37 ºC.
4. Finalmente, ocurre un cambio de conformación que permite la fijación de otra molécula de glucosa y 2 Na+ en el lado extracelular8 (Fig. 2).

Figura 2. Mecanismo postulado para el transporte de Na+ y Glucosa por el SGLT-1. Este proceso aparentemente ocurre en 4 fases muy bien definidas: a) unión de dos iones Na+ al co-transportador (1 y 2) b) Esto produce cambios en la conformación del SGLT-1 que permite la unión de una molécula de glucosa (3). c) Luego ocurre una re-organización estructural que lleva el Na+ y la Glucosa hacia la cara citosólica del transportador (4) para finalmente d) liberar la molécula de glucosa y luego los iones Na+ hacia el citosol.
El gradiente electroquímico de Na+ entre la luz intestinal y la célula epitelial se mantienen gracias a la bomba de Na+/K+ situada en la membrana basolateral, con lo que, 3 iones Na+ se intercambian con 2 K+, lo que mantiene un circuito de circulación de sodio que impulsa la absorción eficiente de la glucosa.
Otro aspecto importante de la absorción de Na+ y glucosa es el co-transporte de agua desde la luz intestinal al interior de la célula epitelial y de allí hacia el compartimiento intravascular (Fig. 3). De hecho, el incremento en la concentración de estos dos solutos genera suficiente fuerza osmótica para impulsar el movimiento de agua hacia la célula epitelial en una magnitud de unos 9-10 litros de H2O en 24 horas, e incluso, la reabsorción de unos 180 litros de agua/día en los túbulos renales9.

Figura 3. La absorción de la glucosa en el epitelio intestinal implica el co-transporte de Na+ por el SGLT-1 y el mantenimiento del gradiente de Na+ gracias a la bomba de Na+/K+. De forma secundaria y salida de Na+ al espacio basolateral genera la suficiente fuerza osmótica para arrastrar y absorber agua hacia los capilares. la fructosa se absorbe mediante otro tipo de transportador, el Glut-5, una proteína que atraviesa la membrana 12 veces. Finalmente, el paso de la glucosa y la fructosa hacia la sangre se lleva a cabo a través del Glut-2, un transportador de baja afinidad y alta capacidad de transporte.
Existe aún controversia de cual es la vía más importante que toma el agua para ingresar al epitelio intestinal, sin embargo, hay 3 vías posibles conocidas:
1. Difusión pasiva del agua a través de la membrana de los enterocitos: Se sabe desde hace tiempo que la membrana plasmática es levemente permeable al agua, en especial, si se crea una diferencia en la concentración de solutos tal como ocurre en el co-transporte de Na+ y glucosa. Sin embargo, la magnitud de la diferencia de gradiente no es suficiente para crear una corriente osmótica suficiente que explique la absorción de 10 litros de agua por día.
2. Difusión de agua a través del SGLT-1 junto con el Na+ y glucosa (Transporte activo secundario): Investigaciones recientes han determinado que el SGLT-1 se comporta como un transportador de agua, movilizando unas 260 moléculas de agua por cada ciclo de transporte de 2 iones Na+ y cada molécula de glucosa, lo cual representa unos 5 litros de agua/día, por lo que, igual que en el caso anterior, no explica la movilidad de 10 litros de agua/día (Fig. 3).
3. Difusión de agua a través de otros co-transportadores de nutrientes (Transporte activo secundario): Otros co-transportadores como los transportadores para aminoácidos y péptidos, así como el co-transportador de Na+/yoduro y el K+/Cl- pueden, al igual que el SGLT-1 pueden transportar de 30 – 50 moléculas de agua por ciclo8.
4. Transporte de agua a través de las Acuaporinas (AQP): Se han descrito proteínas pequeñas de unos 45 kDa que contienen 6 α-hélices trasmembrana que se encargan de servir de poros para el transporte específico de agua y que recibieron el nombre de Acuaporinas. Estas proteínas son capaces de transportar las moléculas de agua a una velocidad 100 veces mayor que cualquier co-transportador de solutos. De esta forma, es atractiva la idea que a través de estos poros se absorba la mayor cantidad de agua a nivel intestinal. Hasta la fecha se conocen 13 isoformas, encontrándose las AQP 3, 7 y 8 en el intestino delgado humano. Su papel en la absorción de agua aún se encuentra bajo investigación activa, aunque es bien conocido su papel en la reabsorción de agua en el túbulo contorneado proximal en el riñón9.
SGLT-2 (SLC5A2)
El gen de este co-transportador se aisló de librerías de tejido renal humano en el cromosoma 16 p11.2 que se expresa fundamentalmente en la corteza renal y en mucho menor grado en el íleo. Este transportador es una proteína integral de membrana de 672 aminoácidos con una estructura secundaria similar al resto de los miembros de esta familia y que se encuentra en las células epiteliales del túbulo contorneado proximal, de allí, que la función principal de este co-transportador es la reabsorción de Na+, glucosa y agua a nivel renal bajo los mismos principios del SGLT-110. Sin embargo, el descubrimiento y la caracterización de la acuaporina 2 en el túbulo contorneado proximal y las acuaporinas 2 y 6 en los túbulos colectores obligarán en un futuro próximo a la revisión de los procesos de transporte de solutos y agua a través del epitelio tubular renal9.
SGLT-3 (SLC5A4 ó SAAT1)
El gen de este Co-transportador se ubica en el cromosoma 22q12.2-q12.3 y se aisló por primera vez de librerías de riñones de cerdo y posteriormente en riñones humanos, ubicándose solo a 0,10 mb corriente abajo del gen del Glut-1 con una distribución de exones e intrones similar, por lo que se cree que se originó de una duplicación de éste último. Este gen codifica una proteína de 659 aminoácidos que se ha detectado en músculo esquelético, sistema nervioso central y en neuronas de los plexos nerviosos sub-mucosos y mioentéricos a nivel de la placa motora. Este transportador tiene baja afinidad por la glucosa (50mM) así como una muy baja capacidad de transporte para la misma. Evidencia reciente indica que se comporta como glucosensor en la membrana plasmática de las neuronas colinérgicas y del tejido muscular liso y estriado, regulando de una forma aún desconocida la actividad muscular11.
SGLT-4 (SLC5A9), SGLT-5 (SLC5A10) y SGLT-6 (SLC5A11)
El SGLT-4 y el SGLT-5 fueron aislados recientemente de librerías de ADNc de intestino delgado humano en los cromosomas 1p32 y 17p11.2 respectivamente, gracias a los esfuerzos de Robert L. Strausberg del Mammalian Gene Collection (MGC) Program Team del Nacional Cancer Institute en los EUA.
El ARNm del SGLT-4 se encuentra fundamentalmente en el intestino y riñón y su transcipto posee actividad transportadora de glucosa con un Km de 2,6 mM. La manosa tiene una potente actividad inhibidora del transporte de glucosa, por lo que se cree que es capaz de transportar casi todos los monosacáridos presentes en la dieta a través del epitelio intestinal o del epitelio tubular renal. El ARNm del SGLT-5 se encuentra fundamentalmente en el intestino delgado y riñón, aún no se cuenta con datos sobre el Km y sustratos a transportar.
El gen del transportador SGLT-6 se encuentra en el cromosoma humano 16p12-p11, el cual está dividido en 16 exones generando una proteina de 675 aminoácidos con 14 dominios transmembrana, compartiendo gran homología con el SGLT-1. De forma interesante, la región del genoma donde se encuentra este transportador se relaciona con el gen responsable del síndrome de discinesia y convulsiones infantiles así como el síndrome de convulsión infantil familiar benigna10,12,13.
Transportadores de difusión facilitada para Hexosas (GLUTS)
Si se considera cualquier Glut dentro del contexto de una gran familia de proteínas puede notarse de forma inmediata que todos poseen características comunes que en términos bioquímicos se denominan "firma molecular de los transportadores de glucosa" y que no es más que un conjunto de secuencias primarias aminoacídicas extremadamente conservadas que determinan estructuras secundarias y terciarias (dominios o motifs) que son responsables de las características funcionales de la proteína: especificidad por uno o más carbohidratos, afinidad por el sustrato, distribución tisular, ubicación celular, regulación de su actividad por hormonas, etc4.
Las primeras firmas en los Gluts pueden estudiarse históricamente con el descubrimiento del primer transportador de glucosa en 1977 a partir de membranas de eritrocitos humanos por Michihiro Kasahara y Meter Hinkle de la Universidad de Cornell y el trabajo clásico llevado a cabo ocho años después en un proyecto conjunto dirigido por Mike Mueckler y Harvey F. Lodish del instituto Whitehead de investigaciones biomédicas, en el cual se dilucidó la secuencia de aminoácidos de este transportador. Los investigadores trabajaron en sentido inverso mediante una sencilla estrategia que consistió en el aislamiento del ADN que codificaba la proteina, secuenciación del número y orden de las bases que integraban ese ADN y finalmente, aplicación del código genético, con lo que se dedujo que codificaba una proteína de 492 aminoácidos que podía organizarse en 25 segmentos, 12 de los cuales eran segmentos hidrofóbicos que cruzan el espesor de la membrana plasmática denominados segmentos T (T = trasmembrana) con una estructura α helicoidal, y 13 segmentos muy hidrofílicos, llamados asas (intra o extracelulares) que conectan a los segmentos trasmembrana. Este transportador característico de los eritrocitos recibió el nombre de Glut 12 (Fig. 4).

Figura 4. La gran familia de los transportadores de glucosa GLUT´s se divide en 3 clases, sin embargo todas comparten ciertas características en común como 12 alfa-hélices trasmembrana, varios dominios de glicosilación y algunos dominios muy conservados relacionados con la traslocación de la glucosa al interior de la célula.
Se ha planteado la posibilidad que los Gluts emergieron evolutivamente por la duplicación de una proteína con seis dominios trasmembrana como lo sugiere la presencia de estructuras repetidas en ambas mitades del transportador. Estas secuencias son el motif GRR/K presente entre las asas transmembrana 2 y 3 (TM2 y TM3) y entre las asas transmembrana 8 y 9 (TM8 y TM9) y el motif EX6R/K presente entre TM4 y TM5 y entre TM10 y TM11. Recientemente se han identificado otros motifs esenciales para la actividad del transportador como el QLS del asa TM7 que determina la afinidad del transportador por su ligando. Inmediatamente después de éste se pueden apreciar dos residuos de glutamina altamente conservados. El primero (Glu282) presente en el Glut 1 se requiere para la unión del inhibidor ATB-BMPA. De igual importancia son dos residuos de triptófano (Try338,412) en el Glut 1 que también están conservados en el Glut 2, 3 y 4, que participan en los cambios conformacionales requeridos para el transporte de la glucosa.
Durante los últimos 4 años se han identificado nuevos genes que codifican proteínas transportadoras tipo TDFH para la glucosa pertenecen a la familia SLC2A (del inglés SoLute Carrier 2A). Sobre la base de la homología de la secuencia primaria, la familia de los Glut´s se puede dividir en tres subfamilias14 (Fig. 5):
a. Familia de la Clase I: Que se encuentra formada por los Glut´s 1, 2, 3 y 4.
b. Familia de la Clase II: Constituida por los Glut´s 5,7,9 y 11
c. Familia de la Clase III: Formada por los Glut´s 6, 8, 10, 12, el Glut-13 ó transportador para mioinositol (HMIT -1) y el Glut14 (3)
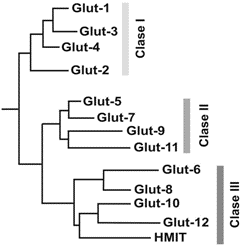
Figura 5. Dendrograma de alineamiento múltiple de todos los miembros de la familia de los Gluts. El alineamiento fue realizado mediante el programa HUSAR mostrando la relación entre cada Glut.
GLUT 1 (SLC2A1): Un Glut de alta afinidad presente en tejidos que utilizan a la glucosa como combustible principal
Este Glut también se conoce transportador de glucosa de eritrocito/cerebro cuya cinética de transporte se ha investigado desde hace más de 40 años principalmente en eritrocitos humanos cuando Widdas y colaboradores en 1952 propusieron la existencia de un mecanismo de transporte de glucosa saturable en la placenta humana. Sin embargo, era moderna del estudio de este transportador comenzó en 1977 cuando Kasahara y Hinkle lograron purificarlo en 1977 a partir de las membranas de eritrocitos, hecho que fue posible gracias a que representa el 5% del total en peso de la membrana plasmática eritrocitaria3 (Fig. 6).

Figura 6. Los Glut-1 y 3 son los transportadores con mayor afinidad por la glucosa, por este motivo se encuentran en aquellos tejidos o células que dependen exclusivamente de glucosa para sus requerimientos Energéticos tales como los eritrocitos, tejido nervioso, células embrionarias y placenta.
El Glut-1 es una proteína altamente hidrofóbica ya que el 60% de sus residuos de aminoácidos son hidrofóbicos, lo cual es consistente con el hecho de ser una proteína trasmembrana que cuenta con la organización secundaria de todos los Gluts: Doce alfa-hélices trasmembrana con asas extra e intracelulares que unen dichas alfa-hélices cuyos grupos amino y carboxiloterminal se encuentran orientados hacia el citosol. Es importante señalar que los aminoácidos más conservados entre los diferentes Glut´s del humano se encuentran en las 12 alfa-hélices y las mayores divergencias se han encontrado en el asa intracelular que conecta las alfa hélices 6 y 7 así como en los dominios amino y carboxilo terminal.
El gen del Glut-1 se ubica en el cromosoma 1p35.31.3 y cuyo cDNA codifica una proteína de 492 aminoácidos con un peso molecular de 54,2 kDa. En adición a esto se han determinado las secuencias polinucleotídicas de este transportador en ratones, ratas, conejos y cerdos, lo que ha revelado una muy remarcable similitud entre especies de alrededor del 97%15,16.
El Glut-1 parece ser el transportador de glucosa más ampliamente distribuido en el ser humano. Este se expresa en numerosos tejidos fetales y adultos como los eritrocitos, células endoteliales, células nerviosas, placenta, glóbulos blancos, células de la retina, riñón (mesangio), tejido adiposo, etc16.
Este Glut posee una alta afinidad por la glucosa (Km = 1-2 mM) por lo que es capaz de transportarla al interior de las células prácticamente a cualquier concentración, por lo que se considera como un transportador basal de glucosa que mantiene su concentración intracelular estable, hecho de gran importancia en aquellas células que requieren un suministro constante de la misma para la producción de energía, tal como sucede en tejido nervioso y eritrocito.
Al analizar la posible estructura secundaria de este Glut mediante programas de simulación se ha predicho que este posee un 82% de su estructura como alfa-hélices, un 10 % como giros beta, y un 8% de enrollamientos al azar. La orientación de las 12 alfa-hélices dentro de la membrana plasmática y la posición relativa de cada una de ellas ha sido propuesta por Mueckler y cols, la cual presenta un canal acuoso central formado por la yuxtaposición de 5 de las 12 alfa hélices: 3,5,7,10 y 11, cada una de las cuales posee carácter antipático. Sin embargo, en un modelo alternativo derivado de estudios de mutagénesis de cisteína en las alfa-hélices 2, 5, 10 y 11 se ha demostrado que todas tienen residuos accesibles a moléculas de agua capaces de formar un canal capaz de transportar glucosa y agua. Estudios posteriores han demostrado que la hélice 7 es la más sensible a los estudios de mutagénesis de cisteína lo que sugiere que se encuentra mas centralmente posicionada a la hora de formar el poro de entrada de la glucosa. La confirmación de este modelo requiere determinaciones más directas en lo que concierne a las distancias inter-hélices.
Las propiedades cinéticas de este Glut han sido extensamente estudiadas pero todavía resulta un foco de polémica la manera de cómo maneja las moléculas de glucosa de un lado a otro de la membrana. Según la propuesta más aceptada, el transporte facilitado de glucosa ocurre mediante cambios conformacionales en la estructura terciaria de transportador inducidos por la unión de la molécula de glucosa a un sitio de unión para ésta en la cara extracelular del transportador y su movimiento progresivo hacia la cara intracelular del mismo donde existe otro sitio de unión para la glucosa. Según este modelo, ambos sitios no pueden estar ocupados al mismo tiempo, por lo que debe transportarse una sola molécula de glucosa a la vez. Inconsistente con este modelo es el hecho de que aparentemente ambos sitios de unión pueden estar ocupados al mismo tiempo, por lo que una propuesta alternativa ha sido postulada por Carruthers y cols en el cual el Glut 1 existe como un homotetrámero que presentan interacciones cooperativas positivas entre sí formando dos dímeros funcionales: dentro de cada dímero, un monómero posee su sitio de extracelular unión a la glucosa libre mientras que el otro monómero tiene su sitio de unión a la glucosa intracelular libre. Al mismo tiempo, el otro dímero tiene una conformación opuesta, lo que permite cambios recíprocos que permiten la entrada de la glucosa al interior celular. Este modelo explica la mayoría de los datos cinéticos que se encuentran en conflicto con el modelo de transporte original3.
GLUT2 (SLC2A2): Un Glut con función glucosensora
El Glut-2 es un transportador de glucosa de baja afinidad (Km = 15–20 mM) que se expresa en el hígado humano adulto, riñón, células beta de los islotes de Langerhans y en la membrana basolateral de las células epiteliales del intestino delgado. Su gen se ubica en el cromosoma 3q26.1-26.3 y posee una extensión de 186,9 MB17.
Gracias a su elevado Km este Glut transporta glucosa proporcionalmente a su concentración por lo que se le atribuye la propiedad de glucosensor en las células que lo poseen, en especial en hígado y célula beta pancreática; así, por ejemplo, con una baja concentración de glucosa en plasma este Glut no es capaz de transportar glucosa al interior de la célula beta y por ende, la secreción de insulina es muy baja. Sin embargo, después de las comidas, cuando se incrementa la concentración plasmática de glucosa en suficiente magnitud para poder ser transportada al interior de la célula beta, la generación de ATP producto del metabolismo de la glucosa es capaz de estimular la liberación de insulina. En este caso, el Glut-2 actúa como un regulador que solo permite la entrada de glucosa cuando está lo suficiente elevada en plasma como para requerir la liberación de una cantidad significativamente importante de insulina18.
Otro caso interesante es la intervención del Glut-2 en el metabolismo hepático de la glucosa. Después de las comidas, el hígado es capaz de incorporar la glucosa proveniente de los alimentos gracias al Glut-2 para ser convertida rápidamente en glucógeno. De forma inversa, durante el período post-pandrial tardío (período comprendido de 6 a 8 horas después de las comidas) el glucógeno sufre degradación generando moléculas de glucosa que salen de la célula hepática a la sangre, manteniendo así los niveles de glucosa plasmática dentro de límites normales. De esta forma, es fácil notar que el Glut-2 es un transportador de tipo bidireccional que puede transportar glucosa desde la sangre al tejido o desde el tejido hacia la sangre, hecho particularmente cierto a nivel hepático y renal funcionando como sensor de la concentración plasmática de glucosa y permitiendo su intercambio entre la sangre y el hepatocito dependiendo de la condición alimentaria predominante en el momento19.
Recientemente se ha descubierto que el Glut-2 tiene también la habilidad de transportar fructosa gracias a un motif denominado HVA en el alfa-hélice Nº 7 que también está presente en el Glut-5 (transportador de fructosa clásico) y sustituye al motif QLS que se observa normalmente en los Gluts de alta afinidad por la glucosa como el Glut-1.
GLUT 3 (SLC2A3): El Glut de más alta afinidad por la glucosa
El Glut-3 es un transportador de glucosa de alta afinidad (Km = 1-2 mM) que fue caracterizado primariamente en cerebro. Bajos niveles de Glut-3 se han detectado en miocardio fetal y adulto, placenta, hígado y músculo. La presencia de este transportador co-agregado con el Glut-1 en tejido nervioso habla a favor de que este transportador tenga funciones de mantenimiento del nivel basal de glucosa en neuronas y placenta20 (Fig. 6). Recientemente se ha comprobado su expresión en las células de trofoectodermo de embriones de ratón. El bloqueo de la expresión de este Glut conlleva a la muerte por apoptosis del embrión comprobando la importancia de este transportador en el desarrollo embrionario.
GLUT 4(SLC2A4): Un Glut con gran movilidad
El Glut-4 es un transportador de alta afinidad para la glucosa (Km = 5 mM) que se expresa fundamentalmente en tejido muscular estriado, tejido muscular cardíaco y adipocito (Fig. 7). Su gen se ubica en el cromosoma 17p13 y tiene una extensión de 8,4 MB21,22,23,24. Este transportador no se expresa en tejidos embrionarios (ni pre ni post-implantación) y es único en el sentido de la regulación de su localización en el citosol o en la membrana por la insulina25. En condiciones basales, la vasta mayoría de las moléculas de Glut-4 se encuentran localizadas dentro de vesículas en el citosol que forman dos tipos de compartimientos bien definidos, ya que un grupo de estas vesículas responden a la señal de la insulina y otro grupo responde fundamentalmente al estímulo que representa la actividad física. Este comportamiento representa un mecanismo muy fino de regulación del metabolismo de la glucosa que solo permite la entrada de glucosa al tejido muscular cuando es lo suficientemente elevada como para estimular la secreción de insulina y que en última instancia favorecerá la entrada del excedente de glucosa al interior muscular. Cuando el nivel de glucosa en plasma es bajo la liberación de insulina también lo es, por lo que durante el ayuno la mayoría de las moléculas de glucosa son derivadas a los tejidos que tienen Glut´s de alta afinidad (Glut-1 y Glut-3) que no son sensibles a la presencia de la insulina. De esta forma, durante el ayuno el músculo es incapaz de incorporar glucosa en cantidades significativas (a menos que se haga actividad física enérgica) preservándola para el tejido nervioso y eritrocito26,27,28,29,30.

Figura 7. Micrografías por fluorescencia con anticuerpos contra Glut-4 de un adipocito antes (A) y después (B) de la estimulación con Insulina. En este tipo de tinción los anticuerpos fluorescentes se unen al Glut-4 pudiéndose notar el cambio en la distribución de los transportadores sin insulina y con la estimulación de la hormona. Puede observarse claramente como en ausencia de insulina los Glut-4 se encuentran dispersos en el citosol (A). La estimulación con insulina produce una dramática migración de los Glut-4 hacia la membrana plasmática y por lo tanto un incremento en el transporte de glucosa al interior celular.
Uno de los hitos más importantes en la bioquímica moderna ha sido la dilucidación de los mecanismos involucrados en la respuesta de las vesículas que contienen Glut-4 a la señal insulínica y que finalmente conducen a la fusión de éstas con la membrana plasmática. Este intrincado sistema requiere el concurso de una serie de proteínas de las vesículas y de la membrana que se denominan en conjunto "proteínas Snare". La traslocación del Glut-4 a la membrana también requiere de la activación de la enzima fosfatidilinisitol-3-cinasa (PI-3K) por intermedio del IRS-1 fosforilado, que forma un complejo con dicha enzima que produce un incremento de su actividad unas 20 veces. Igualmente, una segunda vía de activación de la traslocación (por ejercicio) se lleva a cabo gracias a la activación de la enzima AMPK por el incremento de la relación AMP/ATP y por alosterismo positivo por el AMP31,32,33. Existe evidencia reciente de que una tercera vía de traslocación de Glut-4 a la membrana que involucra la síntesis de óxido nítrico (NO) durante la contracción muscular y activación ulterior de la enzima guanilato ciclasa, ya que en experimentos utilizando Nitroprusiato de Na+(donador de NO) se observó un incremento en el transporte de glucosa en células de músculo esquelético aislado.
GLUT 5 (SLC2A5): Un Glut específico para la Fructosa
El Glut-5 es un transportador específico para fructosa (Km = 10-13 mM) que se expresa fundamentalmente en la células del ribete en cepillo del intestino delgado donde media el paso de la fructosa desde el lumen a la célula epitelial intestinal. Bajos niveles de este transportador también se encuentran en eritrocitos, riñón, espermatozoides, músculo esquelético y tejido adiposo de humanos y ratas (34). Su expresión en el músculo esquelético humano se relaciona a su capacidad de utilizar la fructosa para la glucólisis y la síntesis de glucógeno de forma independiente de la incorporación por medio del Glut-1 y el Glut-4. Este transportador no posee uno de los dominios de reconocimiento de la glucosa, el dominio QLS, en la alfa hélice Nº 7.
Más allá del mecanismo de absorción de fructosa en intestino, mucha polémica ha generado el mecanismo de obtención de energía de los espermatozoides humanos. A pesar que los datos disponibles en la actualidad son consistentes con el concepto de que estas células usan fundamentalmente a la glucosa como fuente de energía, es un hecho cierto que la concentración de glucosa en los túbulos seminíferos es muy baja y que la concentración de ésta en el líquido seminal es muy variable, desde un nivel muy cercano a cero hasta 90 mg/dl. La expresión de varios transportadores de hexosas con diferentes afinidades puede representar una adaptación funcional para transportar diferentes sustratos energéticos cuya concentración varía durante el ciclo de vida de los espermatozoides, de hecho, estudios previos han concluido que en el espermatozoide humano se expresan también los Glut´s 1,2 y 34.
GLUT 6 (SLC2A6): Redefiniendo la clasificación
Toshiaki Kayano y cols. del Instituto Howard Hughes de la Universidad de Chicago aislaron y caracterizaron en 1990 lo que pareció ser el sexto transportador de glucosa de la familia de los Glut´s con un 79,6 % de homología con el Glut 3 y con una ubicación en el brazo largo del cromosoma 5. Sin embargo, estos autores y otros confirmaron que este gen de 3,4 kb de extensión poseía un gran número de codones de interrupción de la transcripción por lo que en condiciones normales no era capaz de producir una proteína (Pseudogen). De esta manera, el lugar del Glut 6 permaneció vacío hasta el descubrimiento de una "primera versión del Glut 9" que rápidamente fue reclasificado como Glut 6 por el Comité de Nomenclatura de Genes HUGO (Human Genome Organizationman Genome Organization)35.
El análisis de la estructura primaria revela la presencia de 12 α hélices transmembrana además de contener todos los dominios que son característicos de los transportadores de glucosa, en particular los dominios PESPR/PETKGR (después de las hélices 6 y 12), el dominio GRR es las asas 2 y 8, residuos de Glutamato y Arginina en las asas intracelulares 4 y 10, 2 residuos de Triptófano (Trp 388 y 412), residuos de tirosina en las hélices 4 y 7 y Glutaminas en las hélices 5 y 7 que también se encuentran en el Glut 14.
El gen de este transportador se encuentra en el cromosoma 9 (9q34) y tienen una longitud aproximada de 8kb y consiste en 10 exones separados por intrones cortos. Mediante análisis por Northern blot el ARNm del Glut 9 se ha encontrado exclusivamente en cerebro y tejido linfoideo (bazo y leucocitos periféricos).
La proteína con mayor similaridad con el Glut 6 es el transportador Glut 8 (44,8% de homología). El Glut 6 y el Glut 8 se encuentran en una rama separada de la Clase III de la familia de los Glut´s (Fig. 2) y exhiben marcadas diferencias con los Glut´s 1 al 5. De hecho, 2 residuos de Arginina están presentes en posición 7 y 8 en el Glut 9, una región que se asocia con la especificidad del transportador por su sustrato. De manera interesante, los transportadores renales de aniones poseen estas Argininas en la misma posición del Glut 6, por lo que se ha especulado que esta proteína es un co-transportador anión/glucosa. Los parientes mas cercanos son el Glut 1 (28,5% de homología), el transportador de inositol de levaduras (26,4% de homología) y el transportador de Arabinosa y Xilosa de E. colli (28,4 y 25,7 % de homología respectivamente)3.
Respecto a la afinidad de este transportador por la glucosa se han realizado experimentos que han medido la actividad de transporte del Glut 9 mediante transfección a células COS-7 con la actividad de transporte constitutiva a una concentración de glucosa de 5 mM pero no con glucosa a una concentración de 1 mM, lo que es consistente con el hecho de que es capaz de ligar a la Citochalacina B con baja afinidad y por ende que este es un Glut de baja afinidad como en el caso del Glut 23,4.
GLUT 7 (SLC2A7): Una historia plagada de errores
Desde hace tiempo se sabe que la fracción microsomal de hígados de rata y humanos debe existir algún tipo de transportador para Hexosas con un alto Km que debería permitir que la glucosa generada de la actividad enzimática de la Glucosa-6-fosfatasa en el retículo endoplásmico liso pueda alcanzar el citosol. De esta manera, un grupo de investigadores de la Universidad de Dundee, Escocia, se dieron a la tarea de analizar posibles secuencias génicas capaces de codificar dicho transportador, lográndose finalmente el aislamiento de un nuevo cDNA en hígado de rata que mostraba gran similaridad con las secuencias de los Glut´s1-6, pero haciéndose notar que dicha homología era mayor para el Glut 2. Para este momento se decidió que este cDNA correspondía al elusivo transportador Glut 7.
Posteriormente, este mismo grupo quiso ampliar los conocimientos iniciales que habían reportado en 1992 orientando la investigación hacia la regulación de la expresión del mismo en seres humanos. Primeramente, ensayaron clonar el equivalente de este Glut mediante el uso de librerías de cDNA usando las sondas apropiadas y preparadas según la supuesta homología con el Glut 7 de rata. Mediante este método se aislaron múltiples clones, pero de los Gluts conocidos hasta ahora, pero se falló en aislar el Glut 7 humano. Posteriormente usaron el método de la transcriptasa reversa-PCR para aislar porciones del gen del Glut 7 usando las sondas apropiadas preparadas sobre las secuencias del Glut 7 de ratas. Este método también falló. Los investigadores concluyeron que a la luz de sus últimas investigaciones que ni los hepatocitos de rata ni el del humano contienen ARNm equivalente al que se clonó de los experimentos iniciales, por lo que se infiere que el hallazgo inicial del Glut 7 fue un artefacto de las técnicas de clonaje usadas en la época4.
Sin embargo, investigaciones muy recientes del Instituto de Nutrición Humana de Alemania (DIfE) han detectado un gen con características similares a la de los Gluts adyacente al gen del Glut 5 en el cromosoma 1p36.2 con el cual tiene gran similitud (58% de homología). Este gen se ha denominado provisionalmente SLC2A7, aunque aún no se conoce el patrón de expresión de este gen ni la especificidad de su transcripto por sustratos.
GLUT 8 (SLC2A8): La carrera por descubrir nuevos Gluts se ha iniciado
Dos líneas de evidencia sugieren la posibilidad de la existencia de nuevos transportadores de glucosa. En primer lugar, en algunos tejidos se han encontrado muy bajos niveles de ARNm de las isoformas conocidas de estos transportadores. En segundo lugar, experimentos llevados a cabo en ratones que no expresan el Glut 4 (Glut-4 Knockout mice) han demostrado que la capacidad de transporte de glucosa en el músculo se conserva casi normal sin observarse un incremento compensatorio en la expresión del Glut 1 ó Glut 3.
Recientemente se ha desarrollado un nuevo abordaje para la identificación de nuevos Gluts mediante la utilización de regiones extremadamente conservadas de los genes que sirven como plantillas que pueden ser comparadas con regiones de todo el genoma humano e intentar localizar regiones que tengan secuencias parecidas a estas regiones conservadas. Este abordaje llevó a la identificación y caracterización del Glut 8 y el Glut 9 (actualmente reclasificado como Glut 6)3,4.
La búsqueda en las bases de datos genómicas llevó a la identificación en el año 1999 de una secuencia característica a la de los Gluts cuyo cDNA fue clonado y expresado por dos grupos de investigadores en el año 2000: Doege y cols., en humanos e Iberson y cols., en ratones, y que en la actualidad se conoce como Glut 8. El hecho de que dos grupos de trabajo aislaran esta proteína de forma simultánea causó cierta confusión, ya que inicialmente este transportador fue denominado como GLUT X1 por uno de estos grupos y Glut 8 por el otro. En la actualidad de acuerdo al comité de nomenclatura de genes HUGO se reserva el nombre de Glut-8 para esta proteína36.
El cADN del gen (ubicado en el cromosoma 9) del Glut 8 humano y de ratón contiene franjas de lectura abierta que codifican una proteina de 477 aminoácidos. Esta secuencia tiene una homología del 29,4% con el Glut 1 y 73 de los 132 (55%) residuos de aminoácidos idénticos en todas las isoformas de Gluts de mamíferos están presentes en el Glut 8. La secuencia primaria de este transportador contiene todos los dominios característicos los Gluts, con la muy interesante carencia de los sitios de glicosilación en el asa 1, el asa E9 es mucho mas larga que en los otros Gluts (y que además contiene sitios de glicosilación) y finalmente que el dominio STS (dominio muy conservado en todos los Gluts) es reemplazado por AET37.
El transportador mas parecido al Glut 8 es el Glut 6 con el cual comparte una homología del 43,6 %, de hecho, como se dijo anteriormente estos dos transportadores pertenecen a la familia III de los transportadores de glucosa y solo se encuentran separados por 5 MB en el mismo cromosoma. Otros parientes cercanos son el Glut 1 (29,4%), el transportador de inositol del Saccharomyces pombe (30,2%) y los transportadores de xilosa (32,8%) y Arabinosa (29,1%) de la E. colli. El alineamiento de las secuencias aminoacídicas y su análisis con el programa PALING ha indicado que la homología del Glut 8 con el transportador de Arabinosa de E. colli (150 aminoácidos idénticos) es mayor que cuando se compara las secuencias del Glut 1 con el mismo transportador de Arabinosa (140 aminoácidos idénticos), por lo que se cree que el Glut 8 es el transportador mas primitivo en mamíferos hasta ahora descubierto3.
Mediante análisis de Northern Blot el transcripto del gen del Glut 8 de 2,4 Kb se ha encontrado de manera predominante en testículos, blastocisto y cerebro (cerebelo e hipocampo) y en mucha menor cantidad en el bazo, próstata, intestino delgado, corazón, cerebro y músculo esquelético. Estudios de afinidad de este transportador llevados a cabo para la 2-desoxiglucosa arrojaron un Km de 2,4 mM, la cual es bastante similar a la del Glut 3, la más alta afinidad conocida para un Glut. En experimentos realizados en ovocitos de Xenopus laevis se encontró que el transporte de glucosa fue inhibido competitivamente por fructosa3,38.
GLUT 9 (SLC2A9): El verdadero Glut 9
Cuando quedó vacío el espacio del Glut 6 en la clasificación (por ser un pseudogen) el lugar fue cubierto por el primer Glut-9 en ser descrito, por lo que quedó un puesto vacante para ser llenado por el que en definitiva sería el verdadero Glut 9. Para que este evento ocurriese no iba a pasar mucho tiempo, ya que en el año 1999 Phay y cols., descubrieron una secuencia genómica con gran homología con el Glut 5. Más importante fue el hecho de que estos investigadores patentaron esta secuencia en los E.U.A39.
Según el alineamiento múltiple de la secuencia de aminoácidos deducida del ADNc el Glut 9 es un transportador perteneciente a la Clase II con una homología de un 55 % con el Glut 5, con el que comparte la pérdida del aminoácido Triptófano en la hélice 10 (el cual se conserva en los transportadores de la Clase I). Otra característica particular de este transportador es la presencia de un asa amino-terminal larga de unos 55 aminoácidos con un motif formado por dos Leucinas3.
El gen que codifica el Glut 9 se encuentra ubicado en el cromosoma 4p15.3 y se expresa fuertemente en el riñón y el hígado, con bajos niveles en intestino delgado, placenta, pulmón y leucocitos40.
GLUT 10 (SLC2A10): ¿Una pareja para el Glut-2?
Este nuevo miembro de la familia SLC2A fue identificado en el año 2000 por Dawson y cols. Es una proteína de 541 aminoácidos que comparte un 35% de homología con los Glut´s humanos 1 y 8. La secuencia de aminoácidos del Glut 10 es casi idéntica en longitud al Glut 9, pero hasta la fecha es el miembro de mayor longitud de la familia. El Glut 10 de ratón comparte un 77,3% de homología con su contraparte humana41. Cuando este transportador se expresa en ovocitos de Xenopus laevis exhibe la capacidad de transportar la 2-desoxi-glucosa que puede ser inhibida por la droga Phloricin. El Km de este Glut es de 0,3mM. El gen del transportador se localiza en el cromosoma 20 (20q12-13.1 y 12 MB de extensión) una región asociada fuertemente con posibles diabetogenes.
La localización del gen y sus propiedades funcionales sugieren que el Glut 10 puede llevar a cabo funciones metabólicas de gran importancia y ser un elemento clave en el desarrollo de diabetes mellitus tipo 242.
Mediante la técnica de Northern blotting se ha determinado la distribución tisular del Glut-10. Este transportador se encuentra en mayor concentración en el hígado (adulto y fetal) y el páncreas, músculo cardíaco, pulmón, cerebro (adulto y fetal), músculo esquelético, placenta y riñón41,42.
GLUT 11 (SLC2A11): ¿Otro transportador de fructosa?
Este es otro nuevo miembro de la Familia SLC2A aislado en el año 2001 por Sasaki y cols. Se ha determinado que el Glut 5 es el pariente más cercano de este transportador con el que comparte un 41,7% de homología. El gen del Glut 11 humano consta de 12 exones de 29 Kb de extensión que se localiza en el cromosoma 22 (22q11.2, y 20 MB de extensión)43. En seres humanos el transcripto de este gen es de 7,2 Kb de longitud y se ha conseguido solo en músculo esquelético y cardíaco y del cual se han descrito 3 tipos de variantes: La primera es causada por la existencia de tres exones de inicio diferentes (SLC2A a, b y c). La segunda variación se debe a un salto en la transcripción del primer exón 6 y la tercera debido a una secuencia prematura de terminación entre los exones 8 y 9. Las dos últimas variaciones representan formas truncadas de los transcriptos sin significado biológico hasta el momento44. La transfección de células COS-7 con cADN del Glut 11 ha demostrado aumentar la capacidad de transporte de glucosa de estas células, sin embargo, un dato de interés, es que a diferencia del Glut 4 la actividad del transporte de glucosa del Glut 11 es inhibida en gran medida por la fructosa, lo que lleva a pensar que este es un transportador para fructosa con baja afinidad para la glucosa43,44.
GLUT 12 (SLC-2A12): ¿El elusivo hermano menor del Glut-4?
Para el momento de finalizar esta publicación, un equipo de investigadores de la Universidad de Melbourne, Australia, reportó la identificación de un nuevo transportador para glucosa, el Glut 12. Este transportador fue hallado en células de cáncer de mama MCF-7 mediante cotejo por homología con el Glut 4. El cADN de este Glut codifica una proteína de 617 aminoácidos que posee las características esenciales de los Glut. Si se logra confirmar este hallazgo estaríamos en presencia del Glut de mayor tamaño45.
El grado de homología de este transportador con el Glut 4 es del 29 % y con el Glut 10 comparte alrededor del 40% de homología. Al igual que este último, el Glut 12 presenta un asa extracelular de gran tamaño entre las α hélices transmembrana 9 y 10. Estudios recientes de inmunofluorescencia han sugerido que en ausencia de insulina el Glut 12 se localiza en la región perinuclear de las células MCF-7. El inmunobloting ha puesto en evidencia la expresión del Glut 12 en músculo esquelético, tejido adiposo e intestino delgado. Este hecho ha planteado la hipótesis que este transportador representa el elusivo segundo sistema de transporte sensible a la insulina que se encuentra en células musculares, ya que su ARNm se ha encontrado en músculo así como en próstata3,45.
GLUT 13 (SLC-2A13): ¿Un transportador de mioinositol dentro de la clasificación de los Glut´s?
El Glut-13 ó transportador de H+/Inositol codifica una proteína transportadora de membrana de 629 aminoácidos con una analogía del 35 % con el Glut 6 y que se expresa fuertemente en células de la glía y en algunas neuronas con la capacidad de transportar mioinositol y glucosa cuando se encuentra a una alta concentración. El inositol y sus derivados fosforilados (Fosfoinositósidos) juegan función importante como osmolitos y como segundos mensajeros en la regulación de la exo y endocitosis de vesículas. La expresión de este transportador en ovocitos de Xenopus laevis ha demostrado que la actividad de transporte es casi exclusiva para el mioinositol y algunos de sus isómeros con una Km de 100 mM y su expresión preferencial en el S.N.C hace pensar que su principal papel esté en la regulación de estos metabolitos a nivel cerebral46.
GLUT 14 (SLC-2A14): La frontera se hace cada día más lejana
En el año 2002 Wu y cols. del instituto Burnham en La Jolla, California, U.S.A identificaron lo que representa el último miembro de transportadores de esta familia ubicado en el cromosoma 12p13.3 (con 17.1 MB de extensión), y unas 10 MB corriente arriba del gen del Glut 3 con el cual comparte un importante parecido. Hasta ahora se había creído que el Glut 14 era un Pseudogen (igual que el Glut 6 en sus principios) resultado de la duplicación del gen del Glut 3. El gen del Glut 14 posee dos formas: una corta que consiste en 10 exones y produce un transcripto de 497 aminoácidos que es similar al Glut 3 en un 94,5%. La segunda forma, llamada forma larga codifica una proteina de 520 aminoácidos que difiere de la anterior en el extremo amino-terminal. Ambas forman poseen como todos los Glut´s 12 α-hélices transmembrana y los dominios relacionados con el transporte de glucosa. Sin embargo, en contraste con el Glut 3 este transportador se expresa fundamentalmente en los testículos donde su ARNm se encuentra en una concentración 4 veces mayor que el Glut 347.
Referencias
1. Zierler, K. 1999 Whole body glucose metabolism. Am J Physiol 276: E409-26. [ Links ]
2. Barrett, M.P., Walmsley, A.R., and Gould, G.W. 1999 Structure and function of facilitative sugar transporters. Curr Opin Cell Biol 11: 496-502. [ Links ]
3. Joost HG; Thorens B. The extended GLUT-family of sugar/polyol transport facilitators: nomenclature, sequence characteristics, and potential function of its novel members. Mol Membr Biol 2001 Oct-Dec; 18(4):247-56. [ Links ]
4. Silverman M: Structure and function of hexose transporters. Annu Rev Biochem 60:757-794, 1991. [ Links ]
5. Thorens B: Facilitated glucose transporters in epithelial cells. Annu Rev Physiol 55:591-608, 1993. [ Links ]
6. Wright EM: The intestinal Na+/glucose cotransporter. Annu Rev Physio 55:575-589, 1993. [ Links ]
7. Wright, E.M., Loo, D.D., Panayotova-Heiermann, M., Hirayama, B.A., Turk, E., Eskandari, S., and Lam, J.T. 1998 Structure and function of the Na+/glucose cotransporter. Acta Physiol Scan Suppl 643: 257-64. [ Links ]
8. Kellett, G.L. 2001 The facilitated component of intestinal glucose absorption. J Physiol 531: 585-95. [ Links ]
9. Wright, E.M. 2001 Renal Na(+)-glucose cotransporter. Am J Physiol Renal Physiol 280: F10-8. [ Links ]
10. Wells RG, Mohandas TK, Hediger MA. Localization of the Na+/glucose cotransporter gene SGLT2 to human chromosome 16 close to the centromere. Genomics. 1993 Sep;17(3):787-9. [ Links ]
11. Roll P, Massacrier A, Pereira S, Robaglia-Schlupp A, Cau P, Szepetowski P. New human sodium/glucose cotransporter gene (KST1): identification, characterization, and mutation analysis in ICCA (infantile convulsions and choreoathetosis) and BFIC (benign familial infantile convulsions) families. Gene, 2002; 285(1-2):141-8. [ Links ]
12. Shigeki Tazawaa, Tokuhisa Yamatoa, Hideki Fujikuraa, Masahiro Hiratochia, Fumiaki Itoha, Masaki Tomaea, Yukiko Takemuraa, Hidetoshi Maruyamab, Tomoyasu Sugiyamac, Ai Wakamatsuc, Takao Isogaic, Masayuki Isajia. SLC5A9/SGLT4, a new Na+-dependent glucose transporter, is an essential transporter for mannose, 1,5-anhydro-D-glucitol, and fructose. Life Sciences 76 (2005) 1039–1050. [ Links ]
13. Wright E, Turk E. The sodium/glucose co-transport family SLC5. European Journal of physiology. Vol 447, 5 feb 2004. Pag 813 – 815. [ Links ]
14. Phay, J.E., Hussain, H.B., and Moley, J.F. 2000 Strategy for identification of novel glucose transporter family members by using internet-based genomic databases. Surgery 128: 946-51. [ Links ]
15. Mueckler M, Caruso C, Baldwin SA, Pánico M, Blench I, Morris HR. Sequence and structure of a human glucose transporter. Science. 1985; 229: 941-5. [ Links ]
16. Fukumoto H, Seino S, Imura H, Seino Y, Eddy RL, Fukushima Y, Byers MG, Shows TB, Bell GI. 1998. Sequence, tissue distribution, and chromosomal localization of mRNA enconding a human glucose transporter – like protein. Proc Natl Acad Sci USA 85: 5434 - 5438. [ Links ]
17. Thorens B, Sarkar HK, Kaback HR, Lodish HF 1988 Cloning and functional expression in bacteria of a novel glucose transporter present in liver, intestine, kidney and beta pancreatic islet cells. Cell 55:281-290. [ Links ]
18. Johnson JH, Newgard CB, Milburn JL, Lodish HF, Thorens B 1990 The high Km glucose transporter of islets of Langerhans is functionally similar to the low affinity transporter of liver and has an identical primary sequence. J Biol Chem 265:6548-6551. [ Links ]
19. Matschinsky FM 1990 Glucokinase as glucose sensor and metabolic signal generator in pancreatic beta-cells and hepatocytes. Diabetes 39647-652. [ Links ]
20. GJ Mantych, DE James, HD Chung, and SU Devaskar Cellular localization and characterization of Glut 3 glucose transporter isoform in human brain Endocrinology, Sep 1992; 131: 1270 – 1278. [ Links ]
21. Katz, E.B., A.E. Stenbit, K. Hatton, R. DePinho and M.J. Charron. Cardiac and adipose tissue abnormalities but not diabetes in GLUT4 deficient mice. Nature 377:151-155 (1995). [ Links ]
22. Tsao, T.-S., R. Burcelin, E.B. Katz, L. Huang and M.J. Charron. Enhanced insulin action due to targeted GLUT4 overexpression exclusively in muscle. Diabetes 45:28-36 (1996). [ Links ]
23. Stenbit, A.E., R. Burcelin, E.B. Katz, T.-S. Tsao, N. Gautier, M.J. Charron and Y. Le Marchand-Brustel. Diverse effects of Glut4 ablation on glucose uptake and glycogen synthesis in red and white skeletal muscle. J. Clin. Invest. 98:629-634 (1996). [ Links ]
24. Xing, A.Y., J.C. Challier, M. Cauzac, M.J. Charron, J. Girard and S. Hauguel De Mouzon. Unexpected expression of GLUT4 glucose transporter in villous stromal cells of human placenta. J. Clin. Emb. & Med. 83:4097-4101 (1998). [ Links ]
25. Shepherd PR, Kahn BB: Glucose transporters and insulin action. New Eng J Med 341:248-257, 1999. [ Links ]
26. Tsao, T.-S., A.E. Stenbit, J. Li, K.L. Houseknecht, J.R. Zierath, E.B. Katz and M.J. Charron. Muscle-specific transgenic complementation of GLUT4-deficient mice: effects on glucose but not lipid metabolism. J. Clin. Invest.100:671-677 (1997). [ Links ]
27. Stenbit, A.E., T.-S. Tsao, J. Li, R. Burcelin, D. Geenen, S.M. Factor, K.L. Houseknecht, E.B. Katz and M.J. Charron. GLUT4 heterozygous knockout mice develop muscle insulin resistance and diabetes. Nature Medicine 3:1096-1101 (1997). [ Links ]
28. Charron, M.J., E.B. Katz and J.R. Zierath. Metabolic and molecular consequences of modifying GLUT4 expression in skeletal muscle. Bioc. Soc. Trans. 25:963-968 (1997). [ Links ]
29. Rossetti, L., A.E. Stenbit, L. Cheng, N. Barzali, E.B. Katz and M.J. Charron. Peripheral but not hepatic insulin resistance in mice with one disrupted allele of the glucose transporter type 4 (GLUT4 gene). J. Clin. Invest. 100:1831-1839 (1997). [ Links ]
30. Zierath, J.R., T.-S. Tsao, A.E. Stenbit, J.W. Ryder, D. Galuska and M.J. Charron. Restoration of hypoxia-stimulated glucose uptake in GLUT4-deficient muscles by muscle-specific GLUT4 transgenic complementation. J. Biol. Chem. 273:20910-20915 (1998). [ Links ]
31. Ryder, J.W., Y. Kawano, D. Galuska, R. Fahlman, H. Wallberg-Henriksson, M.J. Charron and J.R. Zierath. Post-exercise glucose uptake and glycogen synthesis in skeletal muscle from GLUT4-deficient mice. Faseb J. 13:2246-2256 (1999). [ Links ]
32. Elmendorf, J.S., and Pessin, J.E. 1999 Insulin signaling regulating the trafficking and plasma membrane fusion of GLUT4-containing intracellular vesicles. Exp Cell Res 253: 55-62. [ Links ]
33. Chiang, S.H., Baumann, C.A., Kanzaki, M., Thurmond, D.C., Watson, R.T., Neudauer, C.L., Macara, I.G., Pessin, J.E., and Saltiel, A.R. 2001 Insulin-stimulated GLUT4 translocation requires the CAP-dependent activation of TC10. Nature 410: 944-8. [ Links ]
34. Davidson NO, Hausman ML, Ifkovits CA, Buse JB, Gould GW, Burant CF. Human intestinal glucose transporter. Bell GI 1992. [ Links ]
35. Lisinski I; Schurmann A; Joost HG; Cushman SW; Al-Hasani H. Targeting of GLUT6 (formerly GLUT9) and GLUT8 in rat adipose cells. Biochem J 2001 Sep 1;358(Pt 2):517-22. [ Links ]
36. Doege H, Schurmann A, Bahrenberg G, Brauers A, Joost HG. GLUT8, a novel member of the sugar transport facilitator family with glucose transport activity. J Biol Chem 2000. 275:16275–16280. [ Links ]
37. Scheepers A, Doege H, Joost HG, Schurmann A. Mouse GLUT8: genomic organization and regulation of expression in 3T3-L1 adipocytes by glucose. Biochem Biophys Res Commun 2001. 288:969–974. [ Links ]
38. Carayannopoulos, M.O., Chi, M.M., Cui, Y., Pingsterhaus, J.M., McKnight, R.A., Mueckler, M., Devaskar, S.U., and Moley, K.H. 2000 GLUT8 is a glucose transporter responsible for insulin-stimulated glucose uptake in the blastocyst. Proc Natl Acad Sci USA 97: 7313-8. [ Links ]
39. Phay JE; Hussain HB; Moley JF. Cloning and expression analysis of a novel member of the facilitative glucose transporter family, SLC2A9 (GLUT9). Genomics 2000 Jun 1;66(2):217-20. [ Links ]
40. Doege H; Bocianski A; Joost HG; Schurmann A. Activity and genomic organization of human glucose transporter 9 (GLUT9), a novel member of the family of sugar-transport facilitators predominantly expressed in brain and leucocytes. Biochem J 2000 Sep 15;350 Pt 3:771-6. [ Links ]
41. Dawson PA; Mychaleckyj JC; Fossey SC; Mihic SJ; Craddock AL; Bowden DW. Sequence and functional analysis of GLUT10: a glucose transporter in the Type 2 diabetes-linked region of chromosome 20q12-13.1. Mol Genet Metab 2001 Sep-Oct;74(1-2):186-99. [ Links ]
42. McVie-Wylie, A.J., Lamson, D.R., and Chen, Y.T. 2001 Molecular cloning of a novel member of the GLUT family of transporters, SLC2a10 (GLUT10), localized on chromosome 20q13.1: a candidate gene for NIDDM susceptibility.Genomics 72: 1137. [ Links ]
43. Sasaki T; Minoshima S; Shiohama A; Shintani A; Shimizu A; Asakawa S; Kawasaki K; Shimizu N. Molecular cloning of a member of the facilitative glucose transporter gene family GLUT11 (SLC2A11) and identification of transcription variants. Biochem Biophys Res Commun 2001 Dec 21;289(5):1218-24. [ Links ]
44. Doege H; Bocianski A; Scheepers A; Axer H; Eckel J; Joost HG; Schurmann A . Characterization of human glucose transporter (GLUT) 11 (encoded by SLC2A11), a novel sugar-transport facilitator specifically expressed in heart and skeletal muscle. Biochem J 2001 Oct 15;359(Pt 2):443-9. [ Links ]
45. Suzanne Rogers, Maria L. Macheda, Susan E. Docherty, Maynard D. Carty, Michael A. Henderson, Walter C. Soeller, E. Michael Gibbs, David E. James, and James D. Best. Identification of a novel glucose transporter-like protein GLUT-12. 2002:282,3. [ Links ]
46. Uldry M, Ibberson M, Horisberger JD, Chatton JY, Riederer BM, Thorens B. Identification of a mammalian H(+)-myo-inositol symporter expressed predominantly in the brain.EMBO J. 2001 Aug 15;20(16):4467-77. [ Links ]
47. Wu X, Freeze HH. GLUT14, a duplicon of GLUT3, is specifically expressed in testis as alternative splice forms. Genomics. 2002 Dec; 80(6):553-7. [ Links ]
48. Baumann, C.A., Brady, M.J., and Saltiel, A.R. 2001 Activation of glycogen synthase by insulin in 3T3-L1 adipocytes involves c-Cbl-associating protein (CA))-dependent and CAP-independent signaling pathways. J Biol Chem 276: 6065-8. [ Links ]
49. Tsao, T.-S., E.B. Katz, D. Pommer and M.J. Charron. Amelioration of insulin resistance but not hyperinsulinemia in obese mice overexpressing GLUT4 selectively in skeletal muscle. Metabolism 49:340-346 (2000) [ Links ]
50. Li, J., K.L. Houseknecht, A.E. Stenbit, E.B. Katz and M.J. Charron. Reduced glucose uptake precedes insulin signaling defects in adipocytes from heterozygous GLUT4 (+/-) knockout mice. Faseb J. 14:1117-1125 (2000). [ Links ]
51. Stenbit, A.E., E.B. Katz, J.C. Chatham, D.L. Geenen, S.M. Factor, R.G. Weiss, T.-S. Tsao, A. Malhotra, V.P. Chacko, C. Ocampo, L.A. Jelicks and M.J. Charron. Preservation of glucose metabolism in hypertrophic GLUT4 null hearts. Am. J. Phys.279:H313-H318 (2000). [ Links ]
52. Qiang Li, Andrei Manolescu, Mabel Ritzel, Sylvia Yao, Melissa Slugoski, James D. Young, Xing-Zhen Chen, and Chris I. Cheeseman. Cloning and functional characterization of the human GLUT7 isoform SLC2A7 from the small intestine, 2004. Am J Physiol Gastrointest Liver Physiol 287: G236–G242. [ Links ]
