











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.26 n.11 Caracas nov. 2001
Alicia S.G. Poi de Neiff. Bióloga y Doctora en Ciencias Biológicas. Profesora Titular de Limnología, Facultad de Ciencias Exactas y Naturales y Agrimensura de la Universidad Nacional del Noreste (UNNE), Argentina. Miembro de la Carrera del Investigador Científico del Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET). Dirección: Casilla de Correo 291 (3400) Corrientes, Argentina. e-mail: neiff@arnet.com.ar
Sylvina Lorena Casco. Licenciada en Zoología. Profesora en Biología. Tesista doctoral, UNNE. Becaria del CONICET en el Centro de Ecología Aplicada del Litoral (cecoal). Adscripta a la cátedra de Botánica II (Sistemática) en la Facultad de Ciencias Exactas y Naturales y Agrimensura (unne). e-mail: sylvina@compunort.com.ar
Resumen
La caída de hojas de Copernicia alba (palma blanca) fue medida durante un año en dos sitios de un palmar con diferente densidad de árboles. La descomposición y la colonización por invertebrados fueron estimadas durante 261 días, utilizando la técnica de las bolsas de litera, en dos palmares con diferente régimen de inundación. La caída anual de hojas fue 0,97 y 1,83 t.ha-1 año-1 dependiendo de la densidad de los árboles. El máximo valor promedio diario (2,83 g.m-2día-1) fue registrado durante el verano. El tiempo medio para la descomposición de la broza fue de 103 días en el palmar con mayor duración de la inundación y de 94 días en el que estuvo menos días inundado. Se encontraron diferencias significativas en la abundancia de los invertebrados en diferentes palmares. El mayor número de invertebrados por gramo remanente de broza (13 individuos) fue colectado después de 125 días de incubación cuando dominaban los oligoquetos (Enchytraidae). Después de 8, 35, 66 y 261 días otros Oligochaeta (Dero spp.), Diptera (larva de Chironomus) y Coleoptera fueron los taxa más abundantes. Estos resultados tienen implicancia en el aporte de materia orgánica a la planicie de inundación y en el papel de los invertebrados durante su descomposición.
Summary
Leaf litterfall of Copernicia alba (white palm) was measured for one year in two sites of a subtropical floodplain forest with different tree densities. Decomposition and invertebrates colonization were estimated by the litter bag technique during 261 days in two palm forests with different inundation regimes. Depending on the tree densities, annual litterfall were 0.97 and 1.83 t.ha-1year-1. The maximum average daily value (2.83 g.m-2day-1) was registered during the summer. The "half time" of the litter was 103 days in the palm forest with the longest duration of the flooding and 94 days in that which had less number of flooding days. Significant differences were found for invertebrate abundance in different palm forests. The highest number of invertebrates per g remaining litter (13 individuals) was collected after 125 days of incubation when Oligocaheta (Enchytraeidae) were dominant. After 8, 35, 66 and 261 days other Oligochaeta (Dero spp.), Diptera (Chironomus larvae) and Coleoptera, were the most abundant taxa. These results have implications in the input of organic matter to the floodplain and the role of invertebrates during decomposition.
Resumo
A caída de folhas de Copernicia alba (palma branca) foi medida durante um ano em dois lugares de um palmar com diferente densidade de árvores. A descomposição e a colonização por invertebrados foram estimadas durante 261 dias, utilizando a técnica das bolsas de liteira, em dois palmares com diferente regime de inundação. A caída anual de folhas foi 0,97 e 1,83 t.ha-1ano-1 dependendo da densidade das árvores. O máximo valor médio diário (2,83 g.m-2dia-1) foi registrado durante o verão. O tempo médio para a descomposição das material orgânico foi de 103 dias no palmar com maior duração da inundação e de 94 dias no que esteve menos dias inundado. Foram encontradas diferencias significativas na abundância dos invertebrados em diferentes palmares. O maior número de invertebrados por gramo remanescente de materiais orgânicos (13 indivíduos) foi coletado depois de 125 dias de incubação quando dominavam os oligoquetos (Enchytraidae). Depois de 8, 35, 66 e 261 dias outros Oligochaeta (Dero spp.), Díptera (larva de Chironomus) e Coleóptera foram as taxa mais abundantes. Estes resultados implicam no aporte de matéria orgânica à planície de inundação e no papel dos invertebrados durante sua descomposição.
PALABRAS CLAVE / Bosques Tropicales / Palmares Inundables / Copernicia alba / Río Paraná /
Recibido: 03/08/2001. Modificado: 04/10/2001. Aceptado: 16/10/2001
Introducción
Los palmares de Copernicia alba Morong. caracterizan el paisaje del nordeste argentino en áreas periódicamente inundadas de la planicie distal de los ríos Paraná y Paraguay (Figura 1), desde el Gran Pantanal hasta el norte de Santa Fe (paralelo de 28º S, aproximadamente). En la provincia del Chaco los palmares están constituidos por árboles de 8 a 13m de alto y de 10 a 40cm de diámetro. Las láminas palmadas de hasta 70cm de largo están sujetas al extremo del tronco por pecíolos (1,5m de largo) armados de gruesas espinas.

Figura 1. Distribución de Copernicia alba (marcado en gris) y ubicación del área de estudio.
Aun cuando los palmares ocupan extensas áreas en las zonas de transición entre la selva amazónica y las sabanas tropicales, existen escasos trabajos (Frangi y Lugo, 1985; Lugo y Frangi, 1993; Odum, 1970) referidos al aporte de materia orgánica y su velocidad de descomposición.
En la planicie de inundación del río Paraná, en el área de la confluencia con el río Paraguay, se han realizado investigaciones sobre la descomposición y colonización de la materia orgánica producida por distintas especies de plantas acuáticas (Poi de Neiff y Neiff, 1988; Neiff y Poi de Neiff, 1990; Bruquetas de Zozaya y Neiff, 1991; Poi de Neiff, 1991; Bruquetas de Zozaya y Poi de Neiff, 1993), no existiendo información disponible sobre Copernicia alba.
Esta contribución tiene como objetivo conocer el aporte de materia orgánica de C. alba a la planicie del río Paraná y su coeficiente de decaimiento en palmares con distinto régimen de inundación. Simultáneamente, cuantificar el número de invertebrados que habitan la materia orgánica en descomposición.
Materiales y Métodos
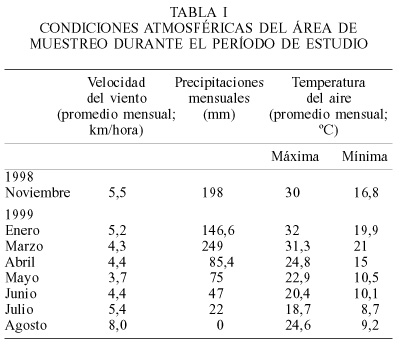
El área de estudio (Figura 1) tiene veranos calurosos e inviernos suaves. En el período comprendido entre noviembre de 1998 y septiembre de 1999 la temperatura máxima (media mensual) más elevada se registró en enero (32°C) y la mínima fue de 9,2°C en el mes de julio (Tabla I). Según el Boletín Agrometeorológico del I.N.T.A. en Colonia Benítez (ubicado en las cercanías del área estudiada) la velocidad del viento a 2m del suelo varió entre 3,7km.h-1 (mayo de 1999) y 8,0km.h-1 en agosto de 1999 (Tabla I). La misma fuente registró un período lluvioso entre noviembre 98 y marzo 99 con máximo aporte en este último mes (249mm mensuales) y un período de escasez de lluvias entre mayo y agosto con mínimos inferiores a 22mm mensuales (Tabla I). Desde septiembre 97 a octubre 98, el área estuvo influenciada por el fenómeno de "El Niño". El río Paraná tuvo un régimen irregular con crecidas extremas que superaron los 7m en el hidrómetro del Pto. de Corrientes y la planicie permaneció inundada por varios meses.
El aporte de materia orgánica de las hojas de Copernicia alba para el período comprendido entre noviembre 98 y septiembre 99 fue estimada en un palmar que permaneció seco durante la mayor parte de este período (PS, 27°25’32"S, 58°52’42"W). En éste se escogieron dos sitios (PS1 y PS2) con diferente distribución de las palmas. Para colectar las hojas caídas se delimitaron, en cada sitio, tres áreas de 80m2 cada una, medidas en un radio de 5m desde la palma ubicada como punto de referencia. Esta área de muestreo involucraba, en promedio, 2 palmas en PS1 y 11 en PS2. A partir del peso la totalidad de las hojas caídas secadas al aire se calculó su peso seco constante utilizando alícuotas de 15g secadas en estufa (48 hs a 105°C) en los dos sitios. El promedio quincenal se expresa en g.m-2 dia-1y el aporte anual en t.ha-1 año-1.
Para conocer la velocidad de descomposición de la materia orgánica producida por Copernicia alba se realizaron experiencias en el palmar donde se midió la caída de hojas (PS) y en otro palmar cercano (PI, 27°26’06"S, 58°54’10"W). PS sólo recibió agua de inundación en octubre 98 durante un lapso de 30 días. PI estuvo inundado desde el inicio del experimento hasta unos 125 días, observándose suelo seco al final del experimento (junio 99). La condición de inmersión prolongada de la materia orgánica en el suelo de los palmares sólo se produce naturalmente durante inundaciones extraordinarias.
En cada palmar se colocaron 20 bolsas de nylon de 20 x 30cm y 2mm de apertura de malla conteniendo 20g de hojas secadas al aire. El experimento fue diseñado para extraer entre 3 y 5 bolsas a los 8, 35, 66, 125 y 261 días de su inicio en septiembre 98.
El material extraído fue lavado para separar, mediante filtración por tamices de 500, 250 y 125µm, los invertebrados asociados al detrito. Posteriormente, el material vegetal remanente fue secado en estufa a 60°C y los invertebrados contados con microscopio estereoscópico.
Al inicio del experimento se pesaron 5 réplicas de 20g de hojas secados al aire y a 60°C para convertir el peso inicial en peso seco constante.
El coeficiente de decaimiento (k) fue calculado a partir de la ecuación Wt=W0.e-kt (Olson, 1963) estimándose a partir de este valor el tiempo para que se descomponga el 50% de la materia vegetal ("vida media"). Se cuantificó la densidad de invertebrados por bolsa y el número de individuos por gramo remanente de materia seca.
Los valores de peso seco de las hojas caídas en cada palmar fueron comparados estadísticamente con ANOVA, previa transformación logarítmica (Steel y Torrie, 1985). Para comprobar la significación de los resultados obtenidos en la experiencia de colonización por invertebrados se utilzó el test Mann-Whitney U (Zar, 1984).
Las experiencias de descomposición fueron realizadas con temperaturas del agua que variaron entre 28°C en verano y 5,5°C en invierno. Al inicio de la experiencia, la profundidad del agua (32cm) permitió medir sus condiciones químicas. El oxígeno disuelto fue bajo (1,27 ± 1,14 mg.l-1; n=3), el pH ácido (6,48 ±0,07; n=3) y la concentración de nutrientes escasa (nitratos+nitritos= 15µg.l-1, amonio= 10 µg.l-1).
Resultados
La producción mensual de broza medida en el palmar (PS) se representa en la Figura 2. Las diferencias fueron significativas al comparar los sitios con mayor y menor densidad de palmas (ANOVA, F1-38 = 6,76; P< 0,05). La caída anual de hojas para el período estudiado fue 0,97 t.ha1año-1 (PS1) y 1,83 t.ha-1año-1 (PS2). En ambos sitios la caída diaria de hojas presentó oscilaciones estacionales las que resultaron más marcadas en el palmar cerrado cuyo valor máximo (2,83 g.m-2 dia-1) se registró en diciembre 98. En PS1 la máxima caída diaria fue menor con máximos en enero 99 (1,02 g.m-2 día-1).

Figura 2. Media de la producción mensual de broza (las barras representan DS).
El porcentaje de pérdida de peso de las hojas de Copernicia alba fue semejante en ambos palmares (Figura 3) y su coeficiente de decaimiento (k) calculado para 261 días fue 0,0074 (PS) y 0,0067 (PI). El tiempo estimado para que se descomponga el 50% de las hojas fue de 94 y 103 días, respectivamente. Sin embargo, los k parciales calculados después de cada período de incubación muestran que en PI la descomposición fue más rápida durante los primeros 8 días, manteniéndose hasta los 66 días, mientras que en PS, la velocidad de descomposición fue mayor entre los 8 y 35 días.

Figura 3. Descomposición de hojas de Copernicia alba (las barras representan DS).
El número de invertebrados por gramo remanente de hojas secas fue significativamente distinto (U=151, U0,05,14,14 = 141, P <0,01) al comparar palmares con diferente número de días con suelo inundado (Figura 4). Los valores máximos y mínimos variaron entre 0 y 6 en PS y entre 2 y 13 en PI.
La mayor colonización por invertebrados (Figuras 4 y 5) se registró en PI a los 125 días de iniciado el experimento, cuando el suelo del palmar estaba cubierto por una fina película de agua. En esta situación predominaron oligoquetos enquitreidos en tanto que a los 66 días y al comienzo del experimento fueron dominantes oligoquetos naididos (Tabla II) registrándose hasta 149 individuos por bolsa, es decir unos 3700 ind.m-2. Después de 35 días de incubación las bolsas de broza estaban colonizadas casi exclusivamente por larvas y pupas de Chironomus grupo riparius (Tabla II). Al final del experimento en el suelo seco del palmar se colectaron adultos, larvas y pupas de Staphilinidae, Carabidae y Pselaphidae (Coleoptera).

Figura 4. Número de invertebrados por 1g de peso seco remanente durante la descomposición de las hojas (las barras representan DS).

En PS sólo se registró colonización por invertebrados después de 125 y 261 días de incubación (Figura 5) y la mayor abundancia de invertebrados (61 individuos por bolsa) ocurrió al final del experimento.

Figura 5. Número de invertebrados por bolsa de broza y abundancia relativa de los grupos taxonómicos que la colonizan.
Discusión
El aporte de hojas a la planicie es bajo si se lo compara con los resultados obtenidos para otros bosques fluviales en el mismo tramo del río Paraná (Neiff y Poi de Neiff, 1990) y con los del río Amazonas (Franken et al., 1979; Klinge, 1977). En este río se incluyen bosques pluriespecíficos donde las palmas representan el 10% de la caída total y aportan sólo 0,2 t.ha1año-1 (Franken et al., 1979). Los mismos autores señalan que, cuando las hojas muertas permanecen unidas a las palmas durante un tiempo largo, el aporte de hojas es desestimable. En palmares de Prestoea montana (Frangi y Lugo, 1985) la cosecha media de litera fue 545 g.m-2 (incluyendo troncos y otras misceláneas) lo que indica la gran variación que existe entre distintas especies de la foresta subtropical inundable.
Los picos de caída de hojas no son coincidentes con los períodos de máxima velocidad del viento y se registraron durante la estación lluviosa, que coincidió con el período durante el cual la planicie recibió agua de inundación del río Paraná.
La velocidad de descomposición de la materia orgánica sobre el suelo fue rápida si se la compara con otras palmas cuyo tiempo medio puede llegar a 462 días (Frangi y Lugo, 1985) lo que podría relacionarse con diferencias en la composición de los tejidos de las distintas especies de palma. Otra posible explicación estaría dada por la duración y frecuencia del hidroperíodo, que depende de la ubicación topográfica de los palmares en la planicie de inundación. Nuestros resultados coinciden con los obtenidos en otras florestas riverinas (Brinson, 1977; Neiff y Poi de Neiff, 1990; Bruquetas y Neiff, 1991) en las cuales las mayores pérdidas de materia seca ocurren cuando las hojas entran en contacto con el agua. La rápida descomposición en los primeros 8 días de incubación en el palmar inundado puede ser atribuida a la lixiviación de las sustancias orgánicas e inorgánicas (Brinson, 1977). En bañados de la planicie del río Paraná próximos al área de estudio, los tiempos medios para la descomposición de Eichhornia crassipes en condiciones de acidez del agua y baja concentración de oxígeno disuelto oscilan entre 37 y 86 días, según se trate de hojas o raíces (Poi de Neiff, 1991). Las hojas de esta macrófita tienen porcentajes de fibra (27 a 29%) y de lignina (0,91 a 1,0%) muy bajos (Neiff et al., 2001) si se los compara con los resultados preliminares obtenidos para C. alba (fibra= 77,6%, lignina= 24,3%). Otras macrófitas de hojas duras, tales como Typha latifolia y Thalia multiflora necesitan entre 231 y 126 días respectivamente para que se descomponga el 50% de sus hojas en condiciones del agua semejantes (Bruquetas de Zozaya y Neiff, 1991; Bruquetas de Zozaya y Poi de Neiff, 1993).
Hubo mayor colonización por invertebrados en el palmar inundado que en el palmar seco, sin embargo, la velocidad de descomposición fue semejante en ambos sitios. Este hecho indica una baja influencia de los invertebrados en el fraccionamiento de las hojas de palma. La mayoría de los taxa registrados pertenecen a la categoría trófica de recolectores o predadores y no se registran partidores. En el sitio de muestreo no se observaron invertebrados de mayor tamaño que la apertura de malla usada, fraccionando las hojas. Este hecho puede relacionarse con la dureza de sus tejidos y con la ausencia de partidores en la planicie de grandes ríos como el Paraná (Neiff y Poi de Neiff, 1990; Bruquetas de Zozaya y Poi de Neiff, 1993) y el Amazonas (Furch y Junk, 1997).
Los resultados de este trabajo indican que el aporte de materia orgánica y el procesamiento del detrito de C. alba en la planicie dependen del régimen de pulsos y constituye diferente oferta trófica para otras comunidades en palmares aislados de la hidrodinámica del río Paraná respecto de aquellos con mayor conexión.
Agradecimientos
Los autores agradecen a Juan José Neiff sus valiosos aportes durante la realización de los muestreos, a dos árbitros anónimos sus atinadas sugerencias, y a Roberto Olivares, Director de Suelos del Ministerio de Producción de la Provincia del Chaco, la provisión de los Boletines Agrometeorológicos del Instituto Nacional de Tecnología Agropecuaria de Colonia Benítez (Chaco). Este trabajo fue financiado por el PIP-CONICET N° 4242.
REFERENCIAS
1 Brinson MM (1977) Decomposition and nutrient exchange of litter in an alluvial swamp forest. Ecology 58: 601-609 p. [ Links ]
2 Bruquetas de Zozaya IY, Neiff JJ (1991) Decomposition and colonization by invertebrates of Typha latifolia L. litter in Chaco cattail swamp (Argentina). Aquatic Botany 40: 185-193. [ Links ]
3 Bruquetas de Zozaya IY, Poi de Neiff A (1993) Descomposición de macrófitos en bañados de la planicie inundable del río Paraná. Ambiente subtropical 3: 1-17. [ Links ]
4 Frangi JL, Lugo AE (1985) Ecosystem dynamics of a subtropical floodplain forest. Ecological Monographs 55 (3): 351-369. [ Links ]
5 Franken M, Irmler U, Klinge DH (1979) Litterfall in inundation, riverine and terra firme forests of Central Amazonia. Tropical Ecology 20 (2): 225-235. [ Links ]
6 Furch K, Junk WJ (1997) The chemical composition, food value and decomposition of herbaceous plants, leaves and leaf litter of floodplain forests. En Junk W (Ed.) The Central Amazon Floodplain, ecology of a pulsing system. Springer-Verlag, Berlin. pp. 187-205. [ Links ]
7 Klinge H (1977) Fine litter production and nutrient return to the soil in three natural forest stands of Eastern Amazonia. Geo-Eco-Trop. 1 (2): 159-167. [ Links ]
8 Lugo AE, Frangi JL (1993) Fruit fall in the Luquillo Experimental Forest, Puerto Rico. Biotropica 25 (1): 73-84. [ Links ]
9 Neiff JJ, Poi de Neiff A (1990). Litterfall, leaf decomposition and litter colonization of Tessaria integrifolia (compositae) in the Paraná river floodplain. Hydrobiologia 203: 45-52. [ Links ]
10 Neiff JJ, Poi de Neiff A, Casco SL (2001) The effect of prolonged floods on Eichhornia crassipes growth in Paraná River floodplain lakes. Acta Limnológica Brasiliensia 13 (1). En Prensa. [ Links ]
11 Odum HT (1970) Summary: an emerging view of the ecological system at El Verde. En Odum HT, Pigeon RF (Eds.) A tropical rain forest. A study of irradiation and ecology at El Verde, Puerto Rico. Division of Technical Information. US Atomic Energy Commission. Oak Ridge, Tennessee. Pp. I-10; 191-281. [ Links ]
12 Poi de Neiff A (1991) Descomposición y colonización del detrito de distintas especies de plantas en ambientes inundables del río Paraná. Biología Acuática 15: 158-159. [ Links ]
13 Olson JS (1963) Energy storage and the balance of producers and decomposers in ecological systems. Ecology 44: 322-331. [ Links ]
14 Steel R, Torrie J (1985) Bioestadística. Principios y procedimientos. 2ª Edición. McGraw-Hill. Bogotá. 622 pp. [ Links ]
15 Zar JH (1984) Biostatistical Analysis (2nd ed). Prentice Hall. New Jersey. 718 pp. [ Links ]
