











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.28 n.9 Caracas set. 2003
VEGETATION OF THE SALADILLO AREA (PROVINCE OF SANTA FE) IN THE SOUTH OF THE CHACO, ARGENTINA
Norma I. Hilgert, José F. Pensiero, Gustavo Marino, Juan P. Lewis and Carlos DAngelo
Norma I. Hilgert. Dr., Universidad Nacional de Córdoba (UNC), Argentina. Professor and Researcher, Centro de Investigaciones Biológicas, Universidad Autónoma de Hidalgo, México. Researcher, CONICET, Argentina. Address: Apartado Postal 69, Plaza Juárez, Pachuca, Hidalgo México C.P. 42001. e-mail: normahilgert@yahoo.com.ar
José F. Pensiero. Dr., UNC. Professor, Facultad de Ciencias Agrarias, Universidad Nacional del Litoral (UNL), Argentina. Researcher, CONICET, Argentina.
Gustavo Marino. Dr. in XX, Universidad Nacional de Buenos Aires, Argentina. Professor, Facultad de Ciencias Agrarias, UNL, Argentina.
Juan P. Lewis. Ph.D., Cambridge University, England. Professor, Facultad de Ciencias Agrarias, Universidad Nacional de Rosario, Argentina. Researcher, CONICET Argentina.
Carlos DAngelo. M.Cs., Pontificia Universidad Católica de Chile. Professor, Facultad de Ciencias Agrarias, UNL, Argentina.
Resumen
En el presente artículo se analiza la vegetación del área delimitada por los ríos Saladillos de la provincia de Santa Fe, Argentina. La región se ubica al sur del Chaco santafesino entre el Arroyo el Toba y los ríos Saladillos al oeste del valle del río Paraná. El clima es cálido a templado, húmedo a subhúmedo. Hay tres tipos de bosques, los "espinillares" de Acacia caven, los "algarrobales" de Prosopis spp. y los "quebrachales" de Schinopsis balansae, distribuidos siguiendo un gradiente geográfico. El número de especies leñosas aumenta hacia el norte, y la estructura y estratificación se hacen más complejas en esa dirección. Las comunidades herbáceas más importantes son los pajonales o sabanas de Spartina argentinensis y Panicum prionitis, junto con otros pastos menos importantes. En las proximidades de las áreas inundadas por largos períodos o durante todo el año se encuentran juncales de Schoenoplectus californicus, totorales de Typha domingensis, canutillares y vegas de distintas especies hidrófilas. Las comunidades se distribuyen siguiendo un gradiente edáfico de salinidad y humedad. El área puede ser considerada un Chaco empobrecido con una fuerte influencia del río Paraná.
Summary
The vegetation of the Saladillo area of the province of Santa Fe, Argentina, is analyzed. The region is located in the south of the Santa Fe Chaco, between the Toba and Saladillo rivers, at the west of the Paraná River valley. Climate is humid to sub-humid and warm to temperate. There are three types of forests, the "espinillares" of Acacia caven, the "algarrobales" of Prosopis spp. and the "quebrachales" of Schinopsis balansae, distributed following a geographical gradient. The number of woody species increases northwards, and stratification and structure becomes more complex in that direction. The most important herbaceous plant communities are tall grasslands or savannas of Spartina argentinensis and Panicum prioniti, along with other less important grasslands. In lower areas flooded for long periods or the whole year, there are rushes of Schoenoplectus californicus, cattails of Typha domingensis and several hygrophilous meadows. The communities are distributed following a soil moisture and salinity gradient. The area can be considered as an impoverished Chaco region with a strong influence of the Paraná River.
Resumo
No presente artigo se analisa a vegetação da área delimitada pelos rios Saladillos da província de Santa Fé, Argentina. A região se localiza no sul del Chaco santafesino entre o Arroio el Toba e os rios Saladillos o oeste do vale do rio Paraná. O clima é quente a temperado, úmido a sub-úmido. Existem três tipos de bosques, os "espinillares" de Acacia caven, os "algarrobales" de Prosopis spp. e os "quebrachales" de Schinopsis balansae, distribuídos seguindo um gradiente geográfico. O número de espécies lenhosas aumenta para o norte, e a estrutura e estratificação se fazem mais complexas nessa direção. As comunidades herbáceas mais importantes são os pajonales ou sabanas de Spartina argentinensis e Panicum prionitis, junto com outros pastos menos importantes. Nas proximidades das áreas inundadas por largos períodos ou durante todo o ano se encontram juncais de Schoenoplectus californicus, totorais de Typha domingensis, canutillares e vegas de distintas espécies hidrófilas. As comunidades se distribuem seguindo um gradiente edáfico de salinidade e umidade. A área pode ser considerada um Chaco empobrecido com uma forte influencia do rio Paraná.
KEYWORDS / Argentina / Chaco / Forests / Grasslands / Savannas /
Received: 04/23/2003. Modified: 08/25/2003. Accepted: 08/27/2003
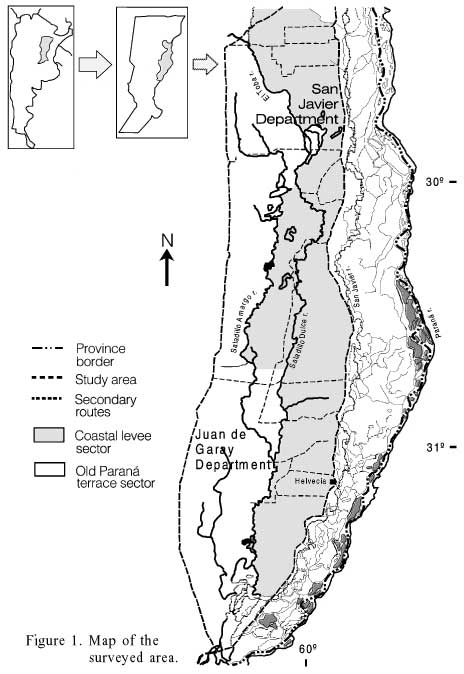
The area of the Saladillos lies at the eastern part of the of Santa Fe province center, comprising the western halves of the Garay and San Javier departments and small parts of other administrative units. At its western limits are the Toba and Saladillo Amargo rivers, while at the eastern limit the Paraná River valley is located (Figure 1). The region consists of plains with a considerable but gentle slope from NW to SE, and without any important orographic or topographic accidents. Most of it is on what the Paraná River bed was, which was modified by an upward movement of the bedrock during the Quaternary (Grassino, 1986; Iriondo, 1991). Several rivers that flow from N to S to the Paraná River drain the area; the main ones are the Saladillo Amargo and the Saladillo Dulce, whose names mean "Bitter Brackish" and "Sweet Brackish", as an allusion to their water taste and composition.
The climate (Table I) is humid to sub humid and warm to temperate; rainfall is mainly during summer and there is a winter drought of variable length (Cáceres, 1980).

The area can be divided into two main physiographic sectors, the old Paraná River terrace and the coastal levee. In the first one, predominant soils are natracualfs, silty loam with sodium carbonate and high pH. On the coastal levee predominant soils are cuarzipsamment, sandy or sandy loams with low nutrient levels (Orellana and Priano, 1978).
The area is covered by woodlands, savannas, tall grasslands and some prairies and hydrophilous meadows. Ragonese (1941) places the area in the region of "espinillares" (prickly mimosa forests), Parodi (1945) considered the area as Chaquenian, and Cabrera (1953; 1976) included the northern portion in the Chaco and the southern portion in the Thorn forest and scrub.
The region is relatively well preserved, although there have been lumbering, grazing and agricultural activities, as well as man-made fires. The extreme south is used for vegetable crops, tomatoes and carrots. Near the town of Helvecia there are important rice cropping areas.
In this paper the vegetation of the Saladillo Rivers area is analyzed, using numerical methods. Plant communities are determined and described.
Material and Methods
Physiognomic units of landscape were determined with 1:150000, 1:50000 and 1:20000 aerial photographs and mapped on 1:100000 charts from the Instituto Geográfico Militar. Then, a general survey of the area was performed in order to define communities of dominance type (Whittaker, 1980), dividing the vegetation into forests, savannas and grasslands.
During November and December 1994, 123 plots (4x4m) of herbaceous communities were analyzed. This plot size was chosen according to Lewis et al. (1990) as the minimum sampling area for this kind of communities. In each plot general descriptions of the vegetation and soil were recorded. All species and their frequencies were recorded using the Braun-Blanquet (1979) cover-abundance combined scale and then converted into the Westhoff and Van der Maarel (1980) scale.
To avoid noise, 112 species, which were found in less than 3 plots, were eliminated from the analysis and an information matrix of 123 plots and 165 species was constructed. Data were ordered using a Detrend Correspondence Analysis (DECORANA), and classified with Wards Cluster analysis using relative euclidean distance as a similarity measure (McCune, 1993).
According to their physiognomic aspects and dominant species, 8 forest stands were selected during July and August 1995. These stands were located in every low level disturbance area found, without any recent anthropic activity, while areas that showed high fragmentation or disturbance intensity were not considered as valid points for this study. The point-centered quarter method (Cottam and Curtis, 1956) was used to measure trees and shrubs. At each stand, mean height and tree cover were assessed, together with total and layer floristic richness and relative frequency of each woody species. Vegetation profiles were made from 250m2 cartographic quadrants (Davis and Richards, 1933; 1934). Data were analyzed using DECORANA and Principal Component Analysis (PCA), and classified according to Wards method (McCune, 1993). In December 1995, the herbaceous layer was analyzed in a similar way as the herbaceous communities.
Plant specimens were determined and deposited in the herbarium of the Facultad de Agronomía y Veterinaria, Universidad Nacional del Litoral (Santa Fe). The nomenclature follows Zuloaga and Morrone (1994; 1996; 1999).
Results
The PCA scatter diagram of forest plots on the plane of axes I and II show three clusters arranged along a horseshoe gradient (Figure 2). DECORANA analysis shows the same three clusters distributed according to a gradient along Axis I (Figure 3). The same three clusters can be recognized when the plots are classified with Wards method (Figure 4).


In most forests Geoffroea decorticans, Prosopis sp., Achatocarpus praecox, Celtis iguanaea and C. pallida are present and can be abundant. The first cluster (A) are "espinillares" where Acacia caven predominates; the second (B) are "algarrobales" of Prosopis sp., which are floristically richer than the former cluster; and the third (C) are "quebrachales" characterized by the presence and abundance of Schinopsis balansae, Aspidosperma quebracho-blanco and Acacia praecox (Table II).

The floristic richness of the woody strata increases significantly northwards (Figure 5) and forests also become structurally more complex (Figures 6 and 7). Individuals of the tree layers are clumped on the three forest types (Figure 6), so the canopy is not continuous but shows gaps of different sizes.


The "espinillares" are more frequent in the south of the region. They are relatively poor and have a 7m high tree layer that sometimes is very disperse. The shrub layer is not conspicuous and Celtis pallida is the only relevant species. There are some epiphytes like Microgramma lycopodioides, Rhipsalis lumbricoides and Tillandsia sp. The herbaceous layer in this region is considerably dense, covering close to 100% of the ground, and Stenotaphrum secundatum, Sida rhombifolia and Cynodon dactylon are frequent (Table III).

TABLE III (CONTINUES)
ASSOCIATION TABLE OF UNDERSTORY

The "algarrobales" are similar, but floristically richer, than the "espinillares". The tree layers are 10-15m high and there is a shrub layer of Celtis pallida, Grabowsquia duplicata, Vassobia brevifolia that sometimes becomes dense (Table II). There are some epiphytes, as in "espinillares", and some lianas such as Pithecoctenium cynanchoides, Dolichandra cynanchoides and Fosteronia glabrescens. The herbaceous layer is heterogeneous, with an important light penetration at in places, being abundant Cynodon dactylon, Carex bonariensis, Paspalum notatum, Setaria stolonifera, S. rhombifolia, etc. In the shaded microsites Oplismenus hirtellus subsp. setarius and Doryopteris pedata are frequent (Table III).
The "quebrachales" are only present in the far north portion of the region. They are much richer than the "algarrobales", with a tree layer >15m high constituted by Schinopsis balansae and Aspidosperma quebracho-blanco, a lower tree stratum and a sometimes dense shrub layer (Figure 7). There are also variants of the "quebrachal": one variant where Aspidosperma quebracho-blanco and Prosopis sp. are abundant on saline soils with a very heterogeneous herbaceous layer formed by a mosaic of "peladales", with a low plant cover composed by Sporobolus pyramidatus, Eragrostis lugens, Selaginella sellowii etc, a depression cover with Luziola peruviana, Leersia hexandra, Panicum hians, and patches of spiny bromeliads Aechmea distichanta and Dyckia remotiflora var. montevidensis (Tables II and III; Figure 4 C1); another variant with Eugenia uniflora, Gleditsia amorphoides, Scutia buxifolia, Hexachlamys edulis (Table II; Figure 4 C2).
When plots of herbaceous communities are classified according to Wards method they appear grouped in two main clusters, one further divided into four plant communities and the other with five very similar hygrophilous communities, a humid prairie, a tall grassland and a halophyte community (Figure 8).

Most of these communities have an upper layer often >1m high of the dominant species and a lower stratum with most other species. The general cover is usually very high, ranging from 70 to 90% of the ground, although plots of the halophyte community may have a high proportion of bare soil (Table IV). Floristically they are very poor and in some plots there is an almost pure population of the dominant species. They contain very few constant species and usually very few species or even none at all in the highest constancy class, while most species are at the lowest constancy class (Table V).


Some of these communities are floristically poorly defined (Table VI) and often the most abundant species are of low constancy.

TABLE VI (continues)
ASSOCIATION TABLE OF HERBACEOUS PLANT COMUNITIES

The four communities of the first cluster are three substitution communities and Spartina argentinensis tall grasslands. In these communities Cynodon dactylon, Setaria parviflora, Aster squamatus, Cyperus entrerianus, Eryngium echinatum and Chaetotropis elongata are fairly constant. Of the substitution communities, 1 and 2 are the richest floristic communities of the region with 133 and 115 species recorded in each of them and the highest average number of species per plot, while community 3 is rather poor, with Cynodon dactylon as the dominant species. Sporobolus indicus, Conyza bonariensis, Dichondra microcalyx and Sisyrinchium platense are common in the three communities. The first two substitution communities are very close to each other (Figure 8), Paspalum notatum, Eragrostis lugens, Deyeuxia viridiflavescens and Verbena gracilescens are fairly constant and faithful to the first one, and the second community appears to be a humid variant of the first one with Eryngium paniculatum, Cyperus corymbosus, Paspalum rufum, Paspalum intermedium, etc. The first is a rather low and comparatively rich prairie or turf, while the second community is a tall grassland.
Community 3 is a turf of Cynodon dactylon with few other species, which may be the result of plowing and overgrazing the Spartina argentinensis tall grasslands and savannas.
Community 4 is a tall grassland or savanna of Spartina argentinensis. The dominant species cover is variable, and at high places the floristic richness is low, and vice versa. Other species of the community are Sarcocornia perennis, Sesuvium portulacastrum, Eryngium coronatum and Cynodon dactylon.
In the other main cluster are gathered hygrophilous communities together with Panicum prionitis tall grasslands or savannas, and halophytes communities. Five of the hygrophilous communities are very close to each other (5 to 9); they are the rushes of Schoenoplectus californicus (5) which appear in rather deep depressions, flooded almost the whole year, with an upper layer of variable density of Schoenoplectus californicus and sometimes a lower layer very similar to the other hygrophilous communities or a Pistia stratiotes pleuston. Community 9 is similar to this community, but the upper layer is a very dense population of Typha domingensis and hardly any other species. Communities 6, 7 and 8 are very similar to each other with the difference between 6 and 7 being the presence of a tall layer of Thalia multiflora and Sagittaria montevidensis, community 7 is a meadow of Ludwigia peploide and community 8 is a hygrophilus turf where Echinochloa helodes, Leersia hexandra and Eleocharis parodii are very frequent. Community 10 is also a low turf of hydrophilous gramineae dominated by Luziola peruviana. The most different communities of this group are successively chained to the hygrophilous communities. Panicum prionitis tall grasslands in the first place and then the community of halophytes (12). The first one has an upper layer of the dominant species and a low one composed mainly by more hygrophilous species. The most common species of the halophytes communities are fleshy Chenopodiaceae (Sarcocornia perennis) and Sesuvium portulacastrum, and in some places it has an upper layer of Spartina densiflora that can be very dense.
When data are analyzed with DECORANA, the hygrophilous communities are pulled to one side by the first axis and all others to the right, while the second axis pulls the halophyte community to the upper part of the dispersion diagram and most other plots to the bottom (Figure 9).

Discussion
There are three kinds of forests that are not very different from each other in the region. They are ordered along gradients. The most important gradient is a latitudinal one, as floristic richness, stratification and complexity increase northwards. But there may be an elevation gradient as well. Further north from this region, floristic richness of tree layers increases even more (Lewis, 1991; Lewis et al., 1994). The forests of Schinopsis balansae appear in the north of the region and are strictly chaquenian forests (Ragonese, 1941; Cabrera, 1976; Lewis and Pire, 1981). The "algarrobales" and "espinillares" are more widespread towards the south, but as in the Chaco, on low saline soils there are "algarrobales" of Prosopis sp. (Lewis and Pire, 1981; Lewis, 1991).
The most widespread herbaceous communities of the region are tall grasslands. Hygrophilous communities, on the other side, cover relatively little space and are confined to small internal depressions.
The two most important tall grasslands are the Spartina argentinensis and Panicum prionitis. The former is a halophilous community very widespread further north which covers a considerably large area in the submeridional lowlands of the Chaco (Lewis et al., 1990) and in most depressed saline areas of the eastern Chaco (Morello and Adámoli, 1974; Lewis and Pire, 1981). Also there are Spartina argentinensis tall grasslands surrounding lake Mar Chiquita in Cordoba (Sayago, 1969), and there are several smaller stands further south, down to parallel 33oS (Lewis and Collantes, 1973; DAngelo et al., 1987). The latter is the most important grassland of the Paraná River valley (Franceschi and Lewis, 1979) but is very common in depressed areas of the eastern Chaco (Lewis and Pire, 1981) and the Paranense phytogeographical province (Cabrera and Willink, 1980). There are also other tall grasslands, such as the Paspalum rufum ones, but they are the result of the destruction of one of these communities. These communities are hyper-seasonal savannas (Sarmiento, 1984), and are flooded during some periods followed by rather long droughts, with some scattered trees. Often, tall grasslands are burnt in order to improve their forage quality, as the re-sprouts of the dominant species are more palatable than the old sclerophyllous leaves. However, periodical fires reduce the floristic richness of these communities and promote the strong dominance of the tall bunch grasses Panicum prionitis and Spartina argentinensis.
Among the hygrophilous communities, both the Schoenoplectus californicus rushes and the Typha domingensis ones are very widespread on the American continent. The former species is found in the Pampa region (Vervoorst, 1967; Lewis et al., 1985), in the Chaco (Morello and Adámoli, 1974; Lewis and Pire, 1981; Franceschi and Lewis, 2000), on the valley of the Paraná River (Burkart, 1957; Franceschi and Lewis, 1979), and even on the high Andes (Heiser, 1979). The latter species is also found in the Pampa region (Vervoorst, 1967), in the Chaco region (Franceschi and Lewis, 2000), the Brasilian Pantanal (Prance and Schaller, 1982), and similar communities with Typha angustifolia and Typha latifolia exist up to North America (Grace and Wetzel, 1981). The other hygrophilous communities are very similar to each other and are very common on the whole Chaco region, and similar as well to some of the communities in the Paraná River valley.
On very saline soils of the Chaco-pampean plain there are communities characterized by the presence of succulent plants, fundamentally Sarcocornia perennis. We have not enough data, but some of the stands have an upper layer of Spartina densiflora and in other regions tall grasslands of this species have been described (Lewis et al., 1985; 1990; DAngelo et al., 1987). Communities with fleshy chenopods, different Sarcocornia species, together with Spartina, are common in marshes located at both sides of the Atlantic Ocean (Rieley and Page, 1990).
If Tables III and VI are compared, the forest understory has a high proportion (31%) of Winter graminea that are scarce in the grassland and savanna regions, where most grasses are Summer graminea.
Lumbering, fire, grazing and agriculture constitute the main disturbance sources in the region. Fires shape the tall grasslands, but plowing promotes their substitution by other grassland types. Forests are completely destroyed to give way to agriculture or forage crops, which are later abandoned and replaced by one of the substitution communities. On the best drier and less saline soils appear the Paspalum notatum prairie, which is rather similar to the "flechillar de abra" found in a neighboring area (Hilgert and DAngelo, 1996) and the Pampean "flechillares" (Vervoorst, 1967; León and Marangon, 1980; León and Anderson, 1983; Lewis et al., 1985) but with very little or none Stipa spp. Individuals. On other soils, other substitution tall grasslands appear which are not well-defined communities, and when overgrazed or present on old rice fields, the vegetation is replaced by a continuous Cynodon dactylon turf.
The hygrophilous communities are strongly chained together as occurs in other regions (Carnevale et al., 1999). This fact and the scatter diagram of data analyzed with DECORANA strongly suggest that all communities are distributed along two main gradients, possibly a soil moisture one and a salinity one, although this has to be proved in other studies.
The region is an impoverished Chaco, lying also in the far south, is less tropical and winter is colder. Also, it appears to have a strong influence of the Paraná River, so the vegetation has strong affinities with the Paranense phytogeographical province.
ACKNOWLEDGEMENTS
The authors thank Ruben Favot, Marcial Bugnon, Jorge Carbone and Pablo Carletti for their collaboration in fieldwork, Susana Stofella for her orientation and training with the Multivariate Analysis, Guillermo Gil for Figure drawings, and Alejandro Córdoba for improving the English text. Finantial aid was provided by CAID, Universidad Nacional del Litoral and CONICET.
REFERENCES
1. Braun-Blanquet J (1979) Fitosociología. Blume. Madrid, Spain. 820 pp. [ Links ]
2. Burkart A (1957) Ojeada sinóptica sobre la vegetación del delta del río Paraná. Darwiniana 11: 457-561. [ Links ]
3. Cabrera AL (1953) Esquema fitogeográfico de la República Argentina. Rev. Mus. La Plata (N.S.) 8, Sec. Bot. 33: 87-168. [ Links ]
4. Cabrera AL (1976) Regiones fitogeográficas argentinas. ACME. Buenos Aires, Argentina. 85 pp. [ Links ]
5. Cabrera AL, Willink A (1980) Biogeografía de América Latina. OEA. Serie Biológica Nº13. Washington DC, USA. 122 pp. [ Links ]
6. Cáceres LM (1980) Caracterización climática de la provincia de Santa Fe. Ministerio de Agricultura y Ganadería. Santa Fe. Departamento Aguas. Santa Fe, Argentina. 35 pp. [ Links ]
7. Carnevale NJ, Torres PS, Lewis JP (1999) Análisis numérico y estacionalidad de las comunidades higrófilas de los bajos submeridionales santafesinos (Argentina). Bol. Soc. Arg. Bot. 34: 39-46. [ Links ]
8. Cottam G, Curtis JT (1956) The use of distance measures in phytosociological sampling. Ecology 37: 451-460. [ Links ]
9. DAngelo CD, Prado DE, Stofella SL, Lewis JP (1987) The Subchaquenian vegetation of the province of Santa Fe (Argentina). Phytocoenologia. 15: 329-352. [ Links ]
10. Davis TAW, Richards PW (1933) The vegetation of Moraballi, British Guiana: an ecological study of a limited area of tropical rainforest. Part I. J. Ecol. 1: 350-384. [ Links ]
11. Davis TAW, Richards PW (1934) The vegetation of Moraballi, British Guiana: an ecological study of a limited area of tropical rainforest. Part II. J. Ecol. 22: 106-155. [ Links ]
12. Franceschi EA, Lewis JP (1979) Notas sobre la vegetación del valle santafesino del río Paraná. R.A. Ecosur 6: 55-82. [ Links ]
13. Franceschi EA, Lewis JP (2000) Composición florística y estacionalidad de Juncales de Scirpus californicus (Cyperaceae) y Totorales de Typha domingensis (Typhaceae) en los bajos submeridionales (Argentina). Bol. Soc. Arg. Bot. 34: 171-178. [ Links ]
14. Grassino SB (1986) Análisis integral de la provincia de Santa Fe. Imprenta Oficial de la Provincia de Santa Fe. Argentina. 383 pp. [ Links ]
15. Grace JB, Wetzel RG (1981) Habitat partitioning and competitive displacement in cattails (Typha): Experimental field studies. American Naturalist 118: 463-474. [ Links ]
16. Heiser CB (1979) The Totora (Scirpus californicus) in Ecuador and Perú. Econ. Bot. 32: 222-236. [ Links ]
17. Hilgert NI, DAngelo CH (1996) Las Comunidades Vegetales de los Dptos. Castellanos y Las Colonias (Santa Fe). Bol. Soc. Arg. Bot. 32: 3-16. [ Links ]
18. Iriondo MH (1991) El Holoceno en el Litoral. Mus. Prov. Ciencias Naturales Florentino Ameghino 3: 1-40. [ Links ]
19. León RJC, Anderson DL (1983) El límite occidental del pastizal pampeano. Tuexenia 3: 67-83. [ Links ]
20. León RJC, Marangon N (1980) Delimitación de comunidades en el pastizal puntano. Sus relaciones con el pastoreo. Bol. Soc. Arg. Bot. 19: 277-288. [ Links ]
21. Lewis JP (1991) Three levels of floristical variation in the forest of Chaco, Argentina. J. Veget. Sci. 2: 125-130. [ Links ]
22. Lewis JP, Collantes MB (1973) El Espinal Periestépico. Ciencia e Investigación 29: 345-408. [ Links ]
23. Lewis JP, Pire EF (1981) Reseña sobre la vegetación del Chaco santafesino. La vegetación de la República Argentina. INTA, Serie Fitogeográfica 18: 1-48. [ Links ]
24. Lewis JP, Collantes MB, Pire EF, Carnevalle NJ, Bocanelli SJ, Stofella SL, Prado DE (1985) Floristic groups and Plant communities of southeastern Santa Fe, Argentina. Vegetatio 60: 67-90. [ Links ]
25. Lewis JP, Pire EF, Prado DE, Stofella SL, Franceschi EA, Carnevale NJ (1990) Plant communities and phytogeographical position of a large depression in the Great Chaco, Argentina. Vegetatio 86: 25-38. [ Links ]
26. Lewis JP, Pire EF, Vesprini JL (1994) The mixed dense forest of the Southern Chaco. Contribution to the study of the flora and vegetation of the Chaco VIII. Candollea 49: 159-168. [ Links ]
27. McCune B (1991) Multivariate analysis on the PC-ORD system. Oregon State University. Corvallis, USA. 139 pp. [ Links ]
28. Morello J, Adámoli J (1974) Las grandes unidades de vegetación y ambiente del Chaco argentino. Vegetación y ambiente de la Provincia del Chaco. INTA. Ser. Fitogeográfica 13: 1-130. [ Links ]
29. Orellana JA, Priano LJJ (1978) Origen y distribución de los suelos santafesinos. Revista FAVE 1: 117-198. [ Links ]
30. Parodi LR (1945) Las regiones fitogeográficas argentinas y sus relaciones con la industria forestal. Plant Sci. Lat. Amer. 16: 127-132. [ Links ]
31. Prance GT, Schaller GB (1982) Preliminary studies of some vegetation types of the Pantanal, Mato Grosso, Brazil. Brittonia 34: 228-251. [ Links ]
32. Ragonese AE (1941) La vegetación de la provincia de Santa Fe. Darwiniana 5: 369-416. [ Links ]
33. Rieley JO, Page S (1990) Ecology of Plant Communities. Logman. Harlow, England. 178 pp. [ Links ]
34. Sarmiento G (1984) The ecology of Neotropical Savannas. Harvard University Press 235 p. [ Links ]
35. Sayago M (1969) Estudio fitogeográfico del norte de Córdoba. Bol. Acad. Nac. Cs. Córdoba 46: 123-427. [ Links ]
36. Vervoorst FB (1967) Las comunidades vegetales de la depresión del salado. Serie. Fitogeográfica. INTA. Buenos Aires, Argentina. 262 pp. [ Links ]
37. Westhoff V, Van Der Maarel E (1980) The Braun-Blanquet approach. In Whittaker RH (Ed.) Classification of Plant Communities. W. Junk. The Hague, Netherlands. pp. 287-339. [ Links ]
38. Whittaker RH (1980) Dominance Types in Classification of Plant Communities. In Whittaker RH (Ed.) Classification of Plant Communities. W. Junk. The Hague, Netherlands. pp. 65-81. [ Links ]
39. Zuloaga FO, Morrone O (ed.) (1994) Catálogo de la familia Poaceae en la República Argentina. Monographs in Systematic Botany. Vol. 47. Missouri Botanical Garden 178 p. [ Links ]
40. Zuloaga FO, Morrone O (eds.) (1996) Catálogo de las plantas vasculares de la República Argentina I. Monographs in Systematic Botany. Vol. 60. Missouri Botanical Garden. USA. 323 pp. [ Links ]
41. Zuloaga FO, Morrone O (ed.) (1999) Catálogo de las plantas vasculares de la República Argentina II. Monographs in Systematic Botany. Vol. 74. Missouri Botanical Garden. USA. 1269 pp. [ Links ]
