










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa Odontológica Venezolana
versión impresa ISSN 0001-6365
Acta odontol. venez v.40 n.1 Caracas ene. 2002
ALGUNAS CONSIDERACIONES SOBRE Candida albicans COMO AGENTE ETIOLÓGICO DE CANDIDIASIS BUCAL.
Germán Pardi. Profesor Asociado. Jefe del Departamento de Ciencias Básicas II, Facultad de Odo tología, U.C.V.
Elba Inés Cardozo. Profesor Agregado. Jefe de la Cátedrade Farmacología y Terapéutica Odontológica, Facultad de Odontología, U.C.V.
RESUMEN:
En el presente artículo, se describen los principales aspectos referentes a Candida albicans, microorganismo mayormente implicado en la Candidiasis Bucal, entre los cuales se destacan: Historia, taxonomía, características morfológicas y fisiológicas (haciendo especial énfasis en la ultraestructura del hongo), características de cultivo, algunas consideraciones ecológicas, así como una serie de factores específicos que afectan en la distribución de Candida en la cavidad bucal.
PALABRAS CLAVE: Candida albicans, agente etiológico, Candidiasis Bucal.
ABSTRACT:
In this article, we make reference about some considerations of Candida albicans, the most important aetiological agent implicated in Oral Candidiosis refering to: History, taxonomy, morphological and physiological characteristics (making special emphasis in ultrastructure), culture characteristics, ecological aspects and specific factors afected Candida distribution within the oral cavity.
KEY WORDS: Candida albicans, aetiological agent, Oral Candidiosis.
Recibido para arbitraje:12/12/2001 Aceptado para publicación: 07/02/00
INTRODUCCIÓN:
En años recientes, ha aumentado el interés con respecto a las infecciones producidas por los hongos del Género Candida en sujetos inmunosuprimidos y médicamente comprometidos. Esto ha traido como resultado que se hayan realizado una gran cantidad de investigaciones, dirigidas hacia la identificación de C. albicans como el principal agente involucrado en Candidiasis, una enfermedad infecciosa la cual puede variar desde lesiones superficiales en piel y mucosas hasta la forma sistémica diseminada1. Es la micosis más común en la cavidad bucal humana1,2,3,4.
Sin embargo, Mata5 señala que la Candidiasis, dependiendo del estado inmunitario del huésped y de la localización de las manifestaciones clínicas, puede ser superficial o profunda. Por lo tanto, la considera como No Clasificable.
1.- HISTORIA.
Uno de los aspectos más importantes a considerar es el relativo a los distintos nombres que ha recibido la principal especie patógena: C. albicans. Desde que Robin la denominó en 1853 Oidum albicans, esta especie estuvo incluída en 100 sinónimos y pasada a través de 18 géneros. De estos géneros, sólo 2 han prevalecido por largo tiempo para referirse a esta especie: El Género Monilia, en el cual estaba incluída la especie Monilia candida (Plaut, 1885), luego Monilia albicans (Zopt, 1890) que dominó la literatura hasta el trabajo de C.M. Berkhout, quien en 1923 propone el Género Candida y la especie C. albicans, que fue aceptado por el 3er Congreso Internacional de Microbiología en Nueva York en 1939. Desde entonces se pasaron al Género Candida todas aquellas levaduras que no encajaban en el Género Monilia, por lo cual todas las afecciones producidas por Candida se conocen con el nombre de Candidiasis3,6.
2.- TAXONOMIA.
Los microorganismos involucrados como agentes etiológicos de la Candidiasis, se encuentran actualmente clasificados taxonómicamente de la siguiente forma:
Reino: Hongo
División : Deuteromycota
Clase: Blastomycetes
Familia: Cryptococcaceae
Género: Candida
Especies: albicans (como la más frecuente y virulenta) y otras especies6.
El Género Candida comprende más de 150 especies, cuya principal característica es la ausencia de forma sexual, con excepción de algunas especies micóticas. Son clasificadas como levaduras, las cuales corresponden a hongos con un modo de desarrollo predominantemente unicelular7.
Solamente una docena de las especies pertenecientes al Género Candida poseen la facultad de adaptarse a una temperatura de 37°C. y pueden ser ocasionalmente patógenas para el hombre, estas son entre otras: C. albicans, C. tropicalis, C. kefyr (pseudotropicalis), C. krusei, C. guillermondi, C. parakrusei, C. zeylanvides, C. stellatoidea y C. brumptii3.
Estudios taxonómicos tratan de diferenciar las diversas especies del Género Candida de acuerdo a sus propiedades. Candida paratropicalis no ha sido considerada distinta a Candida tropicalis8. En el caso de Candida claussenii y Candida stellatoidea la similitud de su morfología y fisiología sobrepasan las diferencias que puedan haber entre estas especies9. Diversos estudios sobre C. albicans y C. stellatoidea no consideran estas dos especies como distintas10. Se ha demostrado por métodos electroforéticos que C. albicans y C. stellatoidea difieren en el número y patrón de cromosomas11, pero también se han observado diferencias entre cepas de C. albicans en cuanto a este aspecto se refiere12.
A través de estudios de biología molecular mediante la prueba de Southern Blotting, utilizando enzimas de restricción para fragmentar el ADN, se demostró que C. albicans y C. stellatoidea son idénticas13.
Recientemente, se publicó un reporte donde se hace mención a los nombres viejos, incorrectos u obsoletos que se le daban a algunas especies de Candida y a los nombres aceptados actualmente. Es así como C. claussenii y C. stellatoidea están reclasificadas en la actualidad como C. albicans, por su parte Candida macedoniensis y Candida pseudotropicalis están reclasificadas como Candida kefyr y Candida paratropicalis está reclasificada como C. tropicalis14.
3.- CARACTERISTICAS DEL HONGO.
C. albicans suele presentarse como una célula oval levaduriforme de 2 a 4 micras, con paredes finas; sin embargo, en tejidos infectados también se han identificado formas filamentosas de longitud variable, con extremos redondos de 3 a 5 micras de diámetro y seudohifas, que son células alargadas de levadura que permanecen unidas entre sí3,15,16.
Las levaduras o blastosporas son microorganismos eucarióticos, las cuales se reproducen asexualmente por un proceso específico de división celular conocido como gemación. Este proceso de división implica la producción de nuevo material celular proveniente de la superficie de la blastospora. Cuando el brote o yema ha crecido y se encuentra en su tamaño óptimo, se suscita la división celular y se forma un tabique o septo entre las dos células16.
La forma filamentosa del hongo (hifa), es una estructura microscópica tubular, la cual contiene múltiples unidades celulares divididas por septos y puede surgir a partir de blastosporas o de hifas existentes. Esta crece contínuamente por extensión apical16,17.
La apariencia microscópica de todas las especies de Candida es similar; todas las levaduras son Gram positivas, pero en algunas ocasiones la forma de las blastosporas puede variar de ovoide a elongada o esférica. Microscópicamente, C. albicans presenta dimorfismo, el cual es una transformación de la forma ovoide de las blastosporas (levaduras) gemantes a hifas16,17.
Por su parte, Samson3 sostiene que el material blanco que crece en los medios de cultivo consiste desde el punto de vista microscópico, en pseudomicelio actualmente llamados filamentos de C. albicans. Se presenta bajo condiciones de cultivo semianaeróbico o facultativo y está formado por células elongadas que se mantienen unidas entre sí como una cadena y blastoconidias o blastosporas que están agrupadas en montones a lo largo del pseudomicelio, en los sitios en que los extremos finales de las células pseudomiceliales se empalman con otras. En contraste con otras especies de Candida, C. albicans tiene una marcada tendencia a formar esporas grandes de pared gruesa, denominadas clamidosporas, sobretodo cuando se cultivan en un medio especial como Agar Harina de Maiz; la clamidospora tiene un diámetro de 7 a 8 micras y casi siempre se origina en el extremo del pseudomicelio. Es una importante característica morfológica en la identificación de C. albicans. Asimismo, tiene la capacidad para producir tubos germinales (filamentación en suero) cuando las colonias son inoculadas en 0,5 ml. de suero a temperatura de 37°C. observándose los resultados después de 2 o 3 horas18,19.
Un tubo germinal se define como una extensión filamentosa de una célula levaduriforme que mide alrededor de la mitad del ancho y tres a cuatro veces el largo de la célula18,19. El tubo germinal de C. albicans ha sido descrito como un tubo sin constricción en el punto de origen y tiene una apariencia similar a "espejo de mano". Este puede formarse al inocular células de C. albicans en suero humano (inclusive si el suero ha sido congelado y almacenado), así como en suero de diversos animales como perro, bovino, conejo, cochino de Guinea y caballo. En cambio, este no se forma en suero caliente coagulado18.
La formación de tubos germinales en suero está afectada directamente por la concentración celular en el inóculo, ya que la proporción de células capaces de formar filamentos, disminuye progresivamente al aumentar la concentración celular por encima de 107 células por ml. De igual forma, demostró este autor que el rango de temperatura en el cual se forman los tubos germinales oscila entre 31°C y 41°C18.
La composición química de C. albicans está representada por 20-40% de proteínas y 30-50% de polisacáridos, mientras que la proporción de lípidos es variable20. La fracción lipídica va a depender de la cepa, edad del cultivo, condiciones ambientales y del origen de la fuente de carbono21.
La pared celular de C. albicans está compuesta principalmente por los polisacáridos Manán, Glucán y Quitina. Aunque la síntesis de los componentes de la pared celular está dinámicamente influenciada por las condiciones de crecimiento y por los estadios metabólicos, en la literatura existen bastantes datos acerca de la composición química de dicha pared. El polisacárido manán representa aproximadamente entre 15,2% y 22,9% del peso seco y poco más de 40% de los polisacáridos de la pared celular del hongo. El D-Glucán ß-1-3 y el D-Glucán ß-1-6 constituyen entre 47% y 60% del peso seco de la pared celular22. Otros componentes han sido reportados, tales como proteínas22,23,24,25,26 en cantidades que oscilan entre 6% y 25%, lípidos entre 1% y 7% y Quitina entre 0,6% y 9% del peso de la pared celular22,27. Las proporciones de los componentes que constituyen la pared celular de las levaduras y de los tubos germinales es relativamente similar, aunque la cantidad de Glucán Alcali-soluble y Alcali-insoluble y de Quitina de C. albicans varía de acuerdo con la forma de crecimiento22,28.
Estudios ultraestructurales de la pared celular de C. albicans han demostrado una compleja microarquitectura. La pared tiene un espesor variable y está compuesta por varias capas, las cuales se han puesto de manifiesto por diferencias en la densidad electrónica. El número de capas y su morfología varían; esta variación está relacionada con varios factores tales como: la etapa de crecimiento celular, la forma de crecimiento (como levadura o como tubo germinal), la capa seleccionada para su estudio, el medio de cultivo empleado para el crecimiento celular y los procedimientos de fijación22. La mayoría de los investigadores han descrito cinco capas dentro de la pared celular, las cuales son (de adentro hacia afuera): Manoproteínas, ß-Glucán-Quitina, ß-Glucán, Manoproteínas y una capa de fibrillas22,29,30 (FIGURA 1).
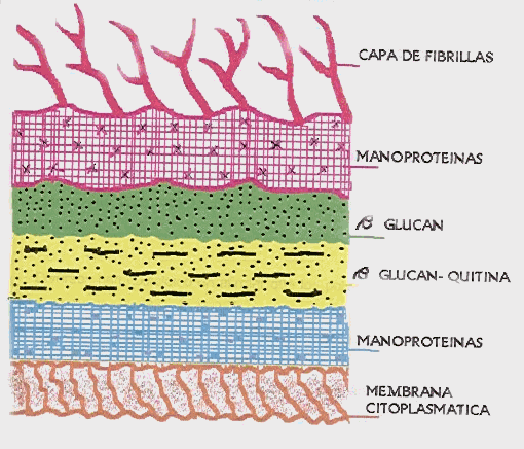
Figura 1: Diagrama esquemático de la pared celular de C. Albicans Tomado de Calderone y Braun 1991
Poulain y colaboradores31 han observado hasta ocho o nueve capas en la pared celular de C. albicans, aunque estos resultados se refieren a una variedad de paredes celulares provenientes de células que crecieron en diferentes medios de cultivo y en distintos períodos de tiempo.
Se ha demostrado que después de cultivar a C. albicans en medios donde haya carencia de nutrientes y someter al hongo bajo esas condiciones por períodos de tiempo muy prolongados, las capas de Manán de la pared celular desaparecen gradualmente32.
Los polisacáridos del tipo Manán están localizados a lo largo de la pared celular y éstos, predominan en las zonas de alta densidad electrónica33,34. Las capas internas de la pared celular están compuestas mayormente por Quitina y Glucán22. Estos componentes le dan rigidez a la célula y son escenciales para la división celular35,36. Están presentes tres tipos de Glucán: 1) Glucán ß-1,6 altamente ramificado, 2) Glucán ß-1,3 altamente ramificado y 3) Un Glucán muy complejo ß-1,6-ß-1,3 mezclado con Quitina. Las proporciones de ciertos tipos de Glucán difieren entre las levaduras y los tubos germinales de C. albicans. Durante las primeras etapas de la formación del tubo germinal, se sintetiza casi exclusivamente Glucán ß-1,322. La Quitina se encuentra en las células en forma de levadura, en las hifas y en los tubos germinales, aunque la proporción es mayor en las hifas28.
La capa externa de fibrillas de la pared celular de C. albicans, tanto en levaduras como en hifas está compuesta de Manán o Manoproteínas, aunque este componente también está localizado en varios lugares de la pared celular. Esta capa ha sido descrita en ocasiones como un revestimiento mucoso o capsular37.
El Manán ha sido identificado como el principal antígeno de la superficie celular de C. albicans22. Basados en estudios de adsorción y aglutinación, Hasenclever y Mitchell38, agruparon a C. albicans en dos serotipos designados A y B.
Básicamente, la representación de las Manoproteínas de la pared celular de C. albicans está constituída por residuos de Manosa unidos entre sí por enlaces a -1,6, los cuales se unen a la porción de Proteína a través de dos residuos de N-Acetil Glucosamina (unidos entre sí por enlaces ß-1,4) y un residuo de Asparagina y residuos de Manosa que se unen a la Proteína a través de residuos de los aminoácidos Serina y Treonina22 (FIGURA 2).

FIGURA 2: Representación de las manoproteínas de la pared celular de C. albicans.
NAcGlc= N-Acetil D Glucosamina
M= Manosa
Ser= Serina
Thr= Treonina
Asn= Asparagina
(Tomado de Calderone y Braun,1991).
La membrana citoplasmática es una estructura que reviste gran importancia, ya que los antibióticos antimicóticos actúan a nivel de la misma, además de contener las enzimas responsables de la síntesis de la pared celular. Esta presenta una doble capa compuesta por lípidos y posee invaginaciones, que se observan como surcos de 200 a 300 nanómetros de longitud, por 35 a 40 nanómetros de espesor39,40. Además de los lípidos, la membrana citoplasmática está compuesta por grandes cantidades de proteínas y carbohidratos en menor proporción41,42.
En el citoplasma, al igual que otras células eucarióticas, C. albicans presenta: ribosomas, mitocondrias con doble capa, gránulos de glucógeno y vacuolas que, contienen en algunas ocasiones cuerpos lipídicos y gránulos de polifosfato. El núcleo es típico de una célula eucariótica, con membrana nuclear limitante, uno o varios nucleolos, ADN y ARN y varios cromosomas43.
El metabolismo de C. albicans se ha relacionado de una forma directa o indirecta con la patogenicidad, la morfología o con los efectos de los antibióticos antimicóticos. El metabolismo de los carbohidratos juega un papel importante en la morfogénesis, en tanto que el metabolismo de aminoácidos y lípidos tiene poca importancia para el crecimiento de este microorganismo44.
Se han podido detectar distintos tipos de fosfolípidos en diversas especies de Candida provenientes de cavidad bucal, tales como C. albicans, C. glabrata, C. guillermondii, C. tropicalis, C. parapsilosis y C. kefyr. Los principales fosfolípidos identificados fueron: Fosfatidiletanolamina y Fosfatidilglicerol. Estos fosfolípidos son muy importantes en relación con el normal funcionamiento de la membrana citoplasmática de los hongos antes mencionados45.
En un estudio reciente, se determinaron cambios fenotípicos en cepas de C. albicans aisladas de la cavidad bucal de pacientes con transplante renal. Se ha sugerido que las cepas del hongo en las que se observaron los cambios fenotípicos, pueden adaptarse a diferentes condiciones ambientales debido a variaciones en sus propiedades bioquímicas, físicas y fisiológicas46.
4.- CULTIVO.
Las especies de Candida crecen bien en medios de cultivo con agar, peptona, dextrosa, maltosa o sacarosa. Las colonias muy pequeñas aparecen en un lapso de 24 a 36 horas en Agar Sabouraud y miden de 1,5 a 2 m.m. de diámetro después de 5 a 7 días. Las colonias son típicamente blancas por completo, pero adquieren un color crema o requemado al continuar envejeciendo. Para aislarlas de las muestras clínicas que siempre llevan bacterias se agregan antimicrobianos como el Cloranfenicol al medio simple47.
En Agar Sabouraud o en otros medios de cultivo similares, las colonias que crecen son lisas, suaves, húmedas y de color y aspecto cremoso. Estas colonias tienen un tamaño que oscila entre 1,5 y 2 m.m. de diámetro, con aspecto de levadura, de consistencia blanda y rápidamente proyectan filamentos hasta la profundidad del agar. Después de 4-5 días se percibe un olor característico de levadura16,47. Otros medios de cultivo en los cuales puede crecer C. albicans son: Pagano-Levin, en el cual las colonias se observan de color crema, Albicans ID (Biomerieux), donde las colonias se observan de color azul y CHROMagar® Candida (CHROMagar), observándose las colonias de esta especie de color verde48.
Las colonias de Candida crecen "in vitro" en condiciones de aerobiosis en medios de cultivo a pH con rango entre 2,5 y 7,5 y temperatura que oscila entre 20°C y 38°C. El crecimiento de colonias se puede detectar entre 48 y 72 horas después de la siembra, y los subcultivos pueden crecer más rápidamente16.
La habilidad de las levaduras de crecer a 37°C es una característica importante a ser considerada en su identificación a partir de muestras clínicas. Las levaduras más virulentas crecen rápidamente a temperaturas que oscilan entre 25°C y 37°C, mientras que las poco virulentas dejan de crecer a 37°C16.
5.- ECOLOGIA.
Los microorganismos del Género Candida son oportunistas que se encuentran como comensales en cavidad bucal, intestino, vagina, secreción bronquial y piel del hombre y de ciertos animales. En la cavidad bucal la colonización es significativamente distinta de sitio a sitio16,47.
En la cavidad bucal de sujetos portadores de especies de Candida, C. albicans comprende entre 60% y 70% de los aislamientos, C. tropicalis comprende 7%, en tanto que C. kruzei y C. guillermondii son aislados con mucha menor frecuencia49.
Una etapa temprana y esencial en el desarrollo de la Candidiasis Bucal es la colonización de la cavidad bucal por parte de C. albicans, un proceso que involucra la adquisición, adherencia y mantenimiento de una población estable de levaduras. La boca posee muchos nichos para la colonización por parte de esta especie, incluyendo entre otros células epiteliales, prótesis dental y células bacterianas de la flora bucal residente50.
No obstante, la capacidad de infección por Candida disminuye porque existe un equilibrio biológico con la flora comensal bacteriana. Las otras especies de Candida se encuentran en la piel, el tubo digestivo y en la naturaleza. C. albicans jamás está presente de manera prolongada en la piel sana, excepto en la región perianal3.
La presencia de C. albicans como comensal en las membranas mucosas de sujetos asintomáticos es común, por lo que en sujetos sanos, existe un balance entre los mecanismos de defensa del hospedero y el potencial invasivo por parte de las levaduras51. Sin embargo, cuando el sistema de defensa del hospedero se daña, tal y como ocurre en sujetos inmunosuprimidos o médicamente comprometidos, la infección por C. albicans, así como por otras especies de Candida puede derivar en el establecimiento de una Candidiasis, la cual se puede manifestar bien sea de manera superficial, involucrando la mucosa bucal, o diseminada, la cual constituye una forma invasiva más seria51,52.
Diversas observaciones clínicas indican que los cambios en el hospedero son usualmente los responsables del desequilibio ecológico. Estos cambios incluyen además de la disminución de los mecanismos de defensa del hospedero, reducción del flujo salival, disminución de las inmunoglobulinas, trauma local con pérdida de la integridad tisular, debilidad general, estados de malnutrición, cuando esta ocurre en sujetos con dietas ricas en carbohidratos, deficiencia de hierro, ácido fólico o vitamina B12, desórdenes endocrinos como hipotiroidismo, enfermedad de Addison (Insuficiencia adrenocortical) y diabetes mellitus, infección por V.I.H., alteraciones de la sangre tales como leucemia aguda y agranulocitosis, antibióticoterapia prolongada, quimioterapia, radioterapia, y xerostomía1,53,54.
C. albicans se puede encontrar en condición facultativamente patógena, desde un estado saprofítico simple, pasando por el comensalismo, hasta la situación de patógeno. Se encuentra libre en la naturaleza donde puede ser aislado, siendo frecuente encontrarlo en la leche bovina. En el ser humano se encuentra como comensal en el tracto respiratorio e intestinal, en la vagina y boca, sobre la piel, donde reside con mayor frecuencia entre los pliegues naturales que son sitios relativamente calientes y de mayor humedad. Resulta muy difícil la infección por C. albicans de animal a hombre, siendo el ciclo de infección más común de animal a animal y de hombre a hombre55. Sin embargo, se realizó un estudio en 36 parejas conyugales que mantuvieron relaciones caracterizadas por intercambio de saliva boca a boca y los resultados obtenidos demostraron que no existió transmisión de C. albicans entre las parejas en las cuales un miembro era portador y el otro no56.
En un estudio realizado por Arendorf y colaboradores57, se determinó la prevalencia de levaduras en individuos surafricanos dentados y con la mucosa bucal sana, y se encontró que 42,5% de los individuos seleccionados portaban levaduras como comensales, mientras que 20% de los mismos, eran portadores de C. albicans como comensal en sus bocas.
Por su parte, Alkumru y Beydemir58 encontraron que la tasa de colonización por Candida fue mayor en el dorso de la lengua que en otros lugares de la cavidad bucal, por lo que sugirieron que la lengua era el reservorio primario de C. albicans en la boca, en tanto que Arendorf y Walker59 afirmaron que la placa dental era colonizada secundariamente por este hongo.
El hallazgo de C. albicans en 58% de los cepillos de dientes de 57 portadores sanos, permitió comprobar "in vitro" que esta especie sobrevive en los cepillos de dientes de nylon y cerda hasta por 2 semanas. También se sugirió en este estudio que el cepillo de cerda, parecía favorecer la supervivencia del hongo60.
6.- FACTORES ESPECIFICOS QUE AFECTAN LA DISTRIBUCION DE Candida EN LA CAVIDAD BUCAL.
- Saliva:
Se ha demostrado que la saliva reduce la capacidad por parte de C. albicans de adherirse sobre el acrílico de las prótesis dentales, mientras que el suero, el cual puede entrar en la cavidad bucal como resultado de un trauma en la mucosa, incrementa la adhesión61,62. En otro estudio se sugirió que las mucinas salivales eran las que actuaban como receptores de las manoproteínas de superficie de C. albicans63. Esto fue confirmado posteriormente al comprobarse que C. albicans adsorbía selectivamente las mucinas salivales, lo cual aumentaba la capacidad por parte de las levaduras de adherírse sobre la superficie de acrílico de las prótesis64,65.
Se ha podido determinar que, la reducción de los niveles de humedad en la cavidad bucal, favorece el crecimiento de bacterias como Staphylococcus aureus, que es resistente a la desecación e inhibe a otros microorganísmos comensales que necesitan de altos niveles de humedad16.
También se ha reportado que, en presencia de pH salival bajo y de altas tensiones de O2 se altera el medio ambiente bucal, trayendo como resultado una reducción en el número de microorganismos de los Géneros Veillonella, Neisseria y Micrococcus, así como un incremento en el número de S. mutans y de especies pertenecientes a los Géneros Candida y Lactobacillus16.
- pH:
Se ha sugerido que el medio ambiente ácido favorece la colonización de la cavidad bucal por parte de especis de Candida16. También se han observado valores bajos de pH en muestras de placa dental obtenidas de prótesis removibles superiores de pacientes con E.S.P., quienes mantenían dietas ricas en glucosa y sacarosa66.
- Adherencia:
Las interacciones entre C. albicans y el hospedero son complejas. En este sentido, se ha sugerido que en los mecanismos de adherencia están involucradas interacciones entre los ligandos de Candida y los receptores de las células hospederas22,67. Por otra parte, se ha señalado que las blastosporas de Candida se adhieren mejor a las células de la mucosa bucal y al acrílico de las prótesis, cuando se hallan en la fase estacionaria que cuando están en la fase exponencial de crecimiento62,68,69.
- Hidrofobicidad de la superficie celular:
Se ha demostrado que la hidrofobicidad de la superficie celular está relacionada con la adherencia de blastosporas de Candida a las células epiteliales humanas y a los materiales plásticos16.
Por otra parte, se ha sugerido que los cambios que se suscitan en las proteínas de la capa externa de la pared celular de C. albicans, son los responsables de las variaciones hidrofóbicas a hidrofílicas en esta especie70. Las células hidrofóbicas de C. albicans se unen difusamente y abundantemente a los tejidos del hospedero, en tanto que la unión de las células hidrofílicas se restringe a sitios específicos del hospedero. Las células hidrofílicas de C. albicans se unen a regiones con macrófagos, en contraste con las células hidrofóbicas, las cuales se unen a los tejidos en aquellos lugares del hospedero libres de macrófagos, por lo que las células hidrofílicas son más fáciles de remover del organismo por fagocitosis que las células hidrofóbicas, las cuales pueden colonizar el epitelio71,72,73.
La adherencia de las especies de Candida a las superficies plásticas es mediada por fuerzas de atracción de London-van der Walls (fuerzas hidrofóbicas y fuerzas electrostáticas). Asimismo, la habilidad de las especies de Candida para adherirse a las superficies de acrílico de las prótesis, puede conferirle a estos microorganísmos un acceso directo al hospedero humano74.
- Bacterias de la cavidad bucal:
Las bacterias pueden contribuir a la colonización y proliferación de especies de Candida en la cavidad bucal75. Un estudio realizado por Jenkinson y colaboradores76, puso en evidencia que la coagregación de Streptococcus sanguis, Streptococcus gordonii, Streptococcus oralis y Streptococcus anginosus con C. albicans se incrementa en ausencia de glucosa. También demostraron estos autores que, al someter a las blastosporas de Candida al calor o a la presencia de proteasas, se elimina la coagregación con S. sanguis y con S. gordonii.
En un estudio de data muy reciente, se ha comprobado que la adhesión de C. albicans a algunas especies de Streptococcus de la cavidad bucal, particularmente S. gordonii y S. sanguis, es promovida por la adsorción selectiva de las proteínas salivales ricas en prolina sobre la superficie celular de los cocos. Asimismo, se ha afirmado que, el reconocimiento selectivo de proteínas salivales adheridas a los cocos por parte de C. albicans, podría constituir un mecanismo para la colonización de esta especie en la cavidad bucal77.
- Hifas:
Existe un acuerdo entre la mayoría de los investigadores con respecto a que las hifas de C. albicans están asociadas con su capacidad invasiva22,69,78.
Se ha demostrado que existe una estrecha correlación entre la formación del tubo germinal y el incremento de la adherencia de C. albicans a las células epiteliales bucales, por lo que se ha sugerido desde entonces que este pudiera ser uno de los mecanismos relacionados con la virulencia por parte de las especies de Candida78. También existen evidencias de que durante la formación de las hifas, se producen proteinasas que ayudan a romper la integridad de la mucosa bucal79.
Varios factores regulan la transición de C. albicans de blastosporas a hifas. Estos incluyen: Temperatura y pH del medio de crecimiento, medios que contienen sustancias inductoras como suero, N-Acetil-D-Glucosamina, L-Prolina y etanol16.
Se ha demostrado "in vitro" que, temperaturas que oscilan entre 37°C y 40°C, pH entre 6,5 y 7,0, así como la presencia de una concentración inicial de blastosporas que no exceda de 106/ml., son escenciales para el crecimiento de las hifas al cabo de varias horas. También se ha demostrado que a pH 2,6, las células adherentes de C. albicans eran capaces de formar hifas, pero las células suspendidas de este hongo no tenían esa capacidad16.
Es importante resaltar que, la presencia de fuentes de carbono y nitrógeno son esenciales para la transformación de blastosporas a hifas en C. albicans16.
- Enzimas:
Estudios realizados por Barrett-Bee y colaboradores, revelaron que la Fosfolipasa A y la Lisofosfolipasa producidas por cepas de C. albicans firmemente adheridas a las células epiteliales bucales, eran las fosfolipasas con mayor actividad enzimática80.
Se ha sugerido que las hifas producidas por C. albicans pueden tener fosfolipasas, las cuales permiten su entrada a las células epiteliales del hospedero16. También se ha reportado que las fosfolipasas extracelulares y las proteasas ácidas pueden activarse a pH bajos66.
Se ha demostrado que, las proteinasas juegan un papel importante en la adherencia y en la invasión de Candida al epitelio bucal81. Por otra parte, se ha podido determinar que luego de la ingestión de blastosporas de Candida por parte de los macrófagos, el hongo sintetiza rápidamente proteinasas, seguido de una actividad proteolítica y de la formación de tubos germinales por parte de las blastosporas fagocitadas que conllevan a la destrucción de los macrófagos16.
CONCLUSIÓN.
Es conveniente destacar para que las membranas mucosas bucal, vaginal e intestinal normales, son capaces de soportar en forma comparativa grandes poblaciones de Candida sin que sufran ningún efecto aparente de enfermedad. Además, la frecuencia de estos microorganismos parece que aumenta con la edad. Sin embargo, cuando existe debilidad, desnutrición, alteración o ausencia de mecanismos de defensa normales del cuerpo (como en la Leucemia Aguda), o una notable descompensación en el balance normal de la superficie de la microbiota por la administración de antibióticos de amplio espectro, estos hongos pueden causar infecciones características que pueden ser bastante serias e incluso poner en peligro la vida del paciente. De allí que el Odontólogo no se limite solamente a conocer los aspectos inherentes al diagnóstico clínico de Candidiasis Bucal, sino que conozca los aspectos referentes a las características del hongo mayormente implicado en esta entidad.
REFERENCIAS BIBLIOGRÁFICAS:
1.-WEBB, B.C.; THOMAS, C.J.; WILLCOX, M.D.P.; HARRY, D.W.S.; KNOX, K.W. (1998): Candida-associated denture stomatitis. Aetiology and management: A review. Part 2. Oral diseases caused by Candida species. Aust Dent J. 43 (3): 160-166. [ Links ]
2.-BUDTZ-JORGENSEN, E. (1990): Etiology, pathogenesis, therapy and prophylaxis of oral yeasts infections. Acta Odontol Scand. 48: 61-69. [ Links ]
3.-SAMSON, J. (1990): Candidiosis buccales: Epidémiologie, diagnostic et traitement. Rev Mens Suisse Odontostomatol. 100: 548-559. [ Links ]
4.-CROCKETT, D.N.; O´GRADY, J.F.; READE, P.C. (1992): Candida species and Candida albicans morphotypes in erythematous candidiasis. Oral Surg Oral Med Oral Pathol. 73: 559-563. [ Links ]
5.-MATA, S. (1997):Cap. 26: Micosis. Clasificación. Micosis Superficiales. En NUÑEZ, M.J.; GOMEZ, M.J.; CARMONA, O. Microbiología Médica. Publicaciones U.C.V. 3ra Edición. pp. 579-611. [ Links ]
6.-CASAS RINCON, G. (1989): Micología General. Caracas. Universidad Central de Venezuela. Ediciones de la Biblioteca, pp. 243-247. [ Links ]
7.-HAY, R.J. (1986): Systemic candidiasis in heroin addicts. Brit Med J. 292: 1.096. [ Links ]
8.-BAKER, J. G.; SALKIN, I. F.; PINCUS, D.H.; D´AMATO, R.H. (1981): Candida paratropicalis, a new species of Candida. Mycotaxon. 13: 115-119. [ Links ]
9.-SHECTER, Y.; LANDAU, J.W.; DABROWA, N. (1972): Comparative electrophoresis and numerical taxonomy of some Candida species. Mycologia. 64: 841-853. [ Links ]
10.-SAEZ, H.; ANDRIEU, S. (1979): Étude mycologique comparée de Candida stellatoidea et Candida albicans. Ann Parasitol. 54: 555-565. [ Links ]
11.-MAGEE, B.B.; MAGEE, P.T. (1987): Electrophoretic karyotypes and chromosome numbers in Candida species. J Gen Microbiol. 133: 425-430. [ Links ]
12.-SNELL, R.G.; HERMANS, I.F.; WILKINS, R.J.; CORNER, B.E. (1987): Chromosomal variations in Candida albicans. Nucl Acid Res. 15: 3.625. [ Links ]
13.-MASON, M.M.; LASKER, B.A.; RIGGSBY, W.S. (1987): Molecular probe for identification of medically important Candida species and Torulopsis glabrata. J Clin Microbiol. 25: 563-566. [ Links ]
14.-MC GINNIS, M.R.; RINALDI, M.G. (1996): Selección de Hongos de Importancia Médica y algunos sinónimos Comunes y Nombres Obsoletos. Las Micosis en Venezuela. 10 (28): 49-53. [ Links ]
15.-VOLK, W.A.; BENJAMIN, D.C.; KADNER, R.J.; PARSONS, J.T. (1989): Microbiología Médica. Interamericana Mc Graw-Hill. 3ra Edición, pp. 533-560. [ Links ]
16.-.WEBB, B.C.; THOMAS, C.J.; WILLCOX, M.D.P.; HARRY, D.W.S.; KNOX, K.W. (1998): Candida-associated denture stomatitis. Aetiology and management: A review. Part 1. Factors influencing distribution of Candida species in the oral cavity. Aust Dent J. 43: 45-50. [ Links ]
17.-ODDS, F.C. (1994): Pathogenesis of Candida infections. J Am Acad Dermatol. 31: S2-S5. [ Links ]
18.-MACKENZIE, D.W.R. (1962): Serum tube identification of Candida albicans. J Clin Pathol. 15: 563-565. [ Links ]
19.-SHEPHERD, M.G. (1992): Fungi and parasites in the oral cavity. En SLOTS, J.; TAUBMAN, M.A. Contemporary Oral Microbiology and Immunology. St. Louis - U.S.A. Mosby -Year Book, Inc. 1 ed. pp. 373-376. [ Links ]
20.-GHANNOUM, M.A.; BURNS, G.R.; ABU ELTEEN, K. (1986): Experimental evidence for the role of lipids in adherence of Candida spp. to human buccal epithelial cells. Infect Immun. 54: 189-193. [ Links ]
21.-PIERCE, A.M.; PIERCE, H.D.; UNRAU, A.M.; OEHLSCHLAGER, A.C. (1978): Lipid composition and polyene antibiotic resistance of Candida albicans. Can J Biochem. 56: 135-142. [ Links ]
22.-CALDERONE, R.; BRAUN, P. (1991): Adherence and Receptor Relationships of Candida albicans. Microbiol Rev. 55(1): 1-20. [ Links ]
23.-ARNOLD, W.N. (1972): Localization of acid phosphatase and b -fructo-furanidase within yeast envelopes. J Bacteriol. 112: 1.346-1.352. [ Links ]
24.-PUGH, D.; CAWSON, R.A. (1979): The surface layer of Candida albicans. Microbios. 23: 19-23. [ Links ]
25.-RAM, S.P.; ROMANA, L.K.; SHEPHERD, M.G.; SULLIVAN, P.A. (1984): Exo (1® 3)-b -glucanase, autolysin and trehalase activities during yeast growth and germ-tube formation in Candida albicans. J Gen Microbiol. 130: 1.227-1.236. [ Links ]
26.-MOLINA, M.; CENAMOR, R.; NOMBELA, C. (1987): Exo-1,3-glucanase activity in Candida albicans: effect of the yeast to mycelium transformation. J Gen Microbiol. 133: 609-617. [ Links ]
27.-REISS, E.; STONE, S.H.; HASENCLEVER, H.F. (1974): Serological and cellular immune activity of peptidoglucomannan fractions on Candida albicans cells walls. Infect Immun. 9: 881-890. [ Links ]
28.-CHATTAWAY, F.W.; HOLMES, M.R.; BARLOW, A.J. (1968): Cell wall composition of the mycelial and blastospore forms of Candida albicans. J Gen Microbiol. 51: 367-376. [ Links ]
29.-CASSONE, A.; SIMONETTI, N.; STRIPPOLI, V. (1973): Ultrastructural changes in the cell wall during germ tube formation from blastospores of Candida albicans. J Gen Microbiol. 77: 417-426. [ Links ]
30.-HOWLETT, J.A.; SQUIER, C.A. (1980): Candida albicans ultrastructure: colonization and invasion of oral epithelium. Infect Immun. 29: 252-260. [ Links ]
31.-POULAIN, D.; TRONCHIN, G.; DUBREMETZ, J.F.; BIGUET, J. (1978): Ultrastructure of the cell wall of Candida albicans blastospores: study of its constituyent layers by the use of a cytochemical technique revelating polysaccharides. Ann Microbiol.129:141-153. [ Links ]
32.-CASSONE, A.; KERRIDGE, D.; GALE, E.F. (1979): Ultrastructural changes in the cell wall of Candida albicans following cessation of growth and their possible relationship to the development of polyene resistance. J Gen Microbiol. 110: 339-349. [ Links ]
33.-CASSONE, A.; MATTIA, E.; BOLDRINI, L. (1978): Agglutination of blastospores of Candida albicans by concanavalin A and its relationship with the distribution of mannan polymers and the ultrastructure of the cell wall. J Gen Microbiol. 105: 263-273. [ Links ]
34.-EVRON, R.; DREWE, J.A. (1984): Demostration of the polysaccharides in the cell wall of Candida albicans blastospores, using silver methenamine staining and a sequence of extraction procedures. Mycopathologia. 84: 141-150. [ Links ]
35.-NOTARIO, V. (1982): b -Glucanases from Candida albicans: purification, characterization and the nature of their attachment to cell wall components. J Gen Microbiol. 128: 747-759. [ Links ]
36.-HILENSKI, L.; NAIDER, F.; BECKER, J.M. (1986): Polyoxin D inhibits colloidal gold wheat germ agglutinin labelling of chitin in dimorphic forms of Candida albicans. J Gen Microbiol. 132: 1.441-1.451. [ Links ]
37.-REISS, E.; DE REPENTIGNY, L.; KUYKENDALL, R.J.; CARTER, A.W.; GALINDO, R., AUGER, P.; BRAGG, S.L., KAUFMAN, L. (1986): Monoclonal antibodies against Candida tropicalis mannan: antigen detection by enzyme immunoassay and immunofluorescence. J Clin Microbiol. 24: 796-802. [ Links ]
38.-HANSECLEVER, H.F.; MITCHELL, W. (1961): Antigenic studies of Candida. I. Observation of two antigenic groups in Candida albicans. J Bacteriol. 82: 570-573. [ Links ]
39.-PREUSSER, H.; ROSTER, H. (1979): Freeze-fracture studies of the plasmalemma of Candida albicans after treatment with econazole-nitrate. Sabouraudia. 17: 389-398. [ Links ]
40.-PESTI, M.; NOVAK, E.K.; FERENCZY, L., SVOBODA, A. (1981): Freeze fracture electron microscopical investigation of Candida albicans cells sensitive and resistant to nystatin. Sabouraudia. 19. 17-26. [ Links ]
41.-MARRIOTT, M.S. (1975): Isolation and chemical characterization of plasma membranes from the yeast and mycelial forms of Candida albicans. J Gen Microbiol. 86: 115-132. [ Links ]
42.-HUBBARD, M.J.; SURAIT, R.; SULLIVAN, P.A.; SHEPHERD, M.G. (1986): The isolation of plasma membrane and characterization of the plasma membrane ATPase from the yeast Candida albicans. Eur J Biochem. 154: 375-381. [ Links ]
43.-RAJASINGHAM, K.C.; CAWSON, R.A. (1978): Septal ultrastructure in Candida albicans. Acta Microbiol Pol. 27: 389-391. [ Links ]
44.-ODDS, F.C.; TRUJILLO-GONZALEZ, A. (1974): Acid phosphatase levels in the genus Candida and their application to the taxonomy and identification of pathogenic Candida species. Sabouraudia. 12: 287-294. [ Links ]
45.-ABDI, M.; DRUCKER, D. (1996): Phospholipid Profiles in Oral Yeast Candida. Archs Oral Biol. 41: 517-522. [ Links ]
46.-AKDENIZ, B.G.; SEN, B.H.; IGLENLI, T.; ATES, M.; DENIZCI, A. A.; ATILLA, G. (2000): Analysis of Candida albicans phenotypes in the oral cavity of renal transplant patients. J Clin Periodontol. 27 (Suppl 1): 92. [ Links ]
47.-NOLTE, W. (1986): Microbiología Odontológica. México. Ed. Interamericana. 4 ed. pp. 549-590. [ Links ]
48.-WILLIAMS, D.W.; LEWIS, M.A.O. (2000): Isolation and identification of Candida from the oral cavity. Oral Diseases. 6: 3-11. [ Links ]
49.-TOTTI, M.A.G.; JORGE, A.O.C., DOS SANTOS, E.B., DE ALMEIDA, O.P.; SCULLY C. (1996): Implantation of Candida albicans and other Candida species in the oral cavitiy of rats. J Oral Pathol Med. 25: 308-310. [ Links ]
50.-HOLMES, A.R.; O´SULLIVAN, J.M.; CANNON, R.D. (1999): Oral Colonization by Candida albicans. H I V Dent. Selected Abstracts from the 1999 American Society of Microbiology´s Conference on Candida and Candidiasis: 1-2. [ Links ]
51.-SYKES, L.M.; COOGAN, M.M. (1997): Yeast counts as a measure of host resistance in dental patients. J of the D A S A. 52: 19-23. [ Links ]
52.-SHEPHERD, M.G. (1986): The pathogenesis and host defence mechanisms of oral candidiosis. NZ Dent J. 82: 78-82. [ Links ]
53.-MARSH, P.; MARTIN, M. (1992): Oral Microbiology. London, Great Britain. Chapman & Hall. Third Edition. pp. 212-226. [ Links ]
54.-ROSSIE, K.; GUGGENHEIMER, J. (1997): Oral candidiasis: clinical manifestations, diagnosis and treatment. Pract Periodontics Aesthet Dent. 9 (6): 635-641. [ Links ]
55.-MATA, M. (1973): Candida albicans saprófito y patógeno de la cavidad bucal. Acta Odontológica Venezolana. Año XI (2-3): 663-680. [ Links ]
56.-LAZARDE L., J. (1989): Estudio sobre la transmisión de Candida albicans entre parejas conyugales. Trabajo de Ascenso. Facultad de Odontología, U.C.V. [ Links ]
57.-ARENDORF, T.M.; ALDRED, M.J.; MORRIS, D.B.; MURPHY, J.P. (1986): Intra-oral yeasts in an indigent South African community. J of the D A S A. 41: 327-330. [ Links ]
58.-ALKUMRU, H.N.; BEYDEMIR, K. (1992): The prevalence of Candida albicans in complete denture and removable partial denture wearers: a comparative study. J Marmara Univ Dent Fac. 1 (3): 218-222. [ Links ]
59.-ARENDORF, T.M.; WALKER, D.M. (1980): Candida albicans: its association with dentures, plaque and the oral mucosa. J of the D A S A. 35: 563-569. [ Links ]
60.-FEO, M. (1981): Supervivencia y desinfección de Candida albicans en el cepillo de dientes. Mycopathologia. 74: 125-134. [ Links ]
61.-SAMARANAYAKE, L.P.; MAC FARLANE, T.W. (1980): Factors affecting the in vitro adherence of Candida albicans to acrylic surfaces. Archs Oral Biol. 25: 611-615. [ Links ]
62.-MC COURTIE, J.; DOUGLAS, L.J. (1981): Relationship between cell surface composition of Candida albicans and adherence to acrylic after growth on different carbon sources. Infect Immun. 32: 1.234-1.241. [ Links ]
63.-NIKAWA, H.; HAMADA, T. (1990): Binding of salivary or serum proteins to Candida albicans in vitro. Archs Oral Biol. 35:571-573. [ Links ]
64.-EDGERTON, M.; SCANNAPIECO, F.A.; REDDY, M.S.; LEVINE, M.J. (1993): Human Submandibular-Sublingual Saliva Promotes Adhesion of Candida albicans to Polymethylmethacrylate. Infect Immun. 61 (6): 2.644-2.652. [ Links ]
65.-HOFFMANN, M.P.; HANDARIS, C.G. (1993): Analysis of Candida albicans adhesion to salivary mucin.Infect Immun. 61:1.940-1.949. [ Links ]
66.-SAMARANAYAKE, L.P.(1986): Nutritional factors and oral candidiosis. J Oral Pathol Med. 15: 61-65. [ Links ]
67.-BOUCHARA, J.; TRONCHIN, G.; ANNAIX, V.; ROBERT, R.; SENET, J.M. (1990): Presence of laminin receptors on Candida albicans germ tubes. Infect Immun. 58: 48-54. [ Links ]
68.-KING, R.D.; LEE, J.C.; MORRIS, A.L. (1980): Adherence of Candida albicans and other Candida species to mucosal epithelial cells. Infect Immun. 27: 667-674. [ Links ]
69.-CUTLER, J.E.; BRAWNER, D.L.; HAZEN, K.C.; JUTILA, M.A. (1990): Characteristics of Candida albicans adherence to mouse tissues. Infect Immun. 58: 1.962-1.980. [ Links ]
70.-HAZEN, K.L.; LAY, J.L.; HAZEN, B.W.; FU, R.C.G.; MURTHY, S. (1990): Partial biochemical characterization of cell surface hydrophobicity and hydrophilicity of Candida albicans. Infect Immun. 58: 3.469-3.476. [ Links ]
71.-HAZEN, B.W.; HAZEN, K.L. (1988): Dynamic expression of cell surface hydrophobicity during initial cell growth and before germ tube formation of Candida albicans. Infect Immun. 56: 2.521-2.525. [ Links ]
72.-HAZEN, K.L. (1989): Participation of yeast cell surface hydrophobicity in adherence of Candida albicans to human epithelial cells. Infect Immun. 57: 1.894-1.900. [ Links ]
73.-HAZEN, K.L.; BRAWNER, D.L.; RIESSELMAN, M.H.; JUTILA, M.A.; CUTLER, J.E. (1991): Differential adherence to hydrophobic and hydrophilic Candida albicans yeast cells to mouse tissues. Infect Immun. 59: 907-912. [ Links ]
74.-KLOTZ, S.A.; DRUTZ, D.J.; ZAJIC, J.E. (1985): Factors governing adherence of Candida species to plastic surfaces. Infect Immun. 50: 97-101. [ Links ]
75.-HSU, L.Y.; MINAH, G.E.; PETERSON, D.E. (1990): Coaggregation of oral Candida isolates with bacteria from bone marrow transplant. recipients. J Clin Microbiol. 28: 2.621-2.626. [ Links ]
76.-JENKINSON, H.F.; LALA, H.C.; SHEPHERD, M.G. (1990): Coaggregation of Streptococcus sanguis and other streptococci with Candida albicans. Infect Immun. 58: 1.429-1.436. [ Links ]
77.-O´SULLIVAN, J.M.; JENKINSON, H.F.; CANNON, R.D. (2000): Adhesion of Candida albicans to oral streptococci is promoted by selective adsorption of salivary proteins to the streptococcal cell surface. Microbiology. 146 (Pt 1): 41-48. [ Links ]
78.-KIMURA, L.H.; PEARSALL, N.N. (1980): Relationship between germination of Candida albicans and increased adherence to human buccal epithelial cells. Infect Immun. 28: 464-468. [ Links ]
79.-CUTLER, J.E. (1991): Putative virulence factors of Candida albicans. Ann Rev Microbiol. 45: 187-218. [ Links ]
80.-BARRETT-BEE, K.; HAYES, Y, WILSON, R.G.; RYLEY, J.F. (1985): A comparision of phospholipase activity, cellular adherence and pathogenicity of yeasts. J Gen Microbiol. 131: 1.217-1.221. [ Links ]
81.-BORG, M.; RUCHEL, R. (1988): Expression of extracellular acid proteinase by proteolytic Candida spp. during experimental infection of the oral mucosa. Infect Immun. 56: 626-631. [ Links ]
