










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa Odontológica Venezolana
versión impresa ISSN 0001-6365
Acta odontol. venez v.46 n.3 Caracas dic. 2008
Aspectos celulares y moleculares de las células madres involucrados en la regeneración de tejidos con aplicaciones en la práctica clínica odontológica
Juan Carlos Munévar Niño, Andrea del Pilar Becerra Calixto, Claudia Bermúdez Olaya
1 Od. MSc, D.E.A. Coordinador Cultivo celular y Regeneración Tisular(*),
2 BCl, Investigador Asistente*,
3 Estudiante(*),
*Instituto Unidad de Investigación Básica Oral (U.I.B.O). Facultad de Odontología. Universidad El Bosque, Bogotá Colombia. (Transversal 9A Bis No. 132 - 55) Correspondencia: munevarjuan@unbosque.edu.co
Abstract:
The medicine and dentistry entering a new era in which the news therapy approaches like genetic therapy, cellular therapy, engineering tissue and medicine regenerative they extended the arsenal of possibilities for our patients. The Stem cells in the field of the regenerative medicine, by its characteristics of self renewal, expansion and differentiation, have shown to be an important alternative for the treatment of pathologies and to alterations in the teeth and periodontal structures.
The tissue regeneration it implies the replacement of weaves affected with identical cells that can be generated from the stimulation of Mesenchymal Stem Cells (MSCs) by different involved molecular and cellular mechanisms in the dental morphogenetic; for that reason different options look for to facilitate the use of MSCs like clinical treatment of periodontal diseases and functional oral injury.
Key words: Adult Stem cells, Engineering Tissue, Oral regeneration
Resumen:
La medicina y odontología clínica están entrando en una nueva era en la cual los nuevos enfoques terapéuticos como la terapia génica, la terapia celular, la ingeniería tisular y la medicina regenerativa ampliaran el arsenal de posibilidades para nuestros pacientes. Las células Madres en el campo de la medicina regenerativa, por sus características de autorrenovación, proliferación y diferenciación, han mostrado ser una importante alternativa para el tratamiento de patologías y alteraciones en los dientes y estructuras periodontales.
La regeneración tisular implica el reemplazo de tejidos afectados con células idénticas que pueden ser generadas a partir de la estimulación de células Madres mesenquimatosas (MSCs) por diferentes mecanismos moleculares y celulares involucrados en la morfogénesis dental; por ello se buscan diferentes opciones para facilitar la utilización de MSCs como tratamiento clínico de enfermedades periodontales y traumas bucales funcionales.
La estandarización de protocolos para la obtención de ASCs autólogas de diferentes tejidos, eliminará problemas de rechazo inmunológico, permitiendo una completa regeneración funcional de los tejidos de la cavidad bucal.
Palabras Clave: Células Madres Adultas; Ingeniería de Tejidos; Regeneración oral.
Recibido para arbitraje:10/08/2006 Aceptado para publicación: 08/05/2007
Durante décadas los tratamientos para patologías y alteraciones en los dientes y estructuras periodontales han sido un tema de intensa investigación. En efecto, las enfermedades dentales, periodontales y craneofaciales además de los traumas se presentan de manera significativa en la práctica clínica odontológica. Un reciente estudio de World Oral Health en el 2003, reveló una cifra de aproximadamente cinco millones de personas en todo el mundo que presentaban alguna patología oral (1), mientras que otro estudio reporta más de 1.4 millones de casos, en los cuales alrededor del 97% d los dientes tratados permanecen funcionales por un periodo de 8 años (2). Por estás razones, en los últimos años, se han buscado nuevas opciones para el tratamiento de enfermedades dentales, periodontales y craneofaciales que generan secuelas estructurales y fisiológicas irreversibles en los pacientes (3,4).
El remoto origen de esta fascinación se remonta a la antigua Grecia. En la mitología griega, la hidra de múltiples cabezas casi derrota a Herácles pues le crecían dos nuevas cabezas por cada una que cortara el héroe y el hígado de Prometeo encadenado devorado por un águila hambrienta cada noche, se regeneraba a la mañana siguiente. Aristóteles (384 - 322 a.c.), observaba como se regeneraban las colas de lagartos y serpientes, así como los ojos de las golondrinas (5). En el siglo XVIII científicos como Abraham Trembley, Charles Bonnet, Peter Simon Pallas, y Lázaro Spallanzani; descubrieron notables habilidades de regeneración en una variedad de organismos como las hidras, los gusanos de tierra, los caracoles, las ranas premetamórficas, lagartijas y salamandras. Por esas razones, en el transcurso del siglo XIX y parte del siglo XX la investigación en regeneración tisular se enfocó primordialmente en la fenomenología de la regeneración y sus fundamentos celulares (6).
Hace unos años se pensaba que los tejidos humanos solo podían tratarse quirúrgicamente mediante injertos, por transplantes procedentes de donantes, con la aplicación de biomateriales sintéticos o mediante dispositivos artificiales. Era difícil vislumbrar que se diseñarían tejidos y órganos sintetizados en el laboratorio como opción para el tratamiento para el alivio del sufrimiento causado por el daño irreparable de los tejidos (4). Por ende, surgió la ingeniería de tejidos para diseñar estrategias terapéuticas novedosas que permitan sintetizar in vitro análogos de tejidos para el tratamiento de patologías con secuelas irreversibles y así mismo restituir ad integrum la estructura anatómica y la función tisular de los órganos afectados (1).
La medicina y odontología clínica están entrando en una nueva era en la cual los nuevos enfoques terapéuticos como la terapia génica, la terapia celular, la ingeniería tisular y la medicina regenerativa ampliaran el arsenal de posibilidades para nuestros pacientes. Una línea de investigación fundamental en ingeniería tisular y medicina regenerativa son las células madres (7).
Las células madres se han definido como células clonogénicas, con un amplio potencial de auto renovación (definida como la capacidad de generar al menos una célula hija con características similares a la célula de origen, manteniéndose al mismo tiempo en un estado indiferenciado), así como la elevada capacidad de proliferación (posibilidad de la célula para dividirse sin cambiar su fenotipo celular indiferenciado) y su potencial de diferenciación (potencial para modificar el fenotipo de la célula de origen en distintos tipos celulares diferentes al tejido embrionario original en varias líneas celulares como medula ósea, sangre periférica, cerebro, piel, pulpa dental y ligamento periodontal, entre otros) (8,9,10).
La biología, las propiedades, clasificación, fenotipos, mecanismos celulares y/o moleculares, bioquímicos, de señalización y las probables aplicaciones terapéuticas de las células madres se han estudiado durante décadas. Podemos clasificarlas según su capacidad de proliferación y diferenciación en totipotenciales (células que tienen el potencial de dar origen a un organismo completo incluyendo el tejido germinal), pluripotenciales (células que pueden dar origen a células de las tres capas germinativas: ectodermo, mesodermo y endodermo); y multipotenciales (son células comprometidas en una línea celular específica y dan origen a células de un órgano o tejido particular) (11,12).
CÉLULAS MADRES ADULTAS
Las células madres multipotenciales, se consideran órgano-especificas y se localizan en tejidos embrionarios y fetales además en tejidos completamente formados de los organismos adultos. Se denominan Células Madres Adultas (ASCs), pues originan células progenitoras y precursoras relacionadas solo con una de las tres capas embrionarias, sin embargo, esta definición está sujeta a controversias. En efecto, en recientes estudios, se sugiere que las ASCs, pueden diferenciarse en otras líneas celulares distintas de las pertenecientes a su capa germinativa de origen, como ejemplo podemos mencionar a las células Madres Hematopoyéticas (HSCs), aisladas de medula ósea o bien de sangre de cordón umbilical humano, que además, de originar células sanguíneas, poseen in vitro el potencial de generar células musculares, neuronales y adiposas (6,10,13,14).
Las ASCs se han caracterizado en distintos tejidos, lo que genera una excelente opción para su posterior utilización terapéutica. En efecto, las ASCs no despiertan los dilemas éticos generados por las Células Madres Embrionarias (ESCs) responsables de grandes controversias científicas, académicas, religiosas y éticas a nivel mundial (6). Las propiedades únicas de las ASC, como el mantenimiento del fenotipo indiferenciado concomitante con la habilidad de autorreplicación por largos periodos de tiempo, la elevada capacidad de proliferación y su potencial de plasticidad o transdiferenciación, permitirán el uso de este tipo de células como alternativa de tratamiento para modular la regeneración tisular de epitelio, miocardio, médula ósea, neuronas y células de la neuroglía, células ? de páncreas, tejidos dentales, pulpares y periodontales, entre otros, para tratar enfermedades como infartos, diabetes y síndromes degenerativos (3).
Los diferentes tipos de ASCs multipotenciales identificados, con las características mencionadas anteriormente expresan un fenotipo hematopoyético (Hemayopoietic Stem cells [HSCs]), mesenquimatoso (Mesenchymal Stem cells [MSCs]), estroma de médula ósea (Bone Marrow Stromal Stem Cells [BMSSCs]) pulpa dental (Dental Pulp Stem Cells [DPSCs]), hepáticas, entre otras (15).
Células Madres Hematopoyéticas
Las HSCs han sido las más estudiadas e identificadas tanto in vitro como in vivo por varios laboratorios y utilizadas clínicamente desde hace más de 50 años. El transplante alógeno de progenitores hematopoyéticos ha demostrado definitivamente que posee células madres multipotenciales hematopoyéticas en la médula ósea y en la sangre periférica (16). Además del potencial hematopoyético, diversos trabajos recientes indican que las HSCs bajo ciertas circunstancias presentan mayor plasticidad por la capacidad de diferenciarse a tejidos derivados de distintas capas embrionarias (17). Las células madres hematopoyéticas de médula ósea y de sangre periférica son capaces de contribuir a la angiogénesis y vasculogénesis in vivo de tal forma que las células CD34+ no sólo contienen progenitores hematopoyéticos sino también células progenitoras endoteliales (17). Hoy en día se acepta que existe un progenitor común endotelial y hematopoyético conocido como hemangioblasto, concepto sustentado por los hallazgos relacionados con el potencial endotelial de las células madres hematopoyéticas de la médula ósea (18). Estudios recientemente publicados apoyan la capacidad de las HSCs de diferenciarse en células de músculo cardíaco al ser estimulados in vitro. El grupo de Orlic y Anversa han demostrado en un modelo de infarto de miocardio murino, como la inyección de células de médula ósea Lin- y c-kit+ CD34+ (fenotipo característico de HSCs) en el área lesionada del corazón colonizan más de la mitad del área infartada. Estas posibles HSCs adquirieron un fenotipo característico de células de miocardio y contribuyen a la mejora y supervivencia de los animales (18). La contribución de las células madres adultas a la regeneración cardiaca ha sido sugerida en modelos de transplante cardíaco en humanos. El grupo de Lagasse y colaboradores han demostrado que HSCs de médula ósea con el fenotipo Lin-, c-kit+, Thy-1, Sca-1 son capaces de hepatocitos murinos en un modelo de daño hepático fulminante (19). El potencial de las HSCs para adquirir características de músculo esquelético, neuronas adultas así como células de la glía, y de contribuir a otros tejidos como el epitelio pulmonar, gastrointestinal, renal o a la piel se ha descrito recientemente in vivo (20).
Células Madres Mesenquimatosas
Las células Madres Mesenquimatosas (MSCs) humanas, se han aislado principalmente del estroma de la médula ósea, tejido adiposo, sangre, gelatina de Wharthon de cordón umbilical humano, dermis y piel. Las MSCs presentan un gran potencial de generar cartílago, hueso, músculo, tendón, ligamento y tejido adiposo, entre otros (13,14, 21).
Una célula es identificada por consenso internacional como célula madre o Stem cell cuando se detecta la expresión de varios marcadores fenotípicos específicos que reflejan la presencia (+) y/o ausencia (-) de antígenos característicos en esa(s) célula(s) (6). En efecto, estudios realizados han demostrado que las MSCs se caracterizan por expresar marcadores positivos para CD29, CD44, CD59, CD73, CD90, CD105, CD117 (c-kit), CD166 y STRO-1 y negativos para CD11a, CD11b, CD14, CD33, CD34, CD28, CD45, como fenotipo característico de HSCs (9, 10, 20).
PAPEL DE LAS CÉLULAS MADRES ADULTAS EN EL DESARROLLO CRANEOFACIAL.
En general, durante el desarrollo craneofacial la formación ósea se puede dar por dos vías: la osificación intramembranosa y la osificación endocondral. La osificación intramembranosa, se caracteriza por ocurrir a nivel embrionario durante la formación y desarrollo de la mayor parte del esqueleto craneofacial. La osificación endocondral es más característica del esqueleto postcraneal, también ocurre en la base del cráneo y en la articulación temporomandibular (22). El cartílago se forma a partir de la condensación de células mesenquimatosas seguido de un proceso de maduración condrocitaria. Durante este proceso, el cartílago es invadido por vasos sanguíneos de donde las células osteoprogenitoras se diferencian para sustituirlo por hueso (23). Una vez las estructuras óseas están formadas, la arquitectura morfológica del hueso es regulada por osteoclastos y osteoblastos cruciales para un constante proceso de remodelado óseo (24).
El desarrollo y crecimiento sutural podría considerarse como una forma especializada de crecimiento de hueso intramembranoso en donde las células óseas se originan a partir de MSCs localizadas en el blastema sutural bajo influencia de la duramadre subyacente y el cartílago capsular nasal (25). El crecimiento de las suturas óseas en desarrollo depende del mantenimiento del balance entre proliferación y diferenciación de las células madres a células óseas para sintetizar las moléculas de la matriz extracelular del tejido óseo y su posterior biomineralización. De este modo, la población de células madres se mantiene hasta que el crecimiento craneal este completo (24,26). Así mismo, el desarrollo y crecimiento del cóndilo mandibular (caracterizado por la presencia de cartílago) de la articulación temporomandibular, es una forma especializada de crecimiento esquelético intramembranoso asociado con células madres precursoras dentro de la capa precondroblástica o proliferativa (27,28).
Una vez formado el hueso, la resorción y aposición ósea están moduladas por el periostio, el cual depende de la generación de osteoblastos y osteoclastos derivados de poblaciones de células madres progenitoras en esta capa osteogénica (23).
Células Madres Adultas. Formación y erupción dental.
En el transcurso del proceso de erupción y formación dental la diferenciación de células Madres osteoblásticas y osteoclasticas es absolutamente necesaria para controlar la sincronía de las fases a lo largo de la vía de erupción y su respectivo movimiento dental. Para crear la vía de erupción por resorción alveolar ósea activa es fundamental el reclutamiento de osteoclastos (29). La diferenciación y proliferación de preosteoclastos pueden ser activadas por el factor estimulante de colonias 1 (CSF-1), el ligando del factor nuclear kappa ? (RANKL) y la osteoprotegerina (OPG) (28).
La activación de factores de transcripción específicos por medio de citoquinas y factores de crecimiento, estimulan la diferenciación a osteoclastos localizados en el periostio periférico al folículo dental que permite inducir la osteogénesis durante la formación del periodonto. Para la activación del osteoclasto, debe actuar el ligando del receptor activador del factor nuclear kappa B (RANKL) (localizado en osteoblastos y células del estroma de la médula ósea) acoplado al receptor activador del factor nuclear kappa B (RANK) (localizado en preosteoclastos). La osteoclastogénesis está regulada por la osteoprotegerina (OPG) que al unirse con RANKL neutraliza el proceso de actividad osteoclástica (30).
Durante el movimiento dental, la diferenciación y activación osteoclástica es regulada en mayor parte por RANKL posiblemente producido en células estromales y osteoblastos de tejido periodontal. Según Shiotani y colaboradores en el 2001, la molécula RANKL se localiza en tejidos periodontales durante el movimiento experimental de molares en ratas. Esa expresión de RANKL se detectó en osteoblastos, osteocitos y fibroblastos, principalmente en el citoplasma, retículo endoplasmático rugoso, y membrana plasmática (31). El RANKL, se expreso en los osteoblastos del tejido periodontal, durante la erupción dental y el movimiento ortodóntico. De este modo, la erupción dental y el movimiento ortodóntico requieren una compresión y retracción del periodonto lo cual estimula la proliferación y diferenciación de las poblaciones de células madres lo que ocasiona una resorción ósea y regeneración en ese medio ambiente local (31). Estos hallazgos sugieren un posible papel de RANKL en la diferenciación osteogénica de las células madres.
Se ha demostrado in vitro que el comportamiento de las células Madres adultas está modulado por factores solubles liberados por las células óseas (16). Heino y colaboradores utilizaron la línea celular MLO-Y4 para estudiar la proliferación, diferenciación y osteoformación a partir de MSCs de médula ósea a osteocitos (32). Se observó un incremento cuatro veces mayor de formación ósea y dos veces mayor en la proliferación de MSCs, al utilizarse el medio de cultivo condicionado con la línea celular osteocitica MLO-Y4. Lo anterior indica que el medio de cultivo de osteocitos estimula la proliferación de las MSCs y su diferenciación a osteoblastos. Sin embargo, se desconocen los factores en los osteocitos que condicionan el medio para inducir a la diferenciación; se presume que las señales entre osteocitos y MSCs ejercen una actividad estimulante de distinta naturaleza desempeñando un papel importante en el control de la formación ósea.
La proliferación y diferenciación de osteoblastos y osteoclastos a partir de células Madres progenitoras mesenquimatosas y/o hematopoyéticas respectivamente son importantes en el crecimiento y el desarrollo postnatal controlado por factores solubles locales como; morfogenes, factores de crecimiento, citocinas, neuropéptidos y hormonas. Los factores de crecimiento estimulan la proliferación y diferenciación celular mediante la activación de receptores específicos en células blanco. Muchos de los factores de crecimiento se encuentran en forma activa asumiendo diversas funciones en diferentes periodos durante el crecimiento y desarrollo de órganos y tejidos corporales (33,34). La vía de señalización del factor de crecimiento transformante (TGF ) ocupa una posición central dentro de las vías de señalización desencadenadas por factores solubles que controlan el crecimiento, diferenciación y destino en las células (35). El TGF induce las células alterando la expresión de los genes como el c-myc y junB controlando la proliferación celular. Warner y colaboradores en el 2003, demostraron que la familia del TGF son reguladores críticos en el desarrollo orofacial de los mamíferos debido a diferencias de expresión por gradientes espaciotemporales en sitios tisulares específicos controlando la proliferación celular y crecimiento del tejido (36). Hormonas como la de hormona de crecimiento, también están involucradas en la formación ósea por medio de la estimulación de precursores óseos o células madres para su proliferación, seguido de una expansión clonal y promoción de la diferenciación hacia linajes osteogénicos (37).
Moléculas de señalización implicadas en la diferenciación y regeneración dental.
Los morfogenes, son señales secretadas a nivel extracelular, dirigiendo la morfogénesis durante las interacciones epitelio- mesénquima. Las vías de señalización morfogenética incluyen cinco clases de genes altamente conservados durante la evolución: proteínas morfogenéticas óseas (BMP), factores de crecimiento fibroblástico (FGFs), proteínas internas wingless (Wnts), proteínas Hedgehog (Hhs), y las moléculas de la familia del factor de necrosis tumoral (TNF) (29,38). Estas familias de genes están principalmente involucradas durante el inicio de la morfogénesis y cito diferenciación (39).
La familia de las BMP, relacionada con la formación y erupción dental, están formadas por seis diferentes clases (BMP2 a BMP7) que son co-expresadas según gradientes temporoespaciales (40). Diez miembros de la familia de las BMP's (BMP2, BMP4, BMP6, BMP7, BMP8), factor de crecimiento de diferenciación (GDF) 1, GDF5, GDF6, GDF7, GDF11 y factor neurotrófico derivado de línea celular glial (GDNF); han sido clonados de la pulpa de incisivos en ratas (41). La BMP4 sintetizadas por el epitelio, inducen al mesénquima hacia un linaje odontogénico, mientras que la BMP2, BMP4 y BMP7 estimulan la expresión sostenida de enamelina e influencian a las células epiteliales y mesenquimatosas a inducir posteriormente la morfogénesis del epitelio (42). Así mismo, la BMP2, BMP4, BMP6, BMP7 y GDF11 se expresan durante la diferenciación de los odontoblastos; mientras que la BMP4 y BMP5 durante la diferenciación de los ameloblastos (43,44). Las vías complejas de señalización de las BMP's están reguladas por la interacción con sitios extracelulares de los receptores transmembranales específicos (45). Existen antagonistas de BMP's como noggin, cordina y folistatina, que modulan la bioactividad de los morfogenes (46). En las células de la pulpa dental, se expresan receptores transmembranales BMP tipo I y II con actividad serina treonina kinasa, que intervienen en los procesos de comunicación e interacción celular epitelio - mesénquima; estos permiten que las señales de las BMP's sean transducidas desde la membrana plasmática hacia el núcleo por medio de proteínas SMADS, receptor activador de Smads (R-SMADS), mediador común de Smads (Co- SMADS) e inhibidor de Smads (I- SMADS) (47).
Los morfogenes son señales que funcionan como factores de diferenciación y crecimiento implicados en el establecimiento de patrones específicos en la arquitectura de órganos y tejidos. Las BMP's originalmente fueron aisladas de matrices de hueso descalcificado; la utilización de BMP2 recombinante humana estimula la diferenciación in vitro de células pulpares indiferenciadas en odontoblastos en cultivos en monocapa y órganotipicos establecidos en matrices 3D (tridimensionales) (48). La proteína recombinante humana BMP2, BMP4 y GDF11 en matrices de gel de agarosa, y TGF 1 asociado con una fracción inactiva soluble de EDTA estimula diferenciación en odontoblastos de cultivos de células de papila dental (49). Efectos similares han sido demostrados en cultivos de dientes con TGF 1-3 y BMP7 (50,51). La proteína recombinante humana BMP2, BMP4 y BMP7 inducen a la formación de dentina reparativa/ regenerativa in vivo (52,53,54). El factor de crecimiento insulínico recombinante humano I en membranas de colágeno induce a una completa restauración y formación de dentina y dentina tubular (55).
Aún no esta totalmente clara la regulación y la transición abrupta de células madres de un estado quiescente a uno activado en términos de proliferación, migración, diferenciación y secreción de la matriz, después de un daño en el tejido pulpar; el control de los mecanismos moleculares de estos diferentes morfogenes es necesario para elucidar el buen uso terapéutico en endodoncia regenerativa.
CARACTERIZACIÓN DE CELULAS MADRES EN EL COMPLEJO PULPODENTINAL
Una posible fuente de MSCs se encuentra en la pulpa dental, este es un tejido conectivo de baja vascularidad rodeado por dentina, conformado por una población heterogénea de células como: periodontoblastos, fibroblastos, células estromales, células endoteliales y perivasculares, células nerviosas, entre otras; estas células mantienen la homeótasis de los diferentes tejidos dentinales mineralizados. La mayoría de las células pulpares son postmitóticas; sin embargo, algunas de estas células aún se dividen y forman capas de nuevas células pulpares con habilidad de diferenciación a odontoblastos y formación de dentina (56). Estas células, junto con los vasos sanguíneos se encuentran en matrices extracelulares creando un microambiente ideal para permitir procesos de reparación.
Diferentes estudios, han buscado definir el fenotipo de células madres mesenquimales de pulpa dental (DP-MSC), el crecimiento in vitro en comparación con MSCs de médula, y su plasticidad in vitro por lo menos en tres tipos de células: osteoblastos, condroblastos y adipositos (57). Gronthos y col; caracterizaron estas células por medio de marcadores específicos de MSCs y observaron su capacidad de autoregeneración, diferenciación a múltiples linajes y su capacidad clonogénica; hallando DPSCs capaces de formar dentina asociada con tejido pulpar in vivo (58). Iohara y colaboradores, por medio de la expresión de mRNA de Dentina sialofosfoproteína (Dspp) y metaloproteínasas de la matriz 20 (MM20) confirmaron la diferenciación de DPSCs en odontoblastos al ser estimuladas por proteínas morfogenéticas óseas (59).
Otros estudios como los publicados por Shi y col. Muestran la expresión de marcadores de MSCs como STRO-1 y CD46 en ligamento periodontal además de una diferenciación de este tejido en odontoblastos, cementoblastos, adipositos y células productoras de colágeno in vitro; al ser transplantadas en ratones inmunosuprimidos muestran la formación de estructuras dentales que pueden contribuir a una eventual reparación (60).
En el Instituto Unidad de Investigación Básica Oral (U.I.B.O) de la Universidad El Bosque estamos realizando un proyecto de investigación con el propósito de determinar la presencia de células madres en muestras de pulpa dental humana. La caracterización de estás células la realizamos identificando la expresión de los marcadores CD 117, FGFR3, CD 90, STRO-1, CD34, CD44 y CD45 mediante inmunohistoquímica y citometría de flujo. Los resultados previos obtenidos revelan un inmunomarcaje del 13.2 % de células CD117 +, mientras un porcentaje del 17.5 % de células FGFR3+ (61). Estos resultados demuestran la presencia de células precursoras en la pulpa dental, probablemente de la línea hematopoyética o mesenquimatosa. Se requieren estudios de caracterización adicionales de las células que expresan el fenotipo FGFR 3+ / CD 117+ con otros marcadores específicos de células Madres como STRO-1, CD-90, SH2, SH3, flujo, técnicas de biología molecular como RT-PCR para analizar el nivel de expresión de ARNm de genes propios de células madres mesenquimales (hMSC's) o células madres Hematopoyéticas (hHSC's).
Estos trabajos de investigación son el pilar fundamental para desarrollar nuevos tratamientos en el futuro basados en células madres. En efecto, existen varios ensayos clínicos con terapia celular basados en células madres que en este momento se efectúan en sujetos humanos. Estas estrategias terapéuticas novedosas tendrán aplicación en la práctica clínica odontológica.
INGENIERÍA DE TEJIDOS Y SU FUTURO EN LA ODONTOLOGÍA
El diente y las estructuras periodontales, son importantes órganos del complejo craneofacial, por esto, las enfermedades dentales y periodontales se consideran una amenaza que contribuyen a la pérdida dental; los tratamientos utilizados para estas enfermedades, no restauran completamente el diente después de la patología o el trauma sufrido, produciendo anquilosis, reabsorción de la raíz y pérdida del diente; la incidencia de agénesis dental congénita en niños y pérdida de dientes en la población adulta se ha convertido en un gran problema de salud pública en los últimos años (62). La utilización de diferentes estrategias para el reemplazo de los dientes afectados, es uno de los objetivos de la ingeniería de tejidos en el campo odontológico (63).
En las últimas décadas, se ha investigado el potencial de ASCs para formar estructuras dentales funcionales (Tabla 1); el laboratorio Sharpe (64), reemplazó células mesenquimatosas adultas con células mesenquimatosas generadas de células madres no dentales (MSCs) en ratones con epitelio bucal embrionario; la recombinación entre MSCs y epitelio bucal embrionario estimularon odontogénesis en ESCs, NSCs, BMSSCs además de la expresión de genes odontogénicos. Estas células recombinadas fueron transferidas en cápsulas renales, observándose el desarrollo de estructuras dentales y asociaciones óseas. Estos resultados proveen un significante avance hacia la creación de primordios de dientes artificiales originados de cultivos celulares que pueden ser usados en reemplazo a dientes faltantes, por medio del transplante dentro de la cavidad oral adulta (65).
Tabla 1. Potencial de ASC para diferenciarse en distintas estructuras y tipos de células craneofaciales (3).
| Origen de Células Madres Adultas | Estructuras o tipos de células diferenciadas | Laboratorios |
| Médula ósea | Estructura dental y hueso | Paul T. Sharpe(60) |
| Pulpa dental | Dentina, odontoblastos, pulpa dental, estructuras de cemento y ligamento periodontal | Songtao Shi (63) |
| PDL | Estructuras de cemento y PDL | Songtao Shi (66) |
| Brote dental | Estructuras dentales | Pamela C. Yelick (67) |
| Médula ósea | Células hematopoyéticas osteoblastos, osteocitos | Edwin M. Horwitzf (68) |
PDL. Ligamento periodontal
Las MSC o células progenitoras multipotentes adultas (MAPC), también pueden diferenciarse en fenotipos celulares que permiten la restauración de las células involucradas en el daño dental. Múltiples estudios buscan diferentes opciones para facilitar la utilización de MSCs como tratamiento clínico de enfermedades periodontales y traumas orales (Figura 1), partiendo de estudios realizados que se enfocan en la regeneración y reparación del tejido de cavidad oral. Co-cultivos de MSCs y ligamento periodontal han mostrado un incremento en la población de MSCs expresando osteocalcina, osteopontina y una disminución en la expresión de sialoproteína ósea, característico del ligamento periodontal, estos hallazgos indican, que el contacto de factores celulares de ligamento periodontal induce a las MSCs a poseer características de este tejido (65).
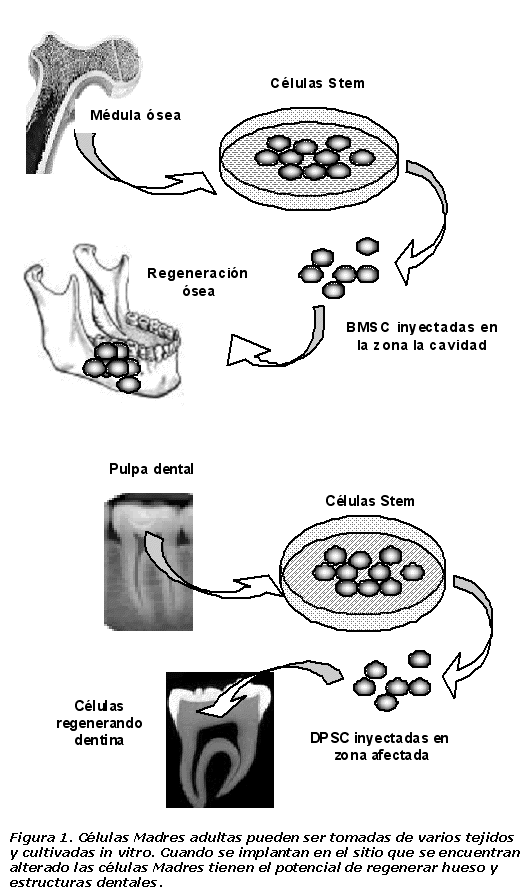
La reparación del tejido periodontal por medio de células progenitoras permite la formación de fibroblastos, osteoblastos y cementoblastos. En la investigación realizada por Shi y col, aislaron células madres de ligamento periodontal (PDLSCs) de 25 pacientes, de los cuales extrajeron sus molares, mostrándose la expresión de marcadores de MSC en las células aisladas, como son STRO-1 y CD146/ MUC18 por medio de citometría de flujo (69). Dentro de condiciones definidas en cultivo, las PDLSC se han diferenciado en cementoblastos, adipositos y células productoras de colágeno que muestran capacidad para regenerar el cemento al ser transplantadas en ratones inmunocomprometidos, contribuyendo a la reparación del tejido periodontal (70).
En tejidos duros de la cavidad oral, donde no se pensaba hallar ASC, se encontraron células clonogénicas con gran capacidad de proliferación (71), adicionalmente, in vivo el transplante de estas células, permiten la generación de tejido duro. Se han encontrado también, células madres en pulpa (PDSC), estas tienen alto poder proliferativo y se obtienen de la disociación enzimática de la pulpa dental humana, que esporádicamente, in vitro puede producir calcificación; y transplantadas en ratones inmunosuprimidos forman fibras de colágeno y mineralizaciones (70).
PERSPECTIVAS
La regeneración dental y periodontal, es un proceso más complejo que la regeneración de otro tipo de tejidos como músculo, sistema nervioso y hematopoyético. La regeneración tisular implica el reemplazo de tejidos afectados con células idénticas que pueden ser generadas a partir de la estimulación de MSC's. Debido a su capacidad de transdiferenciación en condiciones apropiadas, las células madres aisladas del complejo craneofacial podrían ser candidatas para el diseño de estrategias terapéuticas novedosas con aplicaciones en la práctica clínica (72). Por eso se estudian los mecanismos moleculares y celulares involucrados en la morfogénesis dental, caracterizados por una serie de interacciones reciprocas entre el epitelio dental y el mesenquima subyacente, proceso biológico que involucra numerosas moléculas, vías de señalización y factores de crecimiento mediadores de estas interacciones (73). Estudios in vitro e in vivo enfocados en la comprensión del papel que desempeñan durante el desarrollo embrionario esos distintos factores solubles, permitirán determinar la función de estas diferentes señales e interacciones en ingeniería tisular, para sintetizar análogos de tejidos funcionales que se puedan transplantar en animales y seres humanos. La modulación de la inducción tisular por este tipo de señales en un futuro contribuirá a la regeneración de tejidos orales. Por estás razones, es fundamental un completo entendimiento de los mecanismos moleculares durante el desarrollo de órganos como sucede durante la Odontogénesis (74).
Actualmente, la investigación en células madres se consideran como una de las líneas de investigación más atractiva para modular la reparación y regeneración de tejidos u órganos como el tejido dental y periodontal (75). La obtención de ASC en diferentes tejidos como médula ósea, complejo pulpo dentinal, cordón umbilical, entre otros; del propio paciente, serán necesarios para permitir una completa regeneración funcional de los tejidos de la cavidad oral, eliminando los problemas de rechazo inmunológico, debido que el transplante es autólogo (76,77). Los avances en técnicas de terapia génica, tecnología del ADN recombinante, terapia celular y observación de expresión génica de las células madres durante su proliferación in vivo y ex vivo, ofrece una alternativa terapéutica funcional para mejorar el estado del paciente.
Esta claro, que las ASCs juegan un papel crucial en el desarrollo, remodelado, regeneración y reparación del complejo craneofacial a lo largo de la vida. El reciente reconocimiento del papel de las células madres en la regeneración tisular, provee una fuerte base de conocimiento que genera un gran impacto en el tratamiento clínico de diversas patologías. A través de la investigación para la utilización clínica de células madres, se podrán generar diferentes protocolos estandarizados para el tratamiento de patologías bucales.
REFERENCIAS
1. Smith A.J. Tooth tissue Engineering and regeneration a translational vision. (Editorial). J Den Res 2004; 83 (7): 517. [ Links ]
2. Salehrabi R, Rotstein I. Endodontic treatment outcomes in a large patient population in the USA: an epidemiological study. J Endod 2004;30:846 -50. [ Links ]
3. Giannoudis P.V., Pountos I. Tissue regeneration: The past, the present and the future. Injury Int J Care Injured. 2005; 36S: 2S-5S. [ Links ]
4. Guan G, Shi S, Kramer P. Role of Adult Stem Cells in Craniofacial Growth and Repair. Semin Orthod. 2005;11:227-233. [ Links ]
5. Munévar JC. in vitro obtention and characterization of Stem Cells from the human umbilical cord as an alternative of embryonic Stem Cells for regenerative Medicine. Rev Lat Bioetica 2005; 9: 40-71. [ Links ]
6. Munévar JC., Becerra A., Hernández A. Biología de las Células Stem. NOVA. 2005; (3)3: 92-105. [ Links ]
7. La cadena JR. Stem Cells and Bioethics. Rev Latin Bioética. 2005; 9. [ Links ]
8. Fuchs E, Segre J. Stem cell: a new lease on life. Cell 2000; (100): 143-155. [ Links ]
9. Verfaillie, C. Adult stem cells: assessing the case for pluripotency. TRENDS in Cell Biology. 2002; 12( 11): 502-508. [ Links ]
10. Jackson KA, Mi T, Goodell MA. Hematopoietic potencial stem cells isolated from murine skeletal muscle. Proc Natl Acad Sci. 1999; 96(25): 14482-6. [ Links ]
11. Ma L, Feng X.Y, Cui B.L, Law F, Jiang X.W, Ying LY. Human Umbilical Cord Warthon´s Jelly derived Mesenchymal Stem Cells differenciation into nerve like cells. Chin Med J (Engl). 2005; 118 (23): 1987-93. [ Links ]
12. Chaparro O. Stem Cells: dream and reality. Rev Lat Bioética 2005; 9: 22-39. [ Links ]
13. Neubauer M., Hacker M, Bauer-Kressel p, Weiser B, Fischbech C, Schulz MB, Goepferich A. Adipose tissue engineering based on mesenchymal stem cells and basic fibroblast growth factor in vitro. Tissue Eng. 2005; 11 (11-12): 1840-51. [ Links ]
14. Zhou DH, Huang SL, Wu YF, Wei J, Chen GY, Li Y, Bao R.The expansion and biological characteristics of human mesenchymal stem cells. [ Links ]
15. Gimeno MJ, Maneiro E, Rendal E, Ramallal M, Sanjurjo L, Blanco FJ. Cell therapy: a therapeutic alternative to treat focal cartilage lesions. Transplant Proc. 2005 Nov; 37(9):4080-3. [ Links ]
16. Armitage JO. Medical progress: bone marrow transplantation. N Engl J Med 1994; 330: 827-838. [ Links ]
17. Grant MB, May WS, Caballero S, Brown GA, Guthrie SM, Mames RN, et al. Adult hematopoietic stem cells provide functional hemangioblast activity during retinal neovascularization. Nat Med 2002; 8: 607-612. [ Links ]
18. Orlic D, Kajstura J, Chimenti S, Jakoniuk I, Anderson SM, Li B, et al. Bone marrow cells regenerate infarcted myocardium. Nature 2001; 410: 701-705. [ Links ]
19. Lagasse E, Connors H, Al-Dhalimy M, Reitsma M, Dohse M, Osborne L, et al. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat Med 2000; 6: 1229-1234. [ Links ]
20. Martin-Rendon E, Watt SM. Stem cell plasticity. Br J Haematol 2003; 122: 877-891. [ Links ]
21. Tsmark N, Tanoka K, Arikawa- Hirasawa L, y col. Role of CDMP-1 in skeletal morphogenesis promotion of mesenchymal cell recruitment and condrocyte differenciation. J. Cell. Biol. 1999; 144: 151-173. [ Links ]
22. Hinton RJ, Carlson DS. Regulation of growth in mandibular condylar cartilage. Semin Orthod (In press). [ Links ]
23. Tsumaki N, Tanaka K, Arikawa-Hirasawa E, et al. Role of CDMP-1 in skeletal morphogenesis: promotion of mesenchymal cell recruitment and chondrocyte differentiation. J Cell Biol. 1999; 144:161-173. [ Links ]
24. Guoqiang Guan, Songtao Shi, and Phillip-Kramer R. Role of Adult Stem Cells in Craniofacial Growth and Repair. Sem in Ortho. 2005; 07.007. 227-233. [ Links ]
25. Opperman LA. Cranial sutures as intramembranous bone growth sites. Dev Dyn 2000; 219:472-485. [ Links ]
26. Johnson D, Iseki S, Wilkie AO. Expression patterns of Twist and Fgfr1-2 and -3 in the developing mouse coronal suture suggest a key role for twist in suture initiation and biogenesis. Mech Dev 2000; 91:341-345. [ Links ]
27. Carlson DS. Growth of the temporomandibular joint, in Zarb GA, Carlsson GE, Sessle B, Mohl ND (eds): Temporomandibular Joint.2nd ed.Copenhagen, Munksgaard, 1994; 128-158. [ Links ]
28. Fuentes MA, Opperman LA, Bellinger LL, et al. Regulation of cell proliferation in rat mandibular condylar cartilage in explant culture by insulin-like growth factor-1 and fibroblast growth factor-2. Arch Oral Biol 2002; 47:643-654. [ Links ]
29. Wise GE, Frazier-Bowers S, D`Souza RN. Cellular molecular and genetic determinants of tooth eruption. Crit Rev Oral Biol Med. 2000; 13: 323-334. [ Links ]
30. Thesleff I. Epithelial-mesenchymal signalling regulating tooth morphogenesis. J Cell Sci 2003;116:1647-8. [ Links ]
31. Shiotani A, Shibasaki Y, Sasaki T. Localization of receptor activator of NFkappaB ligand, RANKL, in periodontal tissues during experimental movement of rat molars. J Electron Microsc. 2001;50(4):365-9. [ Links ]
32. Heino TJ, Hentunen TA, Vaananen HK. Conditioned medium from osteocytes stimulates the proliferation of bone marrow mesenchymal stem cells and their differentiation into osteoblasts. Exp Cell Res 2004; 294: 458-468. [ Links ]
33. Carlson DS. Growth modification: from molecules to mandibles.Growth Modification: What Works What Doesn't and Why. Craniofac Growth Ser 1999; 35:17-65. [ Links ]
34. Sperber GH. Craniofacial Embryogenesis: Normal Developmental Mechanisms in Understanding Craniofacial Anormalies: Etiopathogenesis of Craniosynostoses and Facial Clefting.New York, Wiley-Liss, 2002. [ Links ]
35. Massague J, Blain SW, Lo RS. TGFbeta signaling in growth control cancer and heritable disorders. Cell 2000; 103:295-309. [ Links ]
36. Warner DR, Pisano MM, Roberts EA, et al. Identification of three novel Smad Binding proteins involved in cell polarity. FEBS Lett 2003; 539: 167-173. [ Links ]
37. Haase HR, Ivanovski S, Waters MJ, et al. Growth hormone regulates osteogenic marker mRNA expression in human periodontal fibroblast and alveolar bone-derived cells. J Periodontal Res 2003; 38:366-374. [ Links ]
38. Ohazama A, Sharpe PT. TNF signalling in tooth development. Curr Opin Genet Devel 2004;14:513-9. [ Links ]
39. Thesleff I. Epithelial-mesenchymal signalling regulating tooth morphogenesis. J Cell Sci 2003;116:1647-8. [ Links ]
40. Dominici M, Pritchard C, Garlits JE, et al. Hematopoietic cells and osteoblasts are derived from a common marrow progenitor after bone marrow transplantation. Proc Natl Acad Sci USA. 2004;101:11761-11766. [ Links ]
41. Jiang Y, Jahagirdar BN, Reinhardt RL, et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 2002; 418:41-49. [ Links ]
42. Bhattacharyya A, Svendsen CN. Human neural stem cells: a new tool for studying cortical development in Down's syndrome. Genes Brain Behav 2003; 2:179-186. [ Links ]
43. Dominici M, Pritchard C, Garlits JE, et al. Hematopoietic cells and osteoblasts are derived from a common marrow progenitor after bone marrow transplantation. Proc Natl Acad Sci SA 2004; 101:11761-11766. [ Links ]
44. Labat ML. Stem cells and the promise of eternal youth: embryonic versus adult stem cells. Biomed Pharmacother 2001; 55:179-185. [ Links ]
45. Markakis EA, Palmer TD, Randolph-Moore L, et al. Novel neuronal phenotypes from neural progenitor cells. J Neurosci 2004; 24:2886-2897. [ Links ]
46. Balemans W, Van Hul W. Extracellular regulation of BMP signaling in vertebrates: a cocktail of modulators. Dev Biol 2002; 250:231-50. [ Links ]
47. Miura M, Gronthos S, Zhao M, et al. SHED: Stem cells from human exfoliated deciduous teeth. Proc Natl Acad Sci USA 2003; 100:5807-5812. [ Links ]
48. McCulloch CA, Nemeth E, Lowenberg B, et al. Paravascular cells in endosteal spaces of alveolar bone contribute to periodontal ligament cell populations. Anat Rec 1987; 219:233-242. [ Links ]
49. Almeida-Porada G, Porada C, Zanjani ED. Adult stem cell plasticity and methods of detection. Rev Clin Exp Hematol 2001; 5:26-41. [ Links ]
50. Sloan AJ, Rutherford RB, Smith AJ. Stimulation of the rat dentine-pulp complex by BMP7 in vitro. Arch Oral Biol 2000;45:173-7. [ Links ]
51. Sloan AJ, Smith AJ. Stimulation of the dentine-pulp complex of rat incisor teeth by transforming growth factor-beta isoforms 1-3 in vitro. Arch Oral Biol 1999;44:149 -56. [ Links ]
52. Nakashima M. Induction of dentin formation on canine amputated pulp by recombinant human bone morphogenetic protein (BMP)-2 and -4. J Dent Res 1994;73:1515-22. [ Links ]
53. Decup F, Six N, Palmier B, et al. Bone sialoprotein-induced reparative dentinogenesis in the pulp of rat's molar. Clin Oral Investig 2000;4:110 -9. [ Links ]
54. Nakashima M. An ultrastructural study of the differentiation of mesenchymal cells in implants of allogeneic dentine matrix on the amputated dental pulp of the dog. Arch Oral Biol 1990;35:277- 81. [ Links ]
55. Lovschall H, Fejeskov O, Flyvbjerg A. Pulp-camping with recombinant human insuline- like growth favtor I (rhIGF-I) in rats molars. Adv Den Res 2001; 15:108-112. [ Links ]
56. Gronthos S, Mankani M, Brahim J, et al. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. PNAS 2000; 97: 13625. [ Links ]
57. Pierdomenico Laura, Bonsi Laura, Calvitti Mario, Rondelli Damiano, Arpinati Mario, y col. Multipotent Mesenchymal Stem Cells with Immunosuppressive Activity Can Be Easily Isolated from Dental Pulp. Inmunobiology and genomics. 2005; 80(6): 836-84. [ Links ]
58. Gronthos S., Brahim J., Li W., Fisher L.W., Cherman N., Boyde A. , DenBesten P., Gehron Robey P and Shi S. Stem Cell Properties of Human Dental Pulp Stem Cells. J Den Resch. 2002; 81(8): 531-535. [ Links ]
59. Iohara K., Nakashima1 M., Ito M., Ishikawa1 M., Nakasima A., and Akamine1 A. Dentin Regeneration by Dental Pulp Stem Cell Therapy withRecombinant Human Bone Morphogenetic Protein 2. J. Den Res. 2004; 83(8):590-595. [ Links ]
60. Byoung-Moo Seo, Masako Miura, Stan Gronthos, Peter-Mark Bartold, Batouli Sara, Brahim Jaime, Young Marian, Gehron Robey Pamela, Cn-Yu Wang, Songtao Shi. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004; 364: 149-55. [ Links ]
61. Munévar JC, Forero J, Lafaurie G, Bautista G. y colaboradores. Determinación de los Marcadores CD117 y FGFR3 en Muestras de Pulpa Dental sana Humana por Inmunohistoquímica. Instituto U.I.B.O. Universidad el Bosque. 2006. [ Links ]
62. Gronthos S, Mankani M, Brahim J, et al. Postnatal human dental pulp stemc ells (DPSCs) in vitro and in vivo. Proc Natl Acd Sci USA. 2000; 97: 13625-13630. [ Links ]
63. Chai Y, Slavkin HC. Prospect for tooth regeneration in the 21st century: a perspective. Micros Res Tech. 2003; 60: 469-479. [ Links ]
64. Duailibi MT, Duailibi SE, Young CS, et al. Bioingeneered teeth from cultured rat tooth bud cells. J Dent res. 2004; 83: 523-528. [ Links ]
65. Bianco P., Robey PG. Stem cells in tissue engineering. Nature. 2001; 414 (6859): 118-121. [ Links ]
66. Seo BM, Miura M, Gronthos S, et al. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004; 364:149- 155. [ Links ]
67. Duailibi MT, Duailibi SE, Young CS, et al. Bioengineered teeth from cultured rat tooth bud cells. J Dent Res 2004; 83:523-528. [ Links ]
68. Dominici M, Pritchard C, Garlits JE, et al. Hematopoietic cells and osteoblasts are derived from a common marrow progenitor after bone marrow transplantation. Proc Natl Acad Sci USA 2004; 101:11761-11766. [ Links ]
69. Seo B.M., Miura m., Gronthos S., et al. Investigation of multipotent post natal stem cells from human periodontal ligament. Lancet. 2004; 364: 149-155. [ Links ]
70. Jervall and Thesleff, 2000, http://bite-it.helsinki.fi/. [ Links ]
71. Ohazama A, Modino S.A., Miletich I., et al. Stem cell- based tissue engineering of murine teeth. J Dent. Res 2004; 83: 518-522. [ Links ]
72. Harada H, Ohsina H. new perspectivas on tooth developenment and the dental stem cell niche. Arch Histol Cyto. 2004; 67(1): 1-11. [ Links ]
73. Garin NC. Autologus bone marrow tranplation in acurate leucemia. J Natl cáncer Inst. 1986; 76:1281-1287. [ Links ]
74. Pereira RF, O´ltec MD, lapter AV, et al. marrow stromal cells as a source of progenitor cells for nonhematopoietic tisúes in transgenic mice whit a phenotype of osteogenesis imperfecta. proa. Natl Acad Sci. USA. 1998; 95: 1142-1147. [ Links ]
75. Grant MB, May WS, Caballero S, Brown GA, Guthrie SM, Mames RN, et al. Adult hematopoietic stem cells provide functional hemangioblast activity during retinal neovascularization. Nat Med 2002; 8: 607-612. [ Links ]
76. Arinzeh T.L., Peter S.J., Archambault M.P., van-den B.C., Gordon S., Kraus K., Smith A., Kadiyala S. Allogeneic mesenchymal stem cells regenerate bone in a critical-sized canine segmental defe ct. J Bone Joint Surg Am 2003; 85-A: 1927-1935. [ Links ]
77. Grinnemo K.H., Mansson A., Dellgren G., Klingberg D., Wardell E., Drvota V., Tammik C., Holgersson J., Ringden O., Sylven C., Le Blanc K. Xenoreactivity and engraftment of human mesenchymal stem cells transplanted into infarcted rat myocardium. J Thorac Cardiovasc Surg 2004; 127:1293-1300. [ Links ]
