










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgronomía Tropical
versión impresa ISSN 0002-192X
Agronomía Trop. v.56 n.2 Maracay jun. 2006
TRABAJO ESPECIAL
ORIGEN, EVOLUCIÓN Y DIVERSIDAD DEL ARROZ
Marco A. Acevedo*, Willian A. Castrillo* y Uira C. Belmonte**
* Investigador. INIA. Centro Investigaciones Agrícolas Estado Guárico. Km 27, carretera Calabozo San Fernando de Apure, sector Bancos de San Pedro, Apdo. 14. Calabozo, estado Guárico, Venezuela. Email: macevedo@inia.gob.ve.
** Ejercicio Libre. Departamento de Genética de la ESALQ/Univesidad de São Paulo, Brasil.
RESUMEN
Este trabajo tiene por objetivo presentar una revisión bibliográfica sobre el origen, evolución y diversidad del género Oryza, en las especies cultivadas. La especie O. sativa es la de mayor importancia económica, ya se cultiva en todo el mundo en climas tropicales y templados (latitud de 35° sur en la Argentina a 50° norte en la China), O. glaberrima, se cultiva solamente en el oeste de África. Se deduce que existen dos patrones evolutivos de origen y domesticación del arroz cultivado, uno en Asia para la especie O. sativa y otro en África para O. glaberrima. La principal consecuencia de la domesticación en O. sativa es la reducción del desgrane, que favorece la cosecha manual y mecanizada. Trabajos de diversidad genética demuestran que la O. sativa presenta mayor variabilidad genética encontrándose hasta 3 sub-especies (Indica, Japónicas y Javánicas), basadas en su ecología y morfología, la tendencia no fue observada en O. glaberrima. Los estudios sobre genoma realizado en la especie O. sativa Subsp Japónica muestran que esta formado por 430 millones de pares de bases y aproximadamente 50% está compuesto de secuencias repetidas. Finalmente, la estrecha base genética encontrada en las variedades de arroz liberadas en el mundo en los últimos años como consecuencia de la uniformidad del núcleo citoplasmática, ha traido como consecuencias un techo en la productividad y vulnerabilidad a factores bióticos. En virtud de ello numerosos investigadores de diversos países han realizado trabajos de mejoramiento genético con ayuda de la técnicas biomoleculares para incrementar la diversidad genética en la especie O. sativa, utilizando para ello germoplasma exótico del género Oryza, principalmente las especies O. glaberrima, O. sativa spontánea, O rufipogon, O. officinalis, pertenecientes a los complejos O. sativa y O. Officinalis.
Palabras Clave: Oryza sativa L.; origen; taxonomia; diversidad; germoplasma exótico.
TRABAJO ESPECIAL
RICE ORIGIN, EVOLUTION AND DIVERSITY
SUMMARY
This review has the objective to present a detailed bibliographical revision about origin, evolution and diversity of Oryza genus, with emphasis in the cultivated species. The O. sativa species is the one of more economic importance and is widely cultivated throughout the world in tropical and temperate climates (latitude 35° south in Argentina to 50° north in China), while O. glaberrima is only cultivated in the western Africa. These studies suggest that there are two evolutionary patterns of origin and domestication of the cultivated rice, being O. sativa from Asia and O. glaberrima from Africa. The most important consequence of domestication in O. sativa is the reduction of dehiscence, which favors the manual and automatic harvest. Studies show that O. sativa presents higher genetic variability geing up to three sub-species (Indica, Japonicas and Javanicas), with different ecology and morphology; however this tendency was not observed in O. glaberrima. Genomic studies show that O. sativa subsp Japonica has 430 megabases and approximately 50% are repeated sequences. Finally, the narrow genetic base found in liberated varieties of rice in the world in the last years, has resulted in a selection progress plateau in yield and vulnerability to biological factors as a consequence of the lack of variability in both the nuclear and cytoplasmic genetic background. Numerous researchers from many countries have carried out studies of breeding programs using molecular technique to increase genetic diversity in O. sativa, using exotic germoplasm of O. genus, mainly the species O. glaberrima, O. sativa spontanea, O. rufipogon, O. officinalis, belonging to the complex O. sativa and O. officinalis.
Key Words: Oryza sativa L.; origin; taxonomy; diversity; exotic germplasm.
RECIBIDO: julio 31, 2006.
INTRODUCCIÓN
El cultivo del arroz, Oryza sativa L., comenzó hace casi 10.000 años, en muchas regiones húmedas de Asia tropical y subtropical. Este cultivo es el alimento básico para más de la mitad de la población mundial. A nivel mundial, ocupa el segundo lugar después del trigo con respecto a superficie cosechada. El arroz proporciona más calorías por hectárea que cualquiera de los otros cereales cultivados.
El arroz pertenece a la División: Angiospermae, Clase: Monocotyledoneae, Orden: Glumiflorae, Tribu: Oryzeae, Familia: Poaceae (gramineae), siendo las especies cultivadas: Oryza sativa L. y Oryza glaberrima Steud, ambas son especies de reproduccción autogama, diploides con 2n=24 cromosomas. El género Oryza tiene más de 24 especies silvestres que crecen en regiones inundadas, semi-sombreadas y bosques en el sureste Asiático, Austria, África, Sur y Centro América.
Dos hipótesis explican el origen de las especies cultivadas, estos estudios suponen que la especie ancestral de O. sativa puede ser O. nivara (Asia perennis) del sur y el sureste de Asia. En cuanto que de O. glaberrima, puede ser África perennis vía O. breviligulata, del África tropical.
El objetivo de este trabajo es realizar una revisión bibliográfica detallada sobre el origen, evolución, domesticación, sistema de reproducción y estudio de la diversidad genética del género Oryza, con énfasis en las especies cultivadas, en razón que los estudios adelantados hasta el momento están publicados en idiomas distintos al español. Además la idea es colocar a disposición de los interesados una revisión bibliográfica actualizada sobre el tema.
Taxonomía y Diversidad Genética
Las especies del género Oryza, por su gran importancia agronómica, han recibido una gran atención por parte de los investigadores de todo el mundo sobre estudios taxonómicos, filogenéticos, entre los cuales se pueden señalar: Linnaeus (1753), Baillion (1894), Prodoehl (1922), Roschevicz (1931), Chevalier (1932), Chatterjee (1948), Sampath y Rao (1951), Richharia (1960), Tateoka (1963), Ghose et al. (1956), Sharma y Shastry (1972), citados por Watanabe (1997). Además Chang (1976), Vaughan (1994), Morishima et al. (1984), Lu et al. (1998), Lu (2004), Oliveira (2004) y más recientemente The Office of the Gene Technology Regulator de Australia (2005).
El número y nombre de las especies correspondientes a este género difiere entre dichos investigadores. Una de las mayores contribuciones a la clasificación del arroz fue dada por Roschevicz en 1931, al proponer la base para los siguientes estudios taxonómicos de género Oryza. En este estudio incluyó 19 especies de este género y los dividió en cuatro secciones (Sativa, Granulata, Coartata y Rhynchoryza). Chevalier en 1932 clasificó el género Oryza dentro de cuatros secciones (Euoryza, Padia, Scherophyllum y Rhynchoryza) y 23 especies. Ghose, Ghatge y Subrahmanyan en 1956, dividen el género Oryza en tres secciones (Sativa, Officinalis y Gramulata) y 21 especies, todos citados por Watanabe (1997).
Por su parte, Morishima y Oka (1960), calcularon el coeficiente de correlación de 42 caracteres y 16 especies del género Oryza, concluyendo que la sección Sativa propuesta por Roschericz en 1931, puede ser subdividida en tres subsecciones (Sativa, Officinalis y Australiensis). Más recientemente, Tateoka (1964) dividió el género en cinco secciones (Latifólia, Sativa, Glaberrima, Rdleyi y Meyesiana). Sharma y Shastry (1971) citado por Watanabe (1997), dividen el género Oryza en tres secciones (Padia, Angustifolia y Oryza), estableciendo dos y hasta tres series dentro de cada sección y 28 especies. Sharma (1986) estudió estas secciones del género Oryza y concluye que la sección Padia está representada por especies del sureste de Asia, siendo plantas pequeñas, perennes, creciendo en ambientes sombreados y adaptadas a suelos bien drenados, semillas sin aristas de tamaño medio, ejemplo (O. schlechteri, O. meyeriana, O. ridleyi).
La sección Angustifolia está representada por especies del grupo de África, adaptadas a ambientes abiertos, plantas perennes y anuales que crecen en lugares pantanosos, ejemplo (O. perrieri, O. brachyantha, O. angustifólia). Entretanto, la sección Oryza está distribuida en los trópicos, plantas altas, adaptadas a condiciones de alta humedad y en ambientes abiertos, ejemplo (O. latifólia, O. australiensis y O. sativa).
Los mismos autores estudiaron los principales caracteres del género Oryza, los cuales están resumidas en el Cuadro 1. Este estudio indica que la sección Padia parece ser más primitiva, mientras Oryza ocupa la posición avanzada. Las especies en secciones o complejos diferentes presentan características morfológicas diferentes, y las especies dentro de una misma sección tienen una delimitación ambigua.

Lu (2004) propone un sistema taxonómico para el género Oryza basado en el sugerido por Sharma y Shastry en 1965, con ciertas modificaciones. En esta propuesta 24 especies fueron reorganizadas y colocadas en 3 secciones: la sección Padia (3 series y 6 especies), la sección Oryza (3 series y 17 especies) y la sección Brachyantha (en 1 serie y 1 especie).
Oliveira (2004) realizó un estudio biogeográfico y ecológico de los arroces silvestres nativos del Brasil, entre ellos: O. glamaepatula (AA) diploide, O. grandiglumis, O. alta y O. latifolia (estas últimas alopoliploides). Concluyó que O. grandiglumis está restringida al Amazona Occidental y extremo Norte de Pantanal brasileño. O. alta está presente en las Planicies del Litoral Atlántico, incluyendo el Valle de Ribeira Maranhense, estado de Paraná, Paraguay y Pantanal.
La especie alotetraploide O. latifolia está centrada en las Planicies Costera del Pacífico y en los Valles de los Ríos Andinos, habitando regiones húmedas e inundadas, también en paisajes típicamente áridos o mixtos, como la Península de la Guajira y Santa Elena de Uairén en Venezuela, así como en el Ecuador. La especie diploide O. glamaepatula, presenta 3 morfotipos o razas geográficas: una en el Caribe, otra en la Amazona y la tercera en el Pantanal brasileño. Esos grupos difieren entre sí no sólo en la morfología, sino también en su estructura genética a nivel de marcadores moleculares (Akimoto, 1999), citado por Oliveira (2004).
Los estudios de filogenia y sistemática, realizados por Oliveira (2004) sobre el género Oryza, permitió sugerir un nuevo arreglo de la sistemática de este género, como sigue, tres secciones: Granulata (2 especies), Ridleyi (2 subsecciones y 5 especies) y Euoryza (2 subsecciones Australiensis e Officinalis), esta última con cuatro series y 19 especies.
Los estudios antes señalados muestran que aún existen muchas controversias sobre la taxonomía, filogenia y sistemática del arroz. Además, no existe un único sistema de nomenclatura, sin embargo, las especies cultivadas (O. sativa y O. glaberrima) y siete silvestres (O. australiensis, O. eichingeri, O. latifolia, O. minuta, O. schechteri, O. ridleyi y O. brachyantha) no le fueron alterados sus nombres originales.
Los nombres de las especies del género Oryza presentado en el Cuadro 2, son los más usados en los estudios actuales de taxonomía, debido a su consistencia en la literatura, los complejos más estudiados son O. sativa y O. officinalis.

Origen y Evolución
Hasta el presente 24 especies son generalmente incluidas dentro del género Oryza. De éstas, las especies asiática perennes conocidas como O. ruffipogon y O. nivara (anual) son los progenitores de la especie cultivada O. sativa L., mientras que las especies africanas perennes (O. barthii) y anual (O. breviligulata) son los progenitores directos de la especie cultivada O. glaberrima Steud., cultivada básicamente en el oeste del África.
Existen dos hipótesis que sustentan el origen de las especies cultivadas de arroz:
(a) Hipótesis de Chang (1976), propone que las 24 especies del género Oryza son originarias del Continente Gondwanaland (continente este formado por uno sólo antes de la separación en los cinco conocidos actualmente) y que la especie ancestral de O. sativa puede ser O. nivara (Asia perennis), del sur y el sureste de Asia. En cuanto que de O. glaberrima, puede ser África perennis vía O. breviligulata, del África tropical. Es válido detallar que Asia, América, África y Oceanía perennis fueron creadas desde un ancestol común en el Continente Gondwanaland y que envolvió especies diferentes con la división de los cinco continentes por diferentes rutas de evolución. En la Figura 1 se representa esta hipótesis. Trabajos más recientes donde fue analizada la variabilidad fenotípica y molecular, indican que esta teoría sería la más adecuada, Morishima, Sano y Oka (1992), citados por Guimarães (1999).

(b) Morishima (1976), citado por Watanabe (1997), propone que Asia perennis es el ancestral de O. sativa, como lo muestra la Figura 2. Esta teoría está basada en: (1) la especie perennes presentaban mayor variabilidad genética que las especies anuales; (2) sativa presentaba características de perennes y (3) existen muchos tipos intermedios entre las especies silvestres y cultivadas que fueron encontrados en la India, presentando características de perennes. La especie spontanea, tipo anual, fue probablemente formada a partir de Asia perennis a través de otra ruta de evolución.

Entonces, se deduce de estos estudios que existen dos patrones evolutivos de origen y domesticación del arroz cultivado, uno en Asia para la especie O. sativa y otro en África para O. glaberrima.
Los recientes trabajos están principalmente enfocados en la determinación de la relación entre los diferentes genomas usando alopoliploides obtenidos artificialmente. Se sabe que el genoma A está presente en las especies cultivadas y en sus parentales más próximos. El genoma C está presente en las especies que tienen principalmente polinización cruzada y se conoce que los genomas B, C y D presentan alguna afinidad entre ellos. Morishima (1984) muestra que las especies cultivadas O. sativa y O. glaberrima, así como sus perentales silvestres mas próximos, perennes (incluyendo O. rufipogon y O. barthii) y anual (O. nivara y O. breviligulata) formaron un grupo que posee el genoma A, en común. La especie O. officinalis y sus parentales con genomas B, C y D formaron otro grupo. Las especies que llevan el genoma A, son subdivididas en A, Ab, Acu y Ag, nomenclatura establecida por el IRRI para designar esterilidad parcial y menor apareamiento cromosómico entre híbridos de diferentes sub-genomas.
Chang (2003), concluye que la especie O. sativa tiene un número básico de cromosomas de (X=12), mientras que el genoma diploide (2n=24), este es considerado relativamente pequeño cuando comparado con cualquier otro genoma estudiado con 430 millones de pares de bases, aproximadamente 50% del genoma esta compuesto de secuencias repetitivas.
De las especies cultivadas, O. sativa es la de mayor importancia económica, ya que es cultivada ampliamente en todo el mundo, incluyendo Asia, América del Norte y Sur, Unión Europea, Oceanía y Centro-este de África. La otra especie cultivada, O. glaberrima, se cultiva solamente en el oeste de los países africanos. Además de las diferencias filogenéticas entre las especies cultivadas del género Oryza, existen diferencias morfológicas entre ellas. Las diferentes formas de O. glaberrima pueden ser diferenciadas de las asiáticas, entre otras, por presentar lígula corta, 0,40 cm y 2,09 cm, respectivamente. Además de tener aspecto rudimentario y fuerte, O. glaberrima también presenta menor número de ramificaciones secundarias en la panícula y las plantas mueren después de la maduración (senescencia precoz). Los otros caracteres no son utilizados para diferenciar ambos genotipos.
La especie O. sativa presenta mayor diversidad genética encontrándose hasta tres sub-especies, las cuales son clasificadas basada en su ecología y morfología en: Indica, Japónicas y Javánicas; mientras que en O. glaberrima, tal tendencia no fue encontrada. Esto puede sugerir que diferencias en el sistema genético de las especies silvestres ancestrales pueden haber llevado a diferentes tipos de evolución de las formas cultivadas. La sub-especie Indica está distribuida en los trópicos y subtrópicos, la Javánica se cultiva en Indonesia, siendo también conocida como Japónica tropical, mientras que la Japónica, se encuentra distribuida en zonas no tropicales (templadas), sin embargo, existe sobre posición de caracteres entre esos tipos.
Glaszmann en 1987 citado por Guimarães (1999), utilizó marcadores isoenzimáticos y agrupó las variedades de arroz en 6 conjuntos, según la distancia genética. Esos grupos fueron identificados con la numeración del I al VI; el grupo I reúne las variedades consideradas Indicas por la clasificación de Oka (1958) y el último agrupa a las variedades Japónicas. De modo general, se puede decir que las variedades adaptadas a sistemas de cultivo de secano pertenecen al grupo VI y los del sistema irrigado al grupo I.
Katayama (1997) evaluó diferentes especies silvestres y cultivadas, muestreadas en varios países (India, Sri Lanka, Indonesia, China, Corea y Tailandia). Cinco características fueron empleadas para la clasificación, las cuales se muestran en el Cuadro 3.
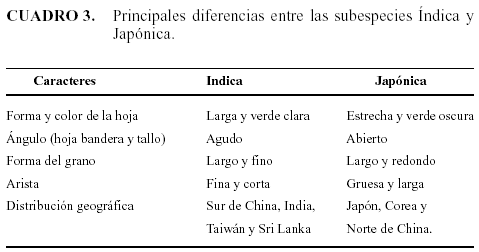
Existen otros caracteres empleados para diferenciar estos dos tipos, tales como la sensibilidad a la temperatura y la tolerancia a la sequía. McDonald (1994) encontró que cultivares Japónica crecen predominantemente en regiones templadas y las temperaturas bajas de 15° - 20 °C no afectan la germinación ni el crecimiento vegetativo, lo contrario ocurre con los cultivares Índica. Los arroces silvestres que se encuentran en la China, Hawai, EEUU y Brasil, contienen características de las subespecies Índicas y Japónicas.
Domesticación, Dispersión y Modo de Reproducción
El proceso de domesticación de la especie O. sativa no está bien claro. De Candoll en 1882 propone que el arroz es originario de India o de Indochina. Vavilov en 1926 y 1951 sugiere la teoría de los Centros Dominantes de Genes, considerando que la domesticación del arroz ocurrió en la India, esta última hipótesis recibió apoyo de un gran número de investigadores de la época. Además esta teoría está basada sobre el reconocimiento de la gran variabilidad genética, así como la estrecha relación ecológica entre las especies silvestres y cultivadas observadas en la India, todos citados por Morishima (1984).
Ho (1969), investigador chino, propone la posibilidad que la domesticación pudo ocurrir en la China, las evidencias arqueológicas e históricas encontradas apoyan esta propuesta, indicando que el cultivo de arroz ocurrió en China 1000 años antes que en la India.
Harlan (1975), consideró para el cultivo del arroz la existencia de varias localidades de domesticación como más razonable. Entonces si esta hipótesis es verdadera, esto permite inferir que la diferenciación entre Índica y Japónica también tiene un origen similar, siendo el ambiente el principal factor diferenciador.
Los estudios recientes sobre el origen y la domesticación del arroz, señalan a la región del Himalaya y suroeste de la China, como Centros de Diversificación y Domesticación de la especie O. sativa. Esto esta apoyado por la presencia y conservación de la variación genética existente en la zona, debido a la diseminación de cruzamientos y aislamiento de dichas condiciones ambientales.
Oka (1988) y OECD (1999), señalan que como producto de la domesticación O. sativa ha desarrollado muchos tipos o variedades que permiten su adaptación a amplias condiciones ambientales, tales como: (a) climas tropicales o templados (desde latitud de 35° sur en la Argentina a 50° norte en la China); (b) diferentes tipos de suelo (110 países, desde el nivel del mar hasta 3.000 m de altitud) y (c) baja o alta dependencia de lámina de agua durante el ciclo de cultivo.
Morishima (1984), resaltó como principal cambio producto de la domesticación del arroz, la disminución del desgrane de la panícula, facilitando la cosecha, este carácter esta dominado por un simple gen recesivo. Otros cambios provenientes del proceso de domesticación se muestran en el Cuadro 4.

Las rutas de dispersión del arroz en el mundo son presentadas en la Figura 3, iniciándose del sureste asiático (India) para la China 3000 años antes de Cristo (aC). De allí fue para Corea y posteriormente para el Japón, en el siglo I aC. También de la India fue llevado para las islas del Océano Indico, principalmente Indonesia y Sri Lanka en la misma fecha. Generalmente los cultivos domesticados en el Oriente del Asia fueron difundidos al Occidente en épocas relativamente recientes. Los comerciantes árabes fueron los primeros en traer el arroz del este de Asia para el Medio Oriente, cerca del siglo IX aC. Simultáneamente en esta época fue llevado para Egipto y otros países del África, donde sólo se cultivaba O. glaberrima. La introducción del arroz al Occidente ocurrió aproximadamente 320 aC, mientras que en América ocurrió en época pos-colombina, siendo traído por los colonizadores españoles, portugueses y holandeses.

Oka y Morishima (1967) encontraron importantes diferencias en el arroz cuando compararon sus formas silvestres y cultivadas, principalmente en el sistema de polinización. Las especies cultivadas son predominantemente autógamas, mientras sus progenitores silvestres son parcialmente alógamos. Las flores de las plantas silvestres tienen varios mecanismos que favorecen la alogamia, entre ellos: grandes anteras, estigma largo que lo expone al polen de las plantas más próximas, así como mayor tiempo de emisión y viabilidad del polen después de la apertura de la flor. Al contrario de las plantas cultivadas donde las anteras son cortas y sobrepuestas al estigma y el polen es liberado antes de la apertura de la flor (cleistogamia), lo cual favorece la autogamia.
La tasa de cruzamiento fue estimada en la especie Asia perennis (perenne) entre 30-50% y en la anual de 5-30%, ambas parentales directos de O. sativa. Las especies africana perennis (perenne) parecen ser predominantemente alógamas con parcial incompatibilidad. En la especie O. breviligulata, (anual), tiene polinización principalmente autógama con 5-30% de alogamia, siendo estas dos últimas, parentales de O. glaberrima.
Utilización del germoplasma silvestre en el mejoramiento genético del arroz
Los arroces cultivados han llegado a diferenciarse de los arroces silvestres, como en otros cultivos, por la intervención del hombre al seleccionar las formas deseables, para su alimentación, principalmente. Como se mencionó anteriormente, el hombre ha desarrollado variedades de O. sativa que se adaptan a un amplio rango de condiciones de ambientales.
En el banco de germoplasma del IRRI existe alrededor de 100.000 accesiones, la mayoría pertenecientes a la especie O. sativa, estos materiales están caracterizados basado en la adaptación a diferentes condiciones de humedad, hábito de crecimiento, altura de planta, tamaño y color del tallo y la hoja bandera, así como a características de la panícula y semilla, además de su reacción a plagas y enfermedades. Todas estas características son de valiosa ayuda a los programas de mejoramiento genético del cultivo, Vaughan (1994).
En los programas de mejoramiento genético de especies autógamas, es usual la utilización de la variabilidad genética disponible en las variedades locales o introducidas (poblaciones heterogéneas y homocigotas). Los cruzamientos entre líneas emparentadas (coeficiente de parentesco F alto) y el uso de repetido número de genitores en los programas de mejoramientos, han conducido al aumento del parentesco, lo cual reduce la base genética. Por otro lado la endogamia natural, debida a la autofecundación de las especies autógamas, restringe la recombinación genética, aumentando la uniformidad genética en el núcleo y el citoplasma, limitando también de algún modo la variabilidad genética.
Cuevas Pérez et al. (1992), estudiaron las variedades de arroz liberadas en América Latina y el Caribe durante el período 1971-1989 y concluyeron que ellas poseen en su genealogía 14 cultivares en común, probablemente provenientes de 7 países y poseen cerca de 69% de similitud en su constitución genética. Esto puede traer como consecuencia la vulnerabilidad genética del germoplasma a los estreses bióticos y abióticos, además de la dificultad para superar el techo de productividad, actualmente existente en las variedades comerciales.
Existen varios métodos en mejoramiento genético que permiten ampliar la base genética entre los cuales están: (a) la selección recurrente, ya que permite utilizar mayor cantidad de progenitores, incrementando la recombinación y aumentando la frecuencia de alelos favorables, (b) utilización de germoplasma exótico (silvestre), siendo que este flujo de genes puede ocurrir entre especies o dentro de una especie. Para que la transferencia de genes tenga éxito hay que satisfacer tres principios básicos: (1) sobre posición espacial, (2) sobre posición temporal (dentro de día y año) y (3) debe existir suficiente afinidad biológica, Den Nijs (2004) citado por TOGTR, (2005).
El flujo de genes a través de la hibridación sexual convencional está restringido a las líneas de O. sativa y especies que llevan el genoma A dentro del género. El flujo entre especies menos relacionadas, principalmente fuera del género oryza, está restringido a técnicas de mejoramiento artificial como rescate de embriones e hibridación somática. En la literatura, en general, sólo se presentan estudios de hibridación entre las especies O. sativa, O. officinalis, O. minuta, O. glaberrima y O. brachyantha, esos cruces no ocurren naturalmente, sino utilizando técnicas moleculares que permiten obtener los híbridos fértiles.
Jena (1994), relata que los híbridos obtenidos con técnicas de rescate de embriones entre O. sativa y Porteresia coartata (miembro de la tribu oryzeae), producen plantas macho estériles, las cuales podrían favorecer la transferencia para tolerancia a salinidad en arroces cultivados.
Martínez et al. (2004), relatan que nuevos alelos para aumentar la variabilidad genética del rendimiento de grano en el arroz comercial en América Latina está en el uso de las 20 especies silvestres del género oryza. Utilizando la técnica desarrollada por la Universidad de Cornell en EE.UU, investigadores del CIAT (Centro Internacional de Agricultura Tropical) en Colombia, están explorando el potencial de algunas especies con la ayuda de marcadores moleculares. Ellos realizaron cruces entre O. rufipogon y O. glaberrima con las especies cultivadas (Bg 90-2, Oryzica 3 y Caiapó) donde las variedades comerciales fueron usadas como recurrentes y las silvestres como donadoras en el programa inter específico de mejoramiento por retrocruzamento.
Las líneas obtenidas del cruce entre Bg 90-2 y O. rufipogon tuvieron rendimiento superior entre 5-25% sobre la variedad comercial que produjo 7,2 t ha-1. En cuanto a la calidad de grano ambos progenitores presentan baja calidad de grano, pero líneas con segregación transgresivas fueron obtenidas, con grano fino y traslúcido. Los mismos autores señalan que Rhizoctonia solani y Sarocladium oryzae son dos enfermedades que últimamente han causado importante perdidas al cultivo en América Latina donde todas las variedades comerciales son susceptibles. Resultados obtenidos del cruce entre Oryzica 3 y O. rufipogon produjeron líneas con una buena tolerancia a ambas enfermedades.
En el mismo trabajo encontraron que O. glaberrima presentó tolerancia a una enfermedad causada por el hongo Polymyza graminis transmisor del virus de la necrosis rallada del arroz (RSNV), enfermedad detectada en Colombia en 1991 y recientemente en Panamá y Brasil, donde los cultivares comerciales son todos susceptibles. Resultados análogos son mostrados por Moncada (2001), trabajando con cruces interespecíficos entre O. sativa y O. rufipogon.
La literatura señala que algunas accesiones de O. gaberrima, así como líneas derivadas del cruzamiento con O. sativa (líneas NERICA), muestran tolerancia a severos estrés por sequía, mostrando diversos mecanismo de tolerancia tales como: arreglo osmótico, arreglo de los estomas y sistema radical profundo. Como se sabe esta característica está controlada por muchos genes que actúan en varios mecanismos fisiológicos, bioquímicas, fonológicos y morfológicos.
Ghneim et al. (2005), investigadores del IVIC e INIA en Venezuela y CIAT en Colombia, iniciaron trabajos utilizando O. glaberrima para estudiar tolerancia a sequía, empleando técnicas de marcadores moleculares (QTL).
En resumen a continuación se presentan algunos genes de interés agronómico introducidos a la especie O. sativa L., provenientes de especies silvestres de arroz:
1. Transferencia del citoplasma macho estéril desde la maleza arroz rojo (O. sativa spontánea), Lin y Yuan (1980),
2. Genes de resistencia a enfermedades causadas por hongos desde O. officinalis, Jena y Khush (1990),
3. Genes con resistencia a enfermedades bacterianas desde O. longistaminata, Khush et al. (1990),
4. Genes con tolerancia a la salinidad desde Porteresia coarctata (miembro de la tribu oryzeae), Jena (1994),
5. Genes QTL para productividad de grano desde O. rufipogon, Moncada (2001),
6. Genes con tolerancia a sequía desde O. glaberrima, Martínez (2004),
7. Genes con resistencia y/o tolerancia a plagas y enfermedades desde O. rufipogon, Martínez (2004).
BIBLIOGRAFÍA
1. CHANG, T.T. 2003. Origin, domestication and diversification. Chapter 1.1. In: W. C. Smith, R. H. Dilday eds. Rice, origin, history, tecnology and production. Jonh Wiley and Sons Inc. Hoboken, New Jersey. p. 3-24. [ Links ]
2. CHANG, T.T. 1976. The origin, evolution, cultivation, dissemination and diversification of Asian and African rices. Euphytica 25: 425-441. [ Links ]
3. CUEVAS PÉREZ, F., E. GUIMARAES, L. BERRIOS and D. GONZÁLEZ. 1992. Genetic base of irrigated rice in Latin America and Caribbean, 1971 to 1989. Crop Sci. 32:1 054-1 059. [ Links ]
4. GHNEIM, T., P. ALEJANDRO, I. PÉREZ, G. TORREALBA, C. MARTÍNEZ, LORIEUX M. y J. TOHME. 2005. Plant Breeding News, Edition 160 10. An Electronic Newsletter of Applied Plant Breeding by FAO and Cornell University. http://www.fao.org/WAICENT/FAOINFO/ AGRICULT/AGP/AGPC/doc/services/pbn.html [ Links ]
5. GUIMARÃES, E. 1999. Hibridação em arroz. In: A. Borem eds. Hibridação artificial de plantas. Universidade federal de Viçosa, p. 101-119. [ Links ]
6. HARLAN, J.R. 1975. Crop and man. Amer. Soc. Agron. Madison, Wisconsin, 295 pp. [ Links ]
7. HO, P.T. 1969. The loess and origin of chinese agriculture. Amer. Historical Review 75:1-36. [ Links ]
8. JENA, K.K. 1994. Prodution of intergeneric hybrid between O. sativa L. and Porteresia coarctata T. Current Science 67: 744-746. [ Links ]
9. JENA, K. and G. KHUSH. 1990. Introgression of gene fron Oryza officinalis Well & Watt to cultivate rice, O. sativa L. Theoretical and Applied Genetic 80: 737-745. [ Links ]
10. KATAYAMA, T. 1997. Relationships between chromosome numbers and genomic constitutions in genus Oryza. In: T. Matsuo, Y. Futsuhara, F. Kikuchi, and H. Yamaguchi eds. Science of the rice plant. Food and Agriculture Policy Research Center, Tokyo. [ Links ]
11. KHUSH, G.S., E. BACALANGCO and T. OGAWA. 1990. A new gene for resistance to bacterial blight from O. longistaminata. Rice Genetic newsletters 7:121-122. [ Links ]
12. LIN, S. and L. YUAN. 1980. Hybrid rice breeding in China. In: Innovative approaches to rice breeding. International Rice Research Instituite, Manila, the philippines, pp. 35-51. [ Links ]
13. LU, B.R. 2004. Taxonomy of the genus Oryza (Poaceae): Historical perpective and current status. International Rice Research Notes IRRN. 24:4-9. [ Links ]
14. MARTÍNEZ, C.P., J. BORRERO, A. ALMEIDA, M. DUQUE, F. CORREA-VICTORIA, J. SILVA y J. TOHME. 2004. Utilization of new alleles from wild rice species to improve cultivated rice in Latin America. CIAT Calí. http://www.ciat.cgiar.org. Cali, Colombia. [ Links ]
15. McDONALD, D.J. 1994. Temperate rice technology for the 21st century: an Australian example. Australian Journal of experiemental Agriculture 34:877-888. [ Links ]
16. MONCADA, P., C.P. MARTÍNEZ, J. BORRERO, M. CHATEL, H. Jr. GOUCH, E. GUIMARÃES, J. THOME and S.R. McCOUCH. 2001. Quantitative trait loci for yield and yield componentes in O. sativa por O. ruffipogon BC2F2 population evaluated in an upland enviroment. Theor. Appl. Genetic. 102: 41-52. [ Links ]
17. MORISHIMA H. 1984. Wild plants and domestication, chapter 1. In: S. Tsunoda, N. Takahashi, eds. Biology of rice, Volumen 7. Elsevier, Amsterdam, p. 3-30. [ Links ]
18. MORISHIMA, H. and H. OKA. 1960. The pattern of interspecific variation in the genus Oryza: it´s quantitative representation by stadistcal methods. Evolution 14:153-165. [ Links ]
19. ORGANIZATION FOR ECONOMIC CO-OPERATION and DEVELOPMENT (OECD). 1999. Concensus document on the biology of Oryza sativa rice, OECD Environmental Health and Safety Publications, Paris. [ Links ]
20. OKA, H.I. 1988. Origin of cultivated rice. Elsevier, Amsterdam, 254 pp. [ Links ]
21. OKA, H.I and H. MORISHIMA. 1967. Variation in the breeding systems of a wild rice, Oryza perennis. Evolution 21:249-258. [ Links ]
22. OLIVEIRA, G. 2004. Ecologia e evolução no gênero Oryza (arroz poaceae). In: 40 Anos de Pós-Graduação em Genética e Melhoramento. Anais, 21 encontro sobre temas de genética e melhoramento, ESALQ/ USP, 2004. Vol 21: 26-32. [ Links ]
23. SHARMA, S.D. 1986. Evolutionory trends in genus Oryza. In: Rice genetic. Proceedings of the International Rice Genetic Symposium, IRRI Manila Phillipines. [ Links ]
24. TATEOKA, T. 1964. Taxonomic studies of the genus Oryza. In: Rice genetic cytogenet. IRRI. Elsevier, Amsterdam: 15-21. [ Links ]
25. THE OFFICE OF THE GENE TECHNOLOGY REGULATOR (TOGTR). 2005. The biology and ecology of rice (Oryza sativa L.) in Australia. Department of health and ageing. Australian Goverment, 30 pp. [ Links ]
26. VAUGHAN, D.A. 1994. The wild relative of rice. A genetic Handbook. IRRI, Manila, Phillipines. [ Links ]
27. WATANABE, Y. 1997. Genomic constitution of genus Oryza. In: T. Matsuo, Y. Futsuhara, F. Kikuchi and H. Yamaguchi eds. Science of the rice plant. Food and Agriculture Policy Research Center, Tokyo. [ Links ]
