










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgronomía Tropical
versión impresa ISSN 0002-192X
Agronomía Trop. vol.62 no.1-4 Maracay dic. 2012
Índice valor de importancia, diversidad y similaridad florística de especies leñosas en tres ecosistemas de los llanos centrales de Venezuela
Importance value index, diversity and floristic similarity of woody species in three ecosystems of the llanos centrales of Venezuela
Pedro E. Soler 1, José L. Berroterán 2, José L. Gil 3 y Rafael A. Acosta 3
1 Profesor. Universidad Rómulo Gallegos (UNERG). San Juan de los Morros 2301, Guárico. Venezuela.
2 Profesor. Universidad Central de Venezuela (UCV). Facultad de Ciencias. Caracas 1053, Distrito Capital. Venezuela.
3 Investigadores. Instituto Nacional de Investigaciones Agrícolas (INIA-CENIAP e INIA Guárico). Venezuela. Correos electrónicos: pedrosoler75@hotmail.com, berroteranjl@gmail.com, jgil@inia.gob.ve
RESUMEN
Con el objeto de analizar la vegetación leñosa de un bosque deciduo tropical, un arbustal y una sabana arbolada, ubicados en los llanos altos centrales de Venezuela, se delimitaron dos áreas muestrales de 400 m² por tipo de vegetación con cuatro parcelas de 100 m2 cada una, donde se censaron las plantas con diámetro a la altura del pecho ≥ 1 cm. La composición florística se determinó mediante el índice valor de importancia (IVI), la similaridad mediante el índice de Jaccard y la diversidad y equitatividad mediante los índices de Shannon-Weaver y Simpson. Según el IVI, en el bosque las especies más importantes fueron Bourreria cumanensis (59,9), Arrabidaea pubescens (37,2), Myrospermun frutescens (22,8), Randia spinosa (21,7) y Guazuma ulmifolia (19,9); en el arbustal Combretum fruticosum (60,2), Mimosa tenuiflora (41,3), Guettarda divaricata (38,3) y Arrabidaea pubescens (34,4); en sabana Byrsonima crassifolia (74,8), Casearia zyzyphoides (57,2) y Curatella americana (50,1). En la vegetación de bosque, arbustal y sabana, la diversidad de Shannon fue 2,55; 2,02 y 2,09 y Simpson 8,7; 5,30 y 5,18. La equitatividad fue 0,80; 0,72 y 0,71 en Shannon y 0,37; 0,32 y 0,28 en Simpson. La similaridad entre bosque y arbustal fue 0,39; entre arbustal y sabana 0,21 y entre bosque y sabana 0,18. El bosque fue el más equitativo al presentar un número menor de especies dominantes. La similaridad florística fue baja en todos los casos, la mayoría de las especies presentes en el bosque no se encontraron en la sabana.
Palabras clave: llanos altos centrales, arbustal, bosque, sabana, vegetación leñosa, vegetación nativa.
ABSTRACT
With the objective to analyze the woody vegetation of a deciduous tropical forest, a shrubland, and a wooded savanna located at the high central plains of Venezuela, two sampling areas of 400 m² were delimited by vegetation type, with four plots of 100 m2 each, where a census of plants with diameter at breast height ≥ 1cm was taken. The floristic composition was determined by the importance value index (IVI), the similarity, using the Jaccard index, and the diversity and evenness using the Shannon-Weaver and Simpson indexes. According to IVI, the most important forest species were Bourreria cumanensis (59.9), Arrabidaea pubescens (37.2), Myrospermun frutescens (22.8), Randia spinosa (21.7), and Guazuma ulmifolia (19.9). In the shrubland, Combretum fruticosum (60.2), Mimosa tenuiflora (41.3), Guettarda divaricata (38.3), and Arrabidaea pubescens (34.4), and at the savanna, Byrsonima crassifolia (74.8), Casearia zyzyphoides (57.2), and Curatella americana (50.1). In the forest, shrubland and savanna, Shannon diversity indexes were 2.55, 2.02, and 2.09, respectively, while Simpson were 8.7, 5.30 and 5.18. The evenness were 0.80, 0.72 and 0.71 for Shannon and 0.37, 0.32 and 0.28 for Simpson. The index of similarity between forest and shrubland was 0.39, between shrubland and savanna 0.21, and between forest and savanna 0.18. Vegetation was very heterogeneous in terms of diversity. The forest was the most equitable, because had fewer dominant species. The floristic similarity was low in all cases. Most forest species did not occur in the savanna.
Key words: high central plains, forest, native vegetation, savanna, shrubland, woody vegetation.
Recibido: 20/03/2012 Aprobado: 26/02/2013 25
INTRODUCCIÓN
La vegetación de los llanos venezolanos está constituida por una mezcla de elementos florísticos y un interesante mosaico de diferentes tipos de vegetación (bosques, sabanas y arbustales), lo que está asociado con el paisaje, las características del suelo y las relaciones hídricas. La región de los llanos de Venezuela abarca una extensión de 237.000 km², representando el área con mayor potencialidad para la producción agropecuaria nacional y con la más alta superficie de la zona de vida del bosque seco tropical (Berroterán, 1994).
Según Fudena (2003), la flora vascular de los llanos comprende 3.137 especies agrupadas en 185 familias y 1.092 géneros, lo que representa el 22,3% de la flora total del país. El bosque seco tropical es muy heterogéneo en cuanto a la diversidad de especies vegetales (Ewel et al., 1976). La composición florística varía para cada estrato y dentro de éste, pueden existir varias asociaciones estrechamente relacionadas (Berroterán, 1994; Baldizán, 2004). La presencia de especies exclusivas, se puede relacionar con facilidades competitivas en condiciones favorables y limitaciones de adaptabilidad y competencia ínter específica en cada estrato (Berroterán, 1994). En el bosque deciduo tropical existe una gran diversidad de familias botánicas, entre las que destacan Mimosaceae, Fabaceae, Caesalpiniaceae, Bignoniaceae, Boraginaceae, Flacourtiaceae y Malvaceae (Baldizán et al., 2006).
Berroterán (1994) indica que los bosques deciduos presentan bajos índices de similitud con las asociaciones florísticas de matorrales y sabanas. Las asociaciones de las sabanas poco arboladas presentan baja similitud con todas las demás asociaciones florísticas a excepción del matorral ralo. Sin embargo, en estos ecosistemas se han identificado una gran diversidad de especies arbóreas y arbustivas con alto potencial para la alimentación animal en sistemas silvopastoriles, o como bancos de proteína en diferentes zonas y condiciones edafoclimáticas.
En algunos bosques del llano alto, se observan especies de valor forrajero, tales como el cují (Acacia macracantha), quiebra jacho (Chloroleucon mangense), dividivi (Caesalpinia coriaria), palma (Copernicia tectorum), cañafístola (Senna atomaria), guácimo (Guazuma tomentosa) y caro-caro (Enterolobium cyclocarpum), que conforman el 76% del total de los elementos arbóreos de la comunidad vegetal (Casado et al., 2001).
El área de los llanos altos centrales venezolanos es una zona de alta relevancia para la economía del país, de allí su importancia para el estudio de su vegetación, sobre todo en los actuales momentos, cuando el uso de tecnologías para la producción de monocultivos está introduciendo cambios susceptibles de generar el deterioro general de los sistemas. En este sentido, es necesario conocer y analizar la composición florística de esta importante región del país en función de establecer algunas estrategias mínimas de funcionamiento y sostenibilidad.
MATERIALES Y MÉTODOS
El estudio se realizó en la subregión natural de los llanos altos centrales de Venezuela, ubicada en la zona de vida, denominada bosque seco tropical, en las coordenadas 9° 14' 6'' y 9° 19' 57'' N y entre 67° 1' 52'' y 66° 58' 44'' O con una altitud de 180 m s. n. m. Políticamente, el área comprende el municipio Mellado del estado Guárico, a 12 km de la población El Sombrero, corresponde a una altiplanicie disectada con relieve suavemente ondulado a ondulado, con formaciones de bosque estacional y sabanas bien drenadas. Los suelos presentan textura gruesa, con niveles medios de materia orgánica (31 g kg-1) y pH moderadamente ácido (5,7) en el bosque y fuertemente ácido (5,2) en el arbustal y la sabana.
La zona presenta un clima tropical estacional subhúmedo (Mogollón y Comerma, 1994) con mesoclima semiárido, precipitación media anual de 1.085 mm con variaciones desde 616 hasta 2.200 mm. Comprende un período húmedo desde mayo a noviembre y un período seco desde diciembre hasta abril, con pedoclima de régimen ústico (López, 1993). La temperatura anual promedio es 27,2 °C con máxima de 34,2 °C y mínima de 21,0 °C. La humedad relativa promedio es 72% con mínima de 47 a 51% en los meses de marzo y abril y máxima de 83 a 87% en los meses húmedos de julio y agosto (Berroterán, 1994).
El estudio de la vegetación leñosa de las comunidades de bosque, arbustal y sabana arbolada, se realizó mediante la descripción de áreas muestrales, haciendo un análisis cuantitativo y cualitativo de la composición, diversidad y similaridad florística de los ecosistemas.
Se delimitaron dos áreas muestras por tipo de vegetación de 400 m² (40 x 10 m) cada una, divididas en cuatro parcelas de 100 m2, ubicadas en áreas que se consideraron representativas de la fisonomía de la vegetación. Se censaron todos los individuos con diámetro a la altura del pecho (dap) ≥ 1 cm.
Composición florística
La composición florística se determinó mediante la cuantificación del índice valor de importancia (IVI) de las especies; este consiste en la sumatoria de los valores relativos de densidad, frecuencia y dominancia e indica la importancia ecológica relativa de las especies de plantas en una comunidad (Curtis, 1959; Finol, 1971, 1976; Mueller-Dombois y Ellenberg, 1974; Matteucci y Colma, 1982). Los parámetros utilizados fueron los siguientes:
Índice de valor de importancia (IVI):
IVIi = Ai + Di + fii = especies de la comunidad, 1...n Abundancia: número de individuos por especie que se encuentran en la comunidad: Abundancia (A): Ai = Ni / S ARi = (Ai / Σ Ai) * 100 i = 1...n
Donde AR = abundancia relativa de la especie i respecto a la abundancia total, Ni = número de individuos de la especie i, S = superficie (ha) e i = especies de la comunidad, 1...n Dominancia: una especie es dominante cuando tiene una gran influencia sobre la composición y forma de la comunidad. Son especies de gran éxito ecológico y relativamente abundante dentro de la comunidad.
Dominancia (D): Di = Abi / S DRi = (Di / Σ Di) * 100 i = 1 n
Donde Ab = sección del fuste a 1,3 m de altura (m2), DR = dominancia (densidad) relativa de la especie i respecto de la dominancia total de la comunidad, i = especies de la comunidad, 1...n y S = superficie (ha).
Frecuencia: es el número de veces que una especie se presenta en una cantidad dada en parcelas o puntos de muestreo. Se evaluó la contribución de cada especie a la constitución de la comunidad mediante la fórmula: FCi = ni / Σni * 100, donde: FC = frecuencia centesimal de la especie i, ni = número de unidades de muestreo donde se encuentra la especie i y Σni = sumatoria del número de unidades de muestreo en las que encuentra la especie i.
Diversidad de especies
Luego de identificar las especies en cada tipo de vegetación, fueron calculados los índices de diversidad más comúnmente utilizados en ecología: Shannon-Wiener (H) y Simpson (D), así como Hill (1973) señala los correspondientes índices de equitatividad (J y E). La diversidad como valor único combina los parámetros de riqueza específica y equitatividad, factores fundamentales que definen la diversidad de una comunidad.
El índice de Shannon-Wiener considera no sólo el número de especies sino su representación (cuantos individuos por especie). Este índice requiere que todas las especies estén representadas en la muestra y es muy susceptible a la abundancia (Magurran, 1988). La mayor limitante es que no contempla la distribución de las especies en el espacio.
El índice de Shannon-Wiener (H) procede de la teoría de la información y mide la diversidad como:
H = -Σpi.ln (pi) pi = ni /N
Donde ni es el número de individuos de la especie i y N es la abundancia total de las especies. El valor de H se encuentra acotado entre 0 y ln(s), tiende a cero en comunidades poco diversas y es igual al logaritmo de la riqueza específica en comunidades de máxima equitatividad. La riqueza específica se midió como el número de especies presentes en una comunidad y la equitatividad como la abundancia de la especie dominante (Magurran, 1988), según la fórmula: equitatividad (J) = H/Hmáx = H/ln s.
La equitatividad se acerca a cero cuando una especie domina sobre todas las demás en la comunidad y se acerca a 1 cuando todas las especies comparten abundancias similares.
El índice de Simpson varía inversamente con la heterogeneidad; por ejemplo, los valores del índice decrecen o aumentan según aumente o decrezca la diversidad. Es en realidad un índice de dominancia, sobrevalora las especies más abundantes en detrimento de la riqueza total.
El índice de Simpson (D) mide la diversidad como D = Σ1/(pi2).
El valor de D se encuentra acotado entre 0 y s, tiende a cero en comunidades poco diversas, y es igual a la riqueza específica (s) en comunidades de máxima equitatividad (E = Σ1/(s * pi2).
Similaridad florística
La similaridad florística de las comunidades de bosque, arbustal y sabana, se evaluó mediante el índice de Jaccard (J), modificado por Ellenberg (1956), que es exclusivamente cualitativo y no considera el grado de participación de cada especie en la dominancia ecológica, donde el valor más cercano a 1 indica mayor semejanza (Magurran, 1988). Este índice se define mediante la ecuación: Índice de Jaccard: Jij = C / S1+ S2 - C Donde: S1 = N° de especies presentes en la zona 1 S2 = N° de especies presentes en la zona 2 C = N° de especies que están presentes en las dos muestras.
Como se puede observar es el cociente de la intersección entre la suma de todo lo no común. Es al contrario del índice de inclusión, un índice simétrico, y como aquel, sus valores oscilan entre 0 y 1. Cuando la intersección es nula, Jij = 0, y cuando los conjuntos son idénticos, Jij = 1.
RESULTADOS Y DISCUSIÓN
Composición florística
En el área estudiada se identificaron 54 especies pertenecientes a 48 géneros y 26 familias. Se observó la dominancia de las familias Fabaceae (14,8%), Caesalpiniaceae (14,8%), Mimosaceae (11,1%), Rubiaceae (7,4%), Bignoniaceae (5,6%), Malpighiaceae (5,6%) y Boraginaceae (5,6%). El 65% de las especies pertenecen a siete familias, el resto (35%) a 19 familias representadas por una especie cada una (ver Figura).
La tendencia a concentrar en pocas familias una alta proporción de especies, también fue observada en la misma zona por Berroterán (1994) y Baldizán et al. (2006), lo cual indica que la riqueza de especies incide directamente en la dominancia de las familias de plantas (Gallegos et al., 2002; Granda y Guamán, 2006).
En el bosque las especies de mayor IVI fueron Bourreria cumanensis, Arrabidaea pubescens, Myrospermun frutescens, Randia spinosa, Guazuma ulmifolia y Guettarda divaricata con valores de 59,9; 37,2; 22,8; 21,7; 19,9 y 17,7, respectivamente. Estas seis especies sumaron 60% del total del IVI. Otras siete especies tuvieron un valor de importancia entre 13 y 6,8; mientras que 21 (64%) especies reportaron un IVI entre 5,5 y 0,8. En muchas de estas especies, alrededor del 24% de los individuos del bosque, presenta una tendencia notable de baja abundancia y frecuencias relativas (Cuadro 1).
Como se muestra en el Cuadro 2, en el arbustal las especies con el mayor IVI fueron Combretum fruticosum (60,2), Mimosa tenuiflora (41,3), Guettarda divaricata (38,3) y Arrabidaea pubescens (34,4). Estas cuatro especies fueron las más importantes con 58% del total del IVI.
Otras especies como Senna atomaria, Caesalpinia coriaria, Platymiscium pinnatun y Bahuinia megalandra presentaron un valor de importancia entre 25,1 y 15,3. El resto (65% de las especies) presentaron valores muy bajos entre 0,5 y 10,2.
En el Cuadro 3 se puede observar que en la sabana arbolada, las especies de mayor IVI fueron Byrsonima crassifolia (74,8), Casearia zyzyphoides (57,2) y Curatella americana (50,1). Estas tres especies sumaron 60,6% del total del IVI. Otro 22,4% correspondió a cinco especies de menor importancia: Godmania macrocarpa (17,1), Randia spinosa (15,1), Miconia albicans (12,1), Arrabidaea pubescens (11,7) y Strychnos fendleri (11,1). Las restantes 16 especies (67%), presentaron valores entre 0,5 y 8,5.




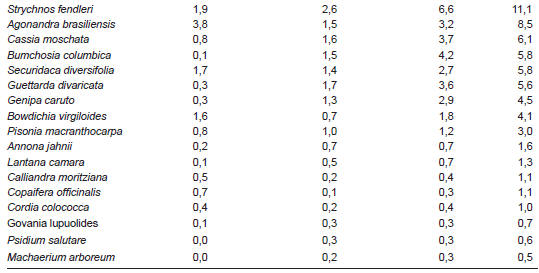
Los resultados permiten afirmar que los ecosistemas de bosque, arbustal y sabana arbolada presentaron grandes diferencias en la importancia ecológica de las especies, tal es el caso de Bourreria cumanensis y Arrabidaea pubescens en el bosque; Combretum fruticosum, Mimosa tenuiflora y Guettarda divaricada en el arbustal y Byrsonima crassifolia, Casearia zyzyphoides y Curatella americana en la sabana. Estas especies desempeñan un papel importante en la tipología de vegetación. Otras especies como Arrabidaea pubescens y Guettarda divaricada aparecieron con valores importantes en el bosque y el arbustal.
Los bajos valores del IVI en la mayoría de las especies indican que son especies de menor dominio florístico. Berroterán (1994) y Baldizán (2004) consideran que la composición florística varía para cada tipo de vegetación y dentro de estas pueden existir varias asociaciones estrechamente relacionadas. Se corrobora la afirmación de Ewell et al. (1976) quienes señalan que el bosque seco tropical es muy heterogéneo en cuanto a la diversidad de especies botánicas.
Diversidad de especies
La diversidad en una comunidad vegetal se puede definir como un conjunto de especies, donde cada una se encuentra representada por cierta cantidad de individuos, es decir, cada especie tiene un valor de abundancia que la caracteriza (Begon et al., 1996).
En el bosque, el arbustal y la sabana arbolada, se observaron 34, 23 y 24 especies representadas por 3158, 5010 y 6582 plantas ha-1, respectivamente.
En el bosque, las especies más abundantes fueron Arrabidaea pubescens, Bourreria cumanensis, Myrospermun frutescens, Randia spinosa, Guettarda divaricada, Bumchosia columbica y Pisonia macranthocarpa, con 575, 575, 413, 325, 200, 113 y 100 plantas ha-1, respectivamente.
En el arbustal sobresalieron Guettarda divaricada, Combretum fruticosum, Arrabidaea pubescens y Senna atomaria, con 1100, 1038, 763 y 688 plantas ha-1, respectivamente. En la sabana predominó Casearia zyzyphoides, Byrsonima crassifolia, Curatella americana y Godmania macrocarpa, con 2275, 1238, 838 y 388 plantas ha-1, respectivamente. Alrededor de 20% de las especies encontradas representaron 72% de los individuos de estas comunidades. El índice de diversidad de Shannon (H) mostró valores de 2,55; 2,02 y 2,09 para la vegetación de bosque, arbustal y sabana, respectivamente (Cuadro 4). El valor del índice de diversidad del bosque fue más alto, mientras que los valores del arbustal y la sabana fueron muy similares.
El índice de diversidad de Simpson (D) que mide tanto la dominancia como la riqueza tuvo un valor más elevado en el bosque (8,72) que el arbustal (5,30) y la sabana (5,18). El índice de equitatividad para Shannon (J) en la vegetación de bosque (0,80), arbustal (0,72) y sabana (0,71) mostraron valores bastante similares. Asimismo, igual comportamiento tuvo el índice de equitatividad de Simpson (E) en el bosque (0,37), el arbustal (0,32) y la sabana (0,28), respectivamente (Cuadro 4).
La equitatividad de Shannon mide la relación entre la diversidad observada y la diversidad máxima, se aproximó a 1 en el arbustal y la sabana. Sin embargo, el bosque tuvo un índice de equitatividad mayor. Esto sugiere que el arbustal y la sabana son comunidades con menos equidad que el bosque, lo cual indica que en estas comunidades existen unas especies mejor representadas que otras, mientras que en el bosque la mayoría de especies poseen una cantidad similar de individuos.
En el caso del bosque, el alto valor del índice de Simpson indica una mayor riqueza de especies con respecto al arbustal y la sabana arbolada, así como la presencia de un número menor de especies dominantes, esto apoya lo sugerido por el índice de equitatividad de Shannon (Magurran, 1988).
Los valores obtenidos para el índice de equitatividad de Simpson (E) en el bosque, arbustal y la sabana fueron bajos, lo que evidencia la baja uniformidad en la distribución de individuos entre las especies dominantes de estos ecosistemas. Esto sugiere una tendencia donde los individuos de las especies dominantes presentan una distribución ligeramente más equitativa en el orden: bosque > arbustal > sabana.
Similaridad florística
La similaridad florística se basa en la incidencia de las especies en cada ambiente (presencia o ausencia) y el resultado se puede visualizar como la proporción o porcentaje de especies compartidas, permitiendo conocer en forma detallada la semejanza entre pares de comunidades florísticas que previamente han sido definidas. Las comunidades nativas de bosque y arbustal presentaron un índice de similaridad de 0,39. Esta similaridad corresponde a 18 especies exclusivas del bosque, siete del arbustal y 16 compartidas (Cuadro 5). Entre bosque y sabana, la similaridad fue 0,18; con 25, 15 y nueve especies del bosque, sabana y compartidas (Cuadro 6). Asimismo, entre sabana y arbustal, el índice fue 0,21; con 16 especies de sabana, 15 arbustal y ocho compartidas (Cuadro 7).
Tomando en cuenta que el valor más cercano a 1 indica mayor semejanza (Magurran, 1988), se puede decir que existió baja similaridad florística entre todas las formaciones estudiadas (Cuadro 8). En todos los casos, los valores estuvieron alejados de 1 (0,18-0,39), siendo las áreas más relacionadas el bosque y el arbustal (índice de Jaccard = 0,39) con 16 especies compartidas; los otros pares de tipos de vegetación mostraron valores de similaridad menor. Estas bajas similaridades indican que la mayoría de las especies de una comunidad no se presentan en la otra.






La baja similaridad florística observada entre las áreas de estudio, puede ser explicada por las diferentes condiciones edáficas presentes en cada tipo de vegetación (reacción del suelo, aluminio intercambiable, pedregosidad, profundidad, entre otras). Los suelos de estas sabanas presentan horizonte pedregoso (70%), pH fuertemente ácido y altos niveles de aluminio intercambiable, lo que condiciona la existencia del tipo de vegetación, caracterizada por especies que toleran diferencialmente las condiciones ecológicas presentes (Soler, 2010).
Existen otros factores condicionantes (Muñoz et al., 2004), como lugares conectados por el viento que presentan floras parecidas independientemente de la distancia que los separe, y lugares muy próximos, pero con pocas especies en común, porque no hay vientos que los conecte. En todo caso, los bosques, los arbustales y las sabanas están asociados espacialmente. No obstante, los bosques deciduos presentaron bajos índices de similitud con las asociaciones florísticas
CONCLUSIONES
Se observó la dominancia de las familias Fabaceae, Caesalpiniaceae, Mimosaceae, Rubiaceae, Bignoniaceae, Malpighiaceae y Boraginaceae. Las especies con mayor IVI fueron Bourreria cumanensis, Arrabidaea pubescens, Myrospermun frutescens, Randia spinosa y Guettarda divaricata en el bosque; Combretum fruticosum, Mimosa tenuiflora, Guettarda divaricada y Arrabidaea pubescens en el arbustal; Byrsonima crassifolia, Casearia zyzyphoides y Curatella americana en la sabana.
En el arbustal y la sabana existió el mayor número de especies dominantes y excluyentes que compiten por el espacio, lo que condiciona la presencia de otras especies. Sin embargo, el bosque es más heterogéneo, es decir, el arbustal y la sabana son comunidades con menos equidad que el bosque.
Existe una mayor riqueza de especies en el bosque con respecto al arbustal y la sabana arbolada. La similaridad florística fue baja en todos los casos; sin embargo, fue más alta entre bosque y arbustal. La mayoría de las especies del bosque no se presentaron en la sabana.
LITERATURA CITADA
1. Baldizán, A. 2004. Producción de biomasa y nutrimentos de la vegetación del bosque seco tropical y su utilización por rumiantes a pastoreo en los Llanos Centrales de Venezuela. Tesis doctorado. Maracay, Ven. Universidad Central de Venezuela. Facultad de Agronomía. 288 p. [ Links ]
2. Baldizán, A., C. Domínguez, D. García, E. Chacón y L. Aguilar. 2006. Metabolitos secundarios y patrón de selección de dietas en el bosque deciduo tropical de los llanos centrales venezolanos. Zootecnia Trop. 24(3):213-232.
3. Begon, M., M. Mortimer and D. Thompson. 1996. Population Ecology. A unified study of animal and plants. 3ra ed. Blackwell Science Publishing. Oxford, Inglaterra. 247 p.
4. Berroterán, J. L. 1994. Ecología de sistemas nativos y agroecosistema maíz en los Llanos Altos Centrales de Venezuela. Tesis doctorado. Maracay, Ven. Universidad Central de Venezuela. Facultad de Agronomía. 445 p. [ Links ]
5. Casado C., N. Benezra, O. Colmenares y N. Martínez. 2001. Evaluación del bosque deciduo como recurso alimenticio para bovinos en los llanos centrales de Venezuela. Zootecnia Trop. 19(2):139-150.
6. Curtis, J. 1959. The vegetation of Wisconsin. An ordination of plant communities. Univ. of Wisconsin Press. Madison. EUA. 657 p. [ Links ]
7. Ellenberg, H. 1956. Aufgaben und Methoden der Vegetationskunde. Eugen Ulmer, Stuttgart. Alemania. [ Links ]
8. Ewel, J., A. Madriz y J. Tosi. 1976. Zonas de Vida de Venezuela. Ministerio de Agricultura y Cría. 2da ed. Fondo Nacional de Investigaciones Agropecuarias. Caracas, Venezuela. 265 p. [ Links ]
9. Finol, H. 1971. Nuevos parámetros a considerarse en el análisis estructural de las selvas vírgenes tropicales. Rev. For. Ven. 13(21):29 42. [ Links ]
10. Finol, H. 1976. Estudio fitosociológico de las unidades 2 y 3 de la reserva forestal de Caparo, estado Barinas. Acta Bot. Ven. 10(1-4):15-103. [ Links ]
11. Fudena (Fundación para la Defensa de la Naturaleza). 2003. Programa de evaluación de la biodiversidad. Fudena. Disponible en línea: http://www.fudena.org.ve/proyectoslbio.htm (Consultado 7 febrero 2008). [ Links ]
12. Gallegos, A., E. Abundio, M. Morales y E. Hernández. 2002. Valor de importancia de especies arbóreas en un bosque tropical de la costa de Jalisco. Resúmenes V Congreso Mexicano de Recurso Forestales. Guadalajara, Jalisco. México. Disponible en línea: http://www.buenastareas.com/ensayos/Valor-De-Importancia-De-Especies-Arb%C3%B3reas/2868416.html (Consultado octubre 2011).
13. Granda, V. y S. Guamán. 2006. Composición florística, estructura, y etnobotánica del bosque seco Algodonal. Lyonia, 11(2) Disponible en línea: http://www.lyonia.org/viewArticle.php?articleID=395 (Consulta: 23 octubre 2007).
14. Hill, M. O. 1973. Diversity and evenness: a unifying notation and its consequences. Ecology. 54(2):427-432. [ Links ]
15. López, M. 1993. Diagnóstico físico y alternativas conservacionistas de tres unidades de tierra de los llanos altos centrales en el estado Guárico. Trabajo de Grado M.Sc. Maracay, Ven. Universidad Central de Venezuela. Facultad de Agronomía. 93 p. [ Links ]
16. Magurran, A. E., 1988. Ecological diversity and its measurement. Princeton University Press, New Jersey. 179 p. [ Links ]
17. Matteucci, S. D. y A. Colma. 1982. Metodología para el estudio de la vegetación. Secretaría General de la Organización de Estados Americanos. Programa Regional de Desarrollo Científico y Tecnológico Washington, EUA. 72 p.
18. Mogollón, L. y J. Comerma. 1994. Suelos de Venezuela. Palmaven, Ex Libris. Caracas, Venezuela. 267 p.
19. Mueller-Dumbois, D. and H. Ellenberg. 1974. Aims and methods of vegetation ecology. John Wiley and Sons, New York. EUA. 547 p.
20. Muñoz, J., A. M. Felicísimo, F. Cabezas, A. R. Burgaz and I. Martínez. 2004. Wind as a long-distance dispersal vehicle in the southern hemisphere. Science. 304(5674):1.144-1.147.
21. Soler, P. 2010. Evaluación del potencial forrajero de la vegetación nativa e intervenida en un área de los Llanos Altos Centrales del estado Guárico, Venezuela. Tesis doctorado. Maracay, Ven. Universidad Central de Venezuela. Facultad de Agronomía.185 p. [ Links ]
