











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkActa Botánica Venezuelica
versión impresa ISSN 0084-5906
Acta Bot. Venez. v.28 n.2 Caracas 2005
ASPECTOS MORFOLÓGICOS Y ANATOMÍA FOLIAR DE LAS ESPECIES DE GNETUML. (GNETACEAE-GNETOPHYTA) PRESENTES EN VENEZUELA
Morphological aspects and leaf anatomy of the species of Gnetum L. (Gnetaceae-Gnetophyta) present in Venezuela
Damelis JÁUREGUI y Carmen BENÍTEZ DE ROJAS
Instituto de Botánica Agrícola, Facultad de Agronomía, Universidad Central de Venezuela. Apdo. Postal 4579. Maracay, Aragua. jaureguid@agr.ucv.ve. cbenitez@telcel.net.ve
RESUMEN
Se realizó una caracterización morfológica general y anatómica foliar de seis especies de Gnetum que crecen en Venezuela con el fin de definir caracteres de valor diagnóstico. Se hicieron observaciones bajo un microscopio estereoscópico y para el estudio anatómico se aplicaron los protocolos convencionales para microscopía óptica. Las especies de Gnetum se diferencian por características de la corteza de los tallos, el largo de los entrenudos, el tamaño de las hojas, el color de la semilla y el número y forma de los cotiledones. Anatómicamente se distinguen por: presencia de hipodermis; cristales; densidad, disposición y grado de engrosamiento de las paredes de las células esclerenquimáticas; y el número de haces vasculares en la vena media. Se presenta una clave basada en las características morfoanatómicas.
Palabras clave:Anatomía, Gnetum, Hoja, Morfología
ABSTRACT
Morphological and anatomical information for six species of Gnetum were investigated to understand which features are important for taxonomic delimitations. Morphological characterization and observations were made using stereoscopic microscopy, following the conventional methodology for taxonomic studies. For anatomic studies, the material was prepared using traditional methods, and examined with a compound microscope. Morphologically, the species are distinguished by stem cortex surface, length of internodes, size of leaves, seed color and form and number of cotyledons. Distinctive anatomical characters are: the presence of hypodermis; crystals; the density, and degree thickness of the cell walls of the sclerenchymatous cells in the mesophyll, and the number of vascular bundles in the midrib. An artificial key based on morphoanatomical characters is presented.
Key words:Anatomy, Gnetum, Leaf,Morphology
INTRODUCCIÓN
El género Gnetum L. es el único dentro de la familia Gnetaceae (Gnetophyta). La importancia filogenética de este género se fundamenta en el hecho de que ha sido propuesto como posible progenitor de las angiospermas; sin embargo, la falta de "pollenkitt" (sustancia lipoidal) en la superficie del grano de polen en este taxón al igual que en las gimnospermas extintas, representa una condición por la que muchos autores no admiten a las Gnetophyta como progenitoras de las angiospermas (Hesse 1984, en Gifford & Foster 1988). Según Judd et al. (1999), las Gnetophyta constituyen uno de los cuatro grupos monofiléticos de plantas con semillas, muy evolucionado y con alto grado de especialización, condición que dificulta su comparación con otras gimnospermas.
Gnetum reúne aproximadamente 39 especies pantropicales, de las cuales 37 son lianas (Price 1996). Stevenson & Zanoni (1991) indican que en el Neotrópico se localizan entre 8 y 10 especies. Por otra parte, Markgraf (en Price 1996) señala siete especies en Centro y Sur América: G. camporum (Markgr.) D.W. Stev. & Zanoni, G. leyboldii Tul., G. nodiflorum Brongn., G. paniculatum Spruce ex Benth., G. schwackeanum Taub. ex Markgr., G. venosum Spruce ex Benth. y G. urens (Aubl.) Blume, todas pertenecientes a la Sección Gnetum, Subsección Araeognemones Markgr. (Price 1996). Estas especies, excluyendo a G. venosum, están presentes en Venezuela y en particular G. camporum es endémica para el país (Stevenson 1995).
Las especies del género Gnetum se reconocen por su hábito trepador lianoide, voluble y leñoso, con inflorescencias ramificadas o simples, con flores unisexuales arregladas en verticilos de espigas o panículas sobre los ejes del estróbilo. Las hojas son simples, glabras, opuestas, decusadas, papiráceas o coriáceas, siempreverdes. El patrón de venación es del tipo pinnado broquidódromo (Hickey 1973) con las venas primarias formando entre ellas arcos prominentes; dicho patrón se corresponde con el de algunas dicotiledóneas. Las semillas son semejantes a drupas de color amarillento. En relación con su anatomía foliar sólo se han registrado algunos caracteres en G. gnemon L.(Maheswari & Vasil 1961), G. ula Brongn. (Maheswari & Vasil 1961; Kausik 1974) y en G. africanum Welw. (Duthie 1912).
Considerando la importancia filogenética y florística del género Gnetum, este trabajo surge con el interés de aportar información de la morfología y anatomía foliar en las especies de este taxón en Venezuela y además, definir caracteres morfoanatómicos diagnósticos que pudiesen contribuir a su delimitación taxonómica.
MATERIALES Y MÉTODOS
Para realizar esta investigación se utilizó material fresco de G. urens colectado en el área Norte de la Gran Sabana Norte, Salto Kawi, estado Bolívar; en el caso de las otras especies se trabajó con material herborizado depositado en los herbarios MY, MER, TFAV y VEN. Se utilizaron dos o más hojas provenientes, en lo posible, de colecciones diferentes. En la Tabla 1 se presentan las diferentes colecciones revisadas.

Para la caracterización morfológica se hicieron observaciones bajo microscopio estereoscópico, siguiendo la metodología convencional en estudios taxonómicos. Para la caracterización anatómica se tomaron trozos de la parte media de las hojas y de los pecíolos los cuales se hidrataron previamente, siguiendo la técnica de Peña & Saralegui (1982). Una vez hidratados, se seccionaron a mano alzada, los cortes obtenidos se tiñeron con azul de toluidina acuosa 0,05% (OBrien & McCully 1981) y se montaron en agua-glicerina (V:V). Con el fin de separar las epidermis del resto de tejidos que conforman las hojas, se colocaron porciones de la parte media de la lámina foliar en solución de Jeffrey, por períodos variables dependiendo de la consistencia de las mismas; las porciones de mesofilo obtenidas se dejaron hasta maceración total con el fin de estudiar la forma de las células esclerenquimáticas presentes.
Para la ilustración de las especies se seleccionaron exsiccata como se indica en las leyendas de las mismas. La terminología empleada para clasificar los estomas se basó en Patel (1979).
RESULTADOS
Aspectos morfológicos
Gnetum camporum (Markgr.) D.W. Stev. & T. Zanoni, in Görts, Fl. Guianas, sér A, fam. 7 (Gnetaceae) 14. 1991 (Fig. 1).

Gnetum urens var. camporum Markgr., Acta Bot. Venez. 6: 371. 1972.
Trepadora leñosa de aproximadamente 2 m alto, tallos con la corteza externa lisa, entrenudos 17-28,5 cm de largo. Hojas muy coriáceas, verde-oliváceo oscuro en la haz, verde pálido en el envés, 4,5-6,5 x 3-4 cm, ovadas, muy fibrosas en ambas superficies, con las venas primarias y secundarias prominentes. Semillas ovoides, 2,5 – 2,8 x 1,5-2 cm, inmaduras verde-rojizas, cuando maduras castaño-violáceos, con dos cotiledones seccionados y fácilmente separables.
Material examinado: BOLÍVAR: Gran Sabana, carretera Fuerte Luepa-Santa Elena, 15/01/1994, N. Ramírez et al. 4751 (VEN).
Gnetum leyboldii Tul., Ann. Sci. Nat., Bot. sér. 4, 10: 117. 1858 (Fig. 2).
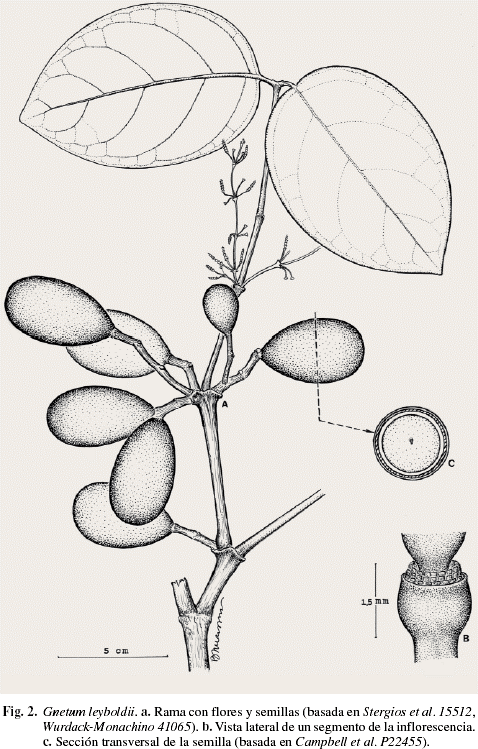
Trepadora leñosa, tallos articulados, con la corteza externa estriada, entrenudos 5-8,5 cm de largo. Hojas coriáceas, verde oscuro brillante en la haz, verde pálido en el envés, 10,5-22 x 6,5-11,5 cm. Semillas ovoides, 3,5-5 x 2-3 cm, inmaduras verdes, cuando maduras de color castaño-verdoso, con un cotiledón ocupando toda el área interna.
Material examinado: AMAZONAS: Selvas ribereñas del Río Emoni, 27/01/92, B. Stergios et al. 15512 (VEN). Bolívar: Municipio Raúl Leoni, 45 km al este de Los Pijiguaos, 06/89, Angel Fernández & L. Delgado 5706 (VEN); margen derecha del Río Cucurital, sector Wareipa, 370 m snm, 22/09/2000, L. Rodríguez & M. García 178 (VEN).
Gnetum nodiflorum Brongn., in Duperrey,Voy. Monde (Phan.) 12. 1829 (Fig. 3).

Trepadora leñosa, tallos lenticelados, con la corteza externa gruesa, tosca, entrenudos 4-11,5 cm de largo. Hojas coriáceas con el margen revoluto, verde oscuro en ambas superficies, 11-17,5 x 5,5-10 cm. Semillas ovoides, 3,5-4,5 x 1,5-2 cm, inmaduras verdes, cuando maduras amarillo-verdosas hasta púrpura, con un cotiledón ocupando toda el área interna.
Material examinado: AMAZONAS: Municipio Manapiare, 110 m snm, 10/04/2000, B. Milano & A. Fernández 01697 (VEN); Municipio Manapiare, Yutajé, 110 m snm, 26/07/1998, B. Milano et al. 136 (VEN); Segunda laguna en Río Pasimoni, 04/11/1994, B. Stergios et al. 16282 (VEN). Bolívar: Campo Cardona, 400 m snm, 04/08/1962, Ch. Brewer 32 (VEN).
Gnetum paniculatum Spruce ex Benth., Hookers J. Bot. Kew Gard. Misc. 8: 357, t.2, 3. 1856 (Fig. 4).

Trepadora leñosa voluble, 6-8 m alto, su porción basal por unos 4-6 m sumergida en agua negra durante la estación lluviosa, tallos lenticelados, con la corteza externa gruesa, entrenudos 4-10 cm de largo. Hojas grueso-coriáceas, verde oscuro en ambas caras, 9,5-16 x 5-11 cm. Semillas esféricas hasta subovoides, 3-4,5 x 2-4 cm, inmaduras verdes con tonalidades rojizas, con el episperma duro y el perfil estrellado, un solo cotiledón.
Material examinado: AMAZONAS: Confluencia de los Ríos Sipapo-Orinoco, Municipio Autana, 22/09/1996, A. Castillo S. 4003 (VEN); Municipio Autana, 28/01/1997, A. Castillo 4454 (VEN); Río Cuao, Municipio Autana, 29/01/1997, A. Castillo 4560 (VEN); Río Sipapo, Municipio Autana, 225 m snm, 15/04/1997, A. Castillo 4857 (VEN); Río Sipapo, entre boca de Cuao y Piedra Chamii, Municipio Autana, 225 m snm, 18/08/1997, A. Castillo 5413 (VEN); Río Sipapo, entre Raudal Caldero y Santa Teresita, Municipio Autana, A. Castillo 5740 (VEN); Márgenes altas Río Atacavi, 07/09/1960, E. Foldats 3761 (VEN); Municipio Atabapo, 28/05/1975, P. Berry 768 (VEN); Municipio Gitanea, 130 m snm, 11/2001, J.S. Wurdack & L.S Adderly 43480 (VEN). Bolívar: Distrito Sifontes, concesión minera Cristina Cuatro, 180 m snm, G. Aymard et al. 4149 (PORT); Carretera hacia el campamento La Felicidad-CVG, 26/09/1989, M. Colella & P. Valdéz 1538 (VEN).
Gnetum schwackeanum Taub. ex Markgraf, Bull. Jard. Bot. Buitenzorg sér 3,10: 450. 1930 (Fig. 5).

Liana, tallos laminados, láminas de corcho delgadas y aladas, de color amarillo claro, tallos engrosados justo arriba y debajo de los nudos, entrenudos 6-9,5 cm de largo. Hojas coriáceas, lustrosas en ambas superficies 11-12 x 6-5 cm. Semillas ovoides, 3,6- 4 x 2,2- 3 cm, con dos cotiledones y el episperma liso.
Material examinado: AMAZONAS: Departamento Atures, 90-100 m snm, 20/02/1985, G. Carnevali et al. 1708 (TFAV, VEN), Departamento Atabapo, 175-805 m snm, 12-14/10/1984, F. Delascio 12422 (TFAV, VEN); Sector El Peligro, 04/2000, B. Stergios et al. 18596 (TFAV).
Gnetum urens (Aubl.) Blume, Tijdschr. Natuurl. Gesch. Physiol. 1: 162. 1834 (Fig. 6).

Liana, trepando hasta 20 m de alto, muy ramificada, tallos con la corteza externa lisa, los nudos engrosados, entrenudos 6-10 cm de largo. Hojas coriáceo-cartáceas, verde oscuro brillantes en la haz, claro en el envés, 3,5-4,5 x 2,5-3 cm. Semillas ovoides, anaranjado intenso, 3-4,5 x 5,2-5,4 cm, con dos cotiledones fácilmente separables, episperma liso.
Material examinado: BOLÍVAR: Gran Sabana, 1.200 m snm, 12/08/01, C. Benítez de Rojas 6268 (MY).
Anatomía foliar
Lámina foliar
Las seis especies estudiadas presentan epidermis uniestratificada en ambas caras con células principalmente de contornos rectangulares (Fig. 7). La pared celular externa más la cutícula es más gruesa en G. urens y G. nodiflorum y más delgada en G. schwackeanum. Dicha pared más la cutícula se proyectan en forma irregular hacia el interior de las células en G. leyboldii, G. nodiflorumy G. paniculatum. En vista paradermal de la superficie adaxial (Fig. 8), se observa que casi todas las especies estudiadas presentan células con paredes rectas o levemente onduladas, visualizándose formas poligonales en las que prevalecen las de cuatro y cinco lados; sólo en G. schwackeanum se distinguen células de contornos ondulados (Fig. 7e). Cabe destacar que las paredes celulares son gruesas y con punteaduras evidentes en todas las especies. La vista paradermal de la superficie abaxial (Fig. 9), muestra células de paredes rectas o muy levemente onduladas con tres y cuatro lados en G. leyboldii, G. paniculatum y G. nodiflorum y onduladas en G. camporum, G. urens y G. schwackeanum (esta especie presenta el mayor grado de ondulación en sus paredes celulares); en estos tres últimos taxones las paredes celulares son más delgadas que en el resto de las especies. En todas las especies los estomas se localizan en la cara abaxial de las hojas (Fig. 9), ubicados inclusive sobre nervaduras. Los estomas, son predominantemente paracíclicos y en ocasiones duploparatetracíclicos, el primero se caracteriza por presentar una célula anexa a cada lado del estoma, de longitud similar a las oclusivas, pero sin cubrir las regiones polares de éstas, en el otro caso la diferencia radica en que existen dos células anexas a cada lado del estoma. En los taxa estudiados fue frecuente observar estomas gemelos, esto es que comparten células anexas.



Con relación al arreglo del mesofilo, todas las especies tienen hojas bifaciales, excepto G. leyboldii que presenta hojas isofaciales (Fig. 7b). El parénquima en empalizada está constituido por tres a cuatro estratos de células en G. Leyboldii (Fig. 7b), G. nodiflorum (Fig. 7c), G. paniculatum (Fig. 7d) y G. Urens (Fig. 7f); mientras que G. camporum (Fig. 7a) y G. schwackeanum (Fig. 7e) presentan sólo dos o tres capas. En todas las especies se observan de siete a ocho estratos de parénquima esponjoso con amplios espacios intercelulares, excepto en G. paniculatum donde este tejido es relativamente más compacto. Las hojas de mayor grosor se presentan en G. urens, mientras que las más delgadas se observan en G. schwackeanum. G. paniculatum es la única especie que en ocasiones presenta hipodermis, ésta se ubica en la cara adaxial de la lámina foliar y se caracteriza por presentar células de mayor volumen que las epidérmicas y con escaso contenido protoplasmático (Fig. 10c). Asimismo, G. paniculatum es la única especie con cristales en el mesofilo.

La presencia de células esclerenquimáticas, fibras y esclereidas (astroesclereidas y/o macroesclereidas) en el mesofilo es constante para las especies estudiadas (Fig. 7 y 10). En este sentido, G. leyboldii, G. nodiflorum y G. Paniculatum presentan mayormente grupos de macroesclereidas, dispuestas con su eje mayor paralelo al nervio medio de la hoja, asimismo presentan astroesclereidas abundantes distribuidas en el mesofilo, pero en G. nodiflorum se disponen perpendicularmente a la superficie mientras que, en las otras dos especies se arreglan tanto vertical como horizontalmente, siendo muy abundantes en G. leyboldii. Cabe destacar que en G. camporumy G. urens abundan fibras y macroesclereidas, dispuestas principalmente con su eje mayor perpendicular al nervio medio de la hoja; las astroesclereidas son escasas, en G. camporum las células presentan paredes secundarias muy engrosadas.
Los haces vasculares son colaterales cerrados, limitados por células parenquimáticas con paredes engrosadas y/o células esclerenquimáticas en todas las especies, a modo de una vaina (Fig. 7b-d, 10b, 10e), excepto G. camporum (Fig. 7a) y G. urens (Fig. 7f), que carecen de dicha vaina.
Nervadura central
La histología de la nervadura central es semejante en las seis especies (Fig. 11), excepto en el número de haces vasculares visibles. La epidermis es uniestratificada en ambas caras con células de contornos mayormente cuadrangulares, la pared celular externa más la cutícula es gruesa en ambas epidermis. Seguidamente está el parénquima con diversos grados de engrosamiento en sus células, abundantes células esclerenquimáticas, en mayor proporción en las proximidades del tejido vascular (cara abaxial), excepto en G. camporum donde hay relativamente menor cantidad de este tipo de células asociadas a los haces vasculares. El sistema vascular está constituido por tres a cuatro haces en G. urens y G. nodiflorum; cinco a seis en G. camporum, G. leyboldii y G. schwackeanum y seis a siete en G. paniculatum; dichos haces son colaterales cerrados, separados por células parenquimáticas con diverso grado de esclerotización y arreglados en forma de arco.

Pecíolo
Anatómicamente, los pecíolos son similares (Fig. 12), externamente se evidencia epidermis uniestratificada con la pared celular más la cutícula gruesa; en ciertas porciones se diferencia una peridermis de poco espesor. Seguidamente, parénquima con abundantes células esclerenquimáticas, en mayor densidad en la región abaxial; en G. paniculatum se observan cristales. En la porción central se localiza el tejido vascular, el cual está constituido por 5-7 haces colaterales cerrados, separados por parénquima con idioblastos esclerenquimáticos. Las regiones parenquimáticas son relativamente amplias en G. camporum, G. Schwackeanum y G. urens y más estrechas en G. leyboldii, G. nodiflorum y G. paniculatum.


DISCUSIÓN
Los resultados obtenidos indican que las especies de Gnetaceae presentes en Venezuela muestran una morfología y anatomía foliar muy semejante entre ellas.
Los caracteres morfológicos macroscópicos permiten distinguir las especies por el grosor y textura de la corteza de tallos, longitud de los entrenudos, tamaño de las hojas, color de la semilla, número y morfología de los cotiledones.
Con relación a los rasgos anatómicos foliares, se puede señalar que la presencia de células epidérmicas con contornos poligonales o casi así, con paredes rectas o ligeramente onduladas descrita en este trabajo para las especies G. leyboldii, G. nodiflorum y G. paniculatum, se corresponde con lo observado en las ilustraciones realizadas de las epidermis de G. gnemon (Maheswari & Vasil 1961), y G. ula (Maheswari &Vasil 1961; Kausik 1974). Asimismo coincide con las observaciones de Rodin (1966) quien reportó que en todas las especies por él estudiadas las células epidérmicas son irregulares y poligonales; no obstante, en este estudio se pudo hacer una caracterización más detallada de los contornos celulares, pudiendo concluir que este carácter permite separar las especies en dos grupos: el primero con células de paredes rectas y el otro con paredes onduladas en mayor o menor grado.
Todas las especies resultaron hipostomáticas; no obstante, Kausik (1974) reportó estomas escasos sobre la epidermis adaxial en G. ula, especialmente sobre la vena media. La presencia de estomas gemelos también fue observada por Maheswari & Vasil (1961) en G. gnemon y G. ula. Por otra parte, los estomas son del tipo sindetoquélico, atendiendo a la clasificación de Florin (1931, en Rodin 1966).
Las diferencias observadas en cuanto al número de capas de células del parénquima en empalizada, así como la presencia de hojas equifaciales en G. Leyboldii pueden asociarse con la exposición de las hojas a una mayor radiación lumínica. Duthie (1912) señaló que el mesofilo de G. africanum está constituido por un solo estrato de células en el parénquima en empalizada y que esta región está mayormente ocupada por parénquima esponjoso. En el mesofilo merece destacar como característica de posible valor diagnóstico la presencia de hipodermis y cristales únicamente en G. paniculatum.
Un rasgo importante de resaltar es la presencia de células esclerenquimáticas en el mesofilo, aspecto que fue considerado con detalle por Rodin (1966). Stevenson & Zanoni (1991) en su clave para separar las especies de Gnetum presentes en las Guayanas, utilizan como carácter de valor diagnóstico la presencia en las hojas de fibras subepidérmicas principalmente en la cara adaxial de las hojas. Estos autores separan a G. camporum y G. urens, de G. leyboldii, G. Nodiflorum y G. paniculatum, indicando que en las dos primeras especies no está presente este tipo de células. En el presente estudio se observa la presencia mayormente de fibras y esclereidas (macroesclereidas y astroesclereidas) para los seis taxones analizados, apreciándose variación en la densidad, distribución y disposición de dichas células. Por otra parte, Duthie (1912) indicó la presencia de una red de células esclerenquimáticas en G. africanum; dicha red está constituida por células estrelladas y fibras muy largas, ambas de paredes muy gruesas.
El estudio del sistema vascular del pecíolo parece indicar que en todas las especies existen entre 5-7 trazas vasculares. No obstante, Rodin (1966) reportó variación en este número, aunque cuatro de las especies presentaron el mismo rango obtenido en este trabajo. En el caso de G. nodiflorum y G. urens pareciera ocurrir una fusión de dichas trazas, una vez que entran al nervio medio de la hoja o quizás un poco antes. A este respecto, Rodin (1966) indicó que los haces vasculares del pecíolo continúan hacia la vena media de la hoja, sin que ocurra fusión entre ellas; sin embargo, la caracterización en este caso se hizo sólo con material diafanizado. Por su parte, Rodin & Paliwal (1970) reportaron un número impar de trazas, 5-7 en G. gnemon y 8 en G. ula. Estos autores indicaron además que las 5-7 trazas en G. gnemon se extienden hacia la vena media como una serie de haces longitudinales y que a niveles sucesivos dicotomizan, lo cual no ocurre en las especies estudiadas ya que en ellas se mantiene el mismo número o se reduce, como se señaló antes.
A nivel del pecíolo, una característica que permite distinguir las especies en dos grupos es la amplitud de las regiones parenquimáticas ubicadas entre los haces vasculares (regiones amplias y estrechas).
CONCLUSIONES
Los resultados obtenidos indican que las especies de Gnetum presentes en Venezuela muestran características morfológicas que permiten diferenciar las especies, tales como: tamaño de las hojas, color de la semilla, número y morfología de los cotiledones. En cuanto a la anatomía foliar, los taxa estudiados son muy semejantes entre sí, distinguibles sólo por algunas características, las cuales permitieron construir una clave; entre ellas se pueden citar: amplitud de radios que separan haces vasculares en el pecíolo, contornos de las células epidérmicas, número de haces vasculares que irrigan la nervadura central, presencia o ausencia de hipodermis y cristales y el patrón de distribución de las astroesclereidas en el mesofilo. Adicionalmente, se puede señalar que la morfología y anatomía de estas hojas es muy parecida a la que exhiben las de angiospermas dicotiledóneas, particularmente aquellas con rasgos escleromorfos. Esto último es indicativo del grado de avance de esta familia dentro de las Gnetophyta.
BIBLIOGRAFÍA
1. Duthie, A.V. 1912. Anatomy of Gnetum africanum. Ann. Bot. 12: 593-602. [ Links ]
2. Gifford, E. & A. Foster. 1988. Morphology and evolution of vascular plants. Freeman and Company, New York. [ Links ]
3. Hickey, L. 1973. Classification of the architecture of dicotyledonous leaves. Amer. J. Bot. 60: 17-33. [ Links ]
4. Judd,W., Ch. Campbell, E. Kellogg & P. Stevens. 1999. Plant Systematics a phylogenetic approach. Sinauer Associates, Inc. Publisher. Sunderland, Massachusetts. [ Links ]
5. Kausik, S.B. 1974. Ontogeny of the stomata in Gnetum ula Brongn. Bot. J. Linn. Soc. 68: 143-151. [ Links ]
6. Maheswari, P. & V. Vasil. 1961. The stomata of Gnetum. Ann. Bot. 25: 313-319. [ Links ]
7. OBrien, T. & M. McCully. 1981. The study of plant structure. Principles and methods. Termarcarphi PTY., Melbourne,Australia. [ Links ]
8. Patel, J.A. 1979. A new morphological classification of stomatal complexes. Phytomorphology 29: 218-229. [ Links ]
9. Peña, E. & H. Saralegui. 1982. Técnicas de anatomía vegetal. Universidad de la Habana, Cuba. [ Links ]
10. Price, R. 1996. Systematics of the Gnetales: a review of morphological and molecular evidence. Int. J. Plant Sci. 157: S40-S49. [ Links ]
11. Rodin, R.J. 1966. Leaf structure and evolution in American species of Gnetum. Phytomorphology 16: 56-68. [ Links ]
12. Rodin, R.J. & G.S. Paliwal. 1970. Nodal anatomy of Gnetum ula. Phytomorphology 20: 103-111. [ Links ]
13. Stevenson, D. 1995. Gnetaceae. In: Flora of the Venezuelan Guayana (Steyermark, J.P., Berry, K. Yatskievych & B. Holst, eds.), pp. 573-576. Missouri Botanical Garden Press, St. Louis. [ Links ]
14. Stevenson, D. & T. Zanoni. 1991. Gnetaceae. In: Flora of the Guianas (Görtz-van, R. ed.), pp. 12-18. Koeltz Scientific Books. USA/Germany. [ Links ]
