











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkActa Botánica Venezuelica
versión impresa ISSN 0084-5906
Acta Bot. Venez. v.29 n.1 Caracas jun. 2006
BIOLOGÍA REPRODUCTIVA Y ASIGNACIÓN DE BIOMASA FLORAL EN SOLANUM GARDNERI SENDTH. (SOLANACEAE): UNA ESPECIE ANDROMONOICA
Reproductive biology and biomass floral allocation in Solanum gardneri Sendth. (Solanaceae): an andromonoecious species
Omaira HOKCHE D.1 y Nelson RAMÍREZ2
1Postgrado en Botánica, Instituto de Biología Experimental, Facultad de Ciencias, Universidad Central de Venezuela, Apartado 20513, Caracas, Venezuela. Dirección actual: Fundación Instituto Botánico de Venezuela, Herbario Nacional de Venezuela, Jardín Botánico de Caracas, Universidad Central de Venezuela, Apartado 2156, Caracas 1010-A, Venezuela. hokchede@rect.ucv.ve
2
Centro de Botánica Tropical, Instituto de Biología Experimental, Facultad de Ciencias, Universidad Central de Venezuela, Apartado 48312, Caracas 1041-A, Venezuela. nramirez@reacciun.veRESUMEN
Las características florales y la estrategia reproductiva de Solanum gardneri Sendth., un arbusto ampliamente distribuido en Venezuela, fueron analizadas considerando el dimorfismo floral asociado a la longitud del estilo con el objeto de establecer si esta condición floral determina un sistema sexual andromonoico. Las flores ofrecen polen como única recompensa, presentan una antesis diurna y los polinizadores fueron abejas pertenecientes a los géneros Augochloropsis, Centris, Exomalopsisy Xylocopa. El sistema de apareamiento es la xenogamia, característico de especies autoincompatibles, asociado con una alta relación Polen/Óvulo y una alta asignación de biomasa masculina y de atracción. La esterilidad femenina del morfo floral con estilo corto, unida a diferencias significativas entre los morfos florales en cuanto al número y tamaño de los óvulos, así como en el tamaño de los granos de polen, indica la condición funcionalmente andromonoica de S. gardneri. La hipótesis de la función masculina parece explicar la producción relativamente alta de frutos. El alto nivel de aborto de óvulos encontrado se ha asociado con una mejor utilización de los recursos a nivel postcigótico y al elevado número de semillas por fruto, lo que probablemente esté relacionado con la estrategia de diseminación, siendo la ornitocoria el mecanismo de dispersión encontrado.
Palabras clave: Andromonoecia, Asignación de biomasa floral, Autoincompatibilidad, Biología reproductiva,Dimorfismo floral, Polinización, Solanum,Venezuela
ABSTRACT
The breeding system and floral characteristics of Solanum gardneri Sendth., a woody shrub widely distributed in Venezuela, were investigated considering the floral dimorphism associated with the pistil length in order to know if this floral condition determines an andromonoic sexual system. Anthesis is diurnal; flowers offering only pollen as reward, and the pollinators are bees, belonging to the genera Augochloropsis, Centris, Exomalopsis and Xylocopa. S. gardneri has a xenogamous mating system, of a self-incompatible species, related to a high P/O ratio, associated with a high biomass allocation to male and attraction functions. Female sterility in short-style flower and significant differences between floral morphs in number and ovule size, and pollen size indicate the andromonocious condition of S. gardneri. The allocation invested in the male function and in the attraction structures promote outcrossing. The male function explains the relatively high fruit-set. The high ovule abortion level found has been associated to a better postcigotic resource allocation level and the higher seed number per fruit is probably related to the fruit dispersal strategy,which is ornitochory.
Key words: Andromonoecy, Floral biomass allocation, Floral dimorphism, Pollination, Reproductive biology, Self-incompatibility, Solanum,Venezuela
INTRODUCCIÓN
La andromonoecia, presencia de flores masculinas (estaminadas) y bisexuales (hermafroditas) en un mismo individuo, es un sistema sexual poco común, presente en aproximadamente 2% de las angiospermas (Yamplosky & Yamplosky 1922) y común en especies polinizadas por animales y por el viento (Bawa & Beach 1981; Bertin 1982; Primack & Lloyd 1980). Una de las hipótesis más aceptadas para explicar la evolución de la andromonoecia es aquella que señala que se trata de un mecanismo para maximizar los recursos asignados a las funciones reproductivas (Diggle 1993). De aquí, se ha reportado que las flores estaminadas requieren menor asignación de recursos que las flores perfectas (Schlessman et al. 2004) y se ha postulado que las plantas hermafroditas, que frecuentemente se autofertilizan, invierten menos en estructuras florales para atraer polinizadores que las especies que regularmente se entrecruzan (Charlesworth & Charlesworth 1987; Lloyd 1987). De acuerdo a los modelos de asignación de recursos, la andromonoecia se presenta en plantas donde el número óptimo de flores funcionalmente estaminadas es mayor que el número de flores que puede producir frutos y el costo de madurar un fruto es alto.
Varias hipótesis se han formulado para explicar el exceso de flores en plantas hermafroditas: limitación de polen, atracción de polinizadores, predictibilidad del ambiente, función masculina de las flores, entre otras (Sutherland 1986a, b, 1987; Sutherland & Delph 1984). En el contexto de la andromonoecia parece imperar la hipótesis de la función masculina, la cual sostiene que el exceso de producción de flores contribuirá incrementando la aptitud masculina por medio de la donación de polen, por lo tanto la asignación de recursos para la producción de flores masculinas y femeninas no sería equivalente (Charlesworth & Morgan 1991).
La mayoría de las especies del género Solanum tienen flores hermafroditas; no obstante, se puede encontrar gran diversidad de formas sexuales que incluyen especies andromonoicas, androdioicas y dioicas (Symon 1970, 1979; Anderson 1979; Coleman & Coleman 1982; Anderson & Symon 1989; Forni-Martins et al. 1998), las cuales pueden ser polinizadas por abejas vibradoras (Buchmann 1983; Bezerra & Machado 2003). Las plantas de Solanum gardneri Sendth. Producen flores con diferentes longitudes de estilo, lo que permite distinguir entre flores con estilo largo que sobrepasa las anteras y flores con estilo corto nunca más allá de las mismas. La presencia de diferentes tipos florales es común en otras especies del género (Coleman & Coleman 1982; Forni-Martins et al. 1998; Bezerra & Machado 2003) y podría ser considerada dentro del contexto de la andromonoecia, si las flores con estilo corto se comportaran como flores funcionalmente masculinas.
Debido a la incidencia de andromonoecia en el género Solanum y a la presencia de dimorfismo floral en Solanum gardneri, se planteó estudiar la biología reproductiva considerando la capacidad de producción de frutos en cruces inter e intra-morfos, así como la asignación de biomasa a las estructuras reproductivas, mecanismos de polinización y las diferencias morfológicas asociadas a las flores.
MATERIALES Y MÉTODOS
Solanum gardneri es un arbusto pequeño, espinoso, de 0,5 a 1,5 m de alto, presente desde México hasta Brasil, incluyendo las Antillas, y está ampliamente distribuido en Venezuela, donde crece en bosques secundarios, en formaciones xerofíticas o hábitat perturbado (Benítez 1981).
Área de estudio
Se muestrearon más de 15 individuos de Solanum gardneri distribuidos a lo largo de un sendero en el Arboretum de la Escuela de Biología, Universidad Central de Venezuela, ubicado en Colinas de Bello Monte, al SO del valle de Caracas, estado Miranda,Venezuela (10°30 Lat. N, 66°53 Long. O), a una altitud de 1.050–1.100 m snm. Esta reserva corresponde a un Bosque Seco Premontano (Ewel et al. 1976), con precipitación media anual que varía entre 550 y 1.100 mm, y temperatura promedio entre 18° y 24°C. La vegetación es alterada, secundaria, compuesta por dos estratos: uno superior dominado por árboles entre 8 y 18 m de altura, y uno inferior compuesto por hierbas y arbustos, mezclados con una regeneración abundante de los árboles del estrato superior.
Morfología floral
El color de las diferentes partes florales se estableció con material fresco utilizando el atlas de colores de Küppers (1979). Se determinó la orientación de las flores con relación al sustrato. El proceso de antesis (apertura de la flor) se estimó en 20 flores de diferentes individuos, cuantificando el número de flores marcadas que abre cada media hora. La longevidad floral se estimó por observaciones periódicas en 20 flores desde la antesis hasta el cierre de las flores.
Se estableció la presencia de dos tipos florales de acuerdo a la longitud del estilo, y se evaluó la morfología separadamente para cada tipo floral (Leppik 1977). Se cuantificaron las dimensiones florales (largo y diámetro de la flor así como de las anteras y del estilo, y la distancia anteras-estigma) para un total de 20 flores provenientes de un mínimo de 10 individuos, utilizando material fresco o preservado en alcohol al 70%. Se caracterizaron la forma y la ornamentación del grano de polen por observación directa bajo el microscopio óptico en 25 flores en antesis o yemas bien desarrolladas. De igual manera, el tamaño de los granos de polen y de los óvulos se evaluó en 20 mediciones usando un micrómetro. Posteriormente, se estimó el área del grano de polen basado en el diámetro del mismo (considerando el área de una esfera: A= .R2, donde A = área, R = radio).
El área estigmática se estimó indirectamente en cinco estigmas provenientes de flores de diferentes individuos, haciendo esquemas en cámara clara. Estos esquemas se pesaron y se dividieron entre el aumento utilizado, el peso de cada esquema se dividió entre el peso correspondiente a 1 cm2 para obtener el área estigmática real (cm2). Esto se realizó para cada morfo floral. El número de granos de polen que teóricamente puede depositarse sobre el estigma se calculó dividiendo el área estigmática estimada indirectamente, entre el área ocupada por el grano de polen sobre la superficie del estigma.
La carga de polen sobre los estigmas se determinó en 10 flores (de cada morfo floral) bajo condiciones naturales, los estigmas fueron colectados seis horas después de la antesis y teñidos con cotton blue en lactofenol. La capacidad de germinación del polen se estimó contando bajo el microscopio óptico los granos provenientes de anteras de diferentes flores en una solución de sacarosa al 20% por una semana. El número de granos de polen germinado fue cuantificado en 3 réplicas.
Estrategia reproductiva
Para determinar el sistema reproductivo se realizaron cuatro pruebas diferentes en cinco individuos, siguiendo el método descrito por Ruiz-Zapata & Arroyo (1978) y Sobrevila & Arroyo (1982): 1. La autofertilización se detectó en flores recién abiertas, aisladas previamente en estado de yemas, y luego polinizadas manualmente. Estas flores fueron nuevamente aisladas para evitar el contacto con polinizadores hasta la formación de frutos. 2. La fertilización cruzada se realizó colocando polen proveniente de diferentes individuos en estigmas receptivos de flores previamente emasculadas y aisladas. 3. El nivel de autogamia se estimó en yemas aisladas verificando posteriormente la formación de frutos. 4. La agamospermia se examinó emasculando yemas florales y manteniéndolas aisladas hasta la formación de frutos. Para cada tipo de prueba aplicada se cuantificó el número de frutos y semillas producido, y se verificó la presencia del embrión en las semillas. Además, para determinar la fertilidad femenina de cada morfo floral se realizaron pruebas de entrecruzamiento entre los dos tipos florales.
Biología de polinización
La actividad de los polinizadores se registró por observaciones directas en el campo en el momento de la antesis, en horas del mediodía y a finales de la tarde, en ocho individuos. Los agentes visitantes fueron capturados, identificados, medidos y examinados con relación a la presencia de polen durante tres días de observación, 5 h por día, para un total de 15. El número de granos de polen sobre cada parte del cuerpo del visitante floral se cuantificó colocando cada porción en una solución de glicerina (1–2 ml) al 50%. Después de agitar con un vortex, se tomó una alícuota de la preparación (0,2–1,8 ml dependiendo del tamaño de la parte del visitante) y se contó el número de granos de polen utilizando un hemacitómetro. Este conteo se realizó en cinco réplicas para cada parte del cuerpo del visitante floral. Para cada agente visitante se determinó la longitud de la proboscis utilizando una solución de KOH al 10% en baño de calentamiento hasta ebullición, para lograr la extensión de la estructura.
Producción de óvulos, polen y relación P/O
El número de óvulos por flor se cuantificó en un microscopio estereoscópico. Se disectaron 100 yemas de flores con estilo corto y 100 yemas de flores con estilo largo provenientes de cinco individuos. El número de granos de polen por antera se determinó utilizando el método descrito por Lloyd (1965), que consiste en suspender cinco anteras de diferentes yemas florales en 0,1 ml de una solución de anilina en lactofenol y glicerina 3:1. Posteriormente, en tres preparaciones, se cuantificaron los granos de polen con un hemacitómetro en un microscopio óptico. El número de granos de polen por flor se estimó multiplicando el número de granos de polen por antera por el número promedio de anteras por flor. La relación Polen/Óvulo (P/O) se estableció dividiendo el número promedio de granos de polen por flor entre el número promedio de óvulos por flor.
Biomasa de flores, frutos y semillas
El peso seco de las flores y de las diferentes partes florales (cáliz, verticilo vegetativo de soporte de la flor; corola, verticilo vegetativo de atracción; androceo y gineceo, órganos sexuales) se cuantificó en flores en antesis, y en frutos y semillas bien desarrolladas. El peso seco floral por morfo, así como el de frutos y semillas, se determinaron en 20 unidades, las cuales fueron secadas a 40°C durante dos (en el caso de flores) y cuatro (en el caso de frutos y semillas) semanas, y luego pesados utilizando una balanza analítica. Posteriormente, se establecieron las siguientes relaciones: Biomasa Masculina (androceo)/Biomasa Femenina (gineceo), Biomasa Reproductiva (androceo + gineceo)/Biomasa Vegetativa (cáliz + corola), y Biomasa de Atracción (corola)/Biomasa de Soporte (cáliz), como parámetros que reflejan la asignación de recursos a las diferentes funciones florales, de acuerdo al sistema de apareamiento de la especie.
Producción de flores, frutos y semillas
El número de flores por inflorescencia y frutos por infrutescencia se determinó por conteos directos en 100 inflorescencias y 100 infrutescencias, respectivamente, en cinco individuos. Se discriminó el número de flores con estilo largo y flores con estilo corto por inflorescencia.
Se determinó el número de semillas por fruto en 50-100 frutos maduros, y se contó el número de semillas abortadas por fruto discriminando, del total de semillas por fruto, aquellas semillas mal formadas, de tamaño reducido, comprimidas o carentes de embrión (Ramírez 1992).
Relación de pesos y aborto de estructuras reproductivas
Para medir la eficiencia reproductiva en ambos tipos de flores se estimaron las variables: Peso Total de Semillas/Fruto, Relación Pericarpo/Semilla, Peso Fruto/Peso Flor, Número de Óvulos/Inflorescencia, de Semillas/Infrutescencia, de Semillas Viables/Inflorescencia, de Semillas Viables/Óvulos por Inflorescencia, %Semillas Abortadas, %Óvulos Abortados, %Flores-Frutos Abortados, %Semillas Producidas (Ramírez 1992; Ramírez & Berry 1993). Es importante señalar que el número de flores con estilo corto no se consideró para determinar el promedio de flores/inflorescencia, para no sobreestimar los niveles de aborto.
Biología de diseminación
Se caracterizó morfológicamente el fruto, considerando las dimensiones y el color, y se estudió el síndrome de dispersión de acuerdo a las características generales del fruto, tomando en cuenta la clasificación de van der Pijl (1972).
Depredación predispersión de frutos
A fin de establecer la presencia de depredación predispersión de frutos se colectaron y se aislaron 120 frutos maduros en bolsas de tela metálica por un período de dos meses. Posteriormente, se calculó el porcentaje de depredación discriminando entre el total de frutos, aquellos que presentaran daño en las semillas.
Análisis de los resultados
La estrategia reproductiva de la especie se determinó utilizando dos índices: 1) Índice de Autoincompatibilidad, dividiendo el número promedio de frutos y semillas producido por flor autofertilizada manualmente entre el número promedio de frutos y semillas producido por flor por medio de fertilización cruzada manual, respectivamente para frutos y semillas, y 2) Índice de Autogamia (o Autopolinización Automática), comparando el porcentaje de frutos y semillas producido por autofertilización automática y autofertilización controlada, respectivamente para frutos y semillas (Ruiz-Zapata & Arroyo 1978). Se utilizó una prueba de t de Student de contrastación de medias (Sokal & Rohlf 1981) para establecer diferencias en las características florales en y entre morfos florales.
RESULTADOS
Biología floral y relación Polen/Óvulo
Solanum gardneri presenta inflorescencias axilares tipo corimbo con flores axilares, pleomórficas, con una orientación de horizontal hacia abajo a vertical hacia abajo respecto al sustrato. Las flores poseen pétalos blancos, anteras amarillas con tecas paralelas y dehiscencia poricida. Las flores no producen néctar y ofrecen polen como única recompensa para los polinizadores. El polen es solitario, de forma esférica, con una superficie lisa. El gineceo es sincárpico, con estigma difuso y estilo terete. La antesis es diurna y ocurre entre las 06:00 y 07:30 a.m. Más del 70% de las flores analizadas abrieron entre las 06:00 y 06:30 a.m. y pueden permanecer abiertas de uno a dos días.
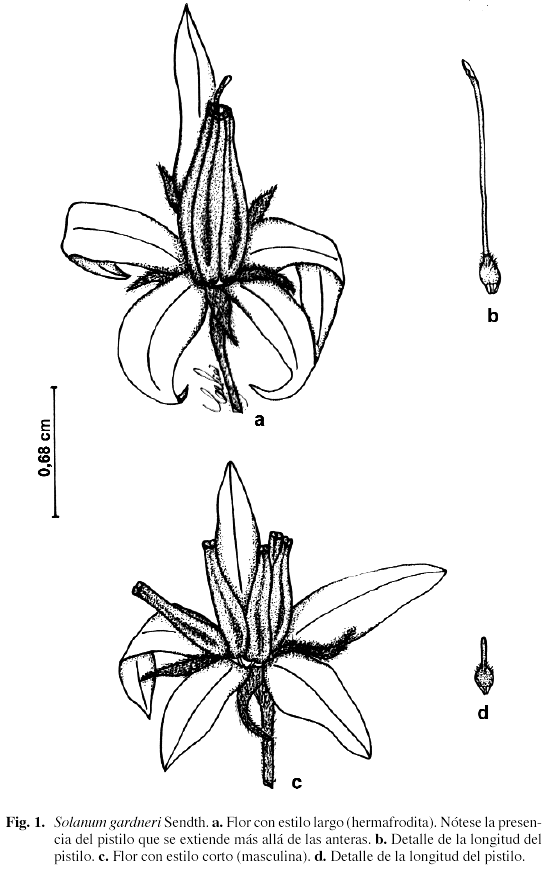
El análisis morfométrico floral muestra que S. gardneri presenta dimorfismo floral, encontrándose en una misma inflorescencia flores con estilo largo y flores con estilo corto, de aspecto similar (Fig. 1). Las dimensiones de los dos morfos florales se muestran en la Tabla 1. Ambos morfos difirieron significativamente en cuanto a la longitud de las flores, del estilo, de las anteras y a la distancia anteras - estigma (medida ésta por la diferencia en longitud entre ambos órganos, por tratarse de anteras con dehiscencia poricida) (Tabla 2, Anexo 1). También se encontraron diferencias significativas tanto en el número como en el tamaño de los óvulos producidos y en el tamaño de los granos de polen entre los morfos florales (Tabla 2, Anexo 1). El número de granos de polen producido por flor con estilo largo fue mayor que el producido por flor con estilo corto; sin embargo, esta diferencia no fue significativa (Tabla 2, Anexo 1). Las flores con estilo largo presentaron un área estigmática significativamente mayor que las flores con estilo corto (Tabla 2, Anexo 1). La relación P/O fue mayor en las flores con estilo corto asociada con un menor número de óvulos por flor (Tabla 2).


Estrategia reproductiva
Como resultado de las pruebas reproductivas se encontró que S. Gardneri es una especie xenógama obligada, que sólo produjo frutos y semillas por fertilización cruzada (Tabla 3). El índice de autoincompatibilidad genética (IAI = 0) muestra que se trata de una especie autoincompatible, de acuerdo al criterio de Ruiz-Zapata & Arroyo (1978) y Sobrevila & Arroyo (1982). En flores con estilo corto no hubo formación de frutos, lo cual muestra que estas flores son funcionalmente masculinas, y las flores con estilo largo son hermafroditas, de aquí que se pueda considerar a S. gardneri como una especie andromonoica.

Carga de polen
El número de granos de polen estimado teóricamente que puede cubrir el área estigmática fue 1.314,42 para flores hermafroditas, y 1.017,34 para flores masculinas. Para flores hermafroditas, el número de granos de polen obtenido experimentalmente sobre el estigma (X = 1.328,44, DS = 496,52) se correspondió con el número de granos de polen estimado teóricamente; esta variable no fue determinada para flores masculinas. El número promedio de granos de polen germinados fue de 152,33, lo que representa un porcentaje de germinación bajo (X = 19,33%, DS = 2,75, N = 457) comparado con el número promedio de granos de polen no germinados (X = 80,67%, DS = 2,75, N = 1.944). El número de granos de polen germinados relativo al número promedio de granos de polen depositados sobre el estigma (152,33 x 1.328,44/100 = 2.023,61) es considerablemente mayor que el número promedio de óvulos por flor (Tabla 2).
Biología de polinización
Las flores de S. gardneri fueron visitadas por abejas de las familias Anthophoridae, Apidae y Halictidae. Los polinizadores fueron discriminados de los visitantes florales de acuerdo al comportamiento y posición de las cargas de polen sobre el cuerpo. La máxima actividad de los polinizadores correspondió con las primeras horas después de la antesis, destacándose Exomalopsis sp., Xylocopa frontalis (Anthophoridae), Centris (Hemisiella) trigonoides, Centris inermis y Augochloropsis callichroa (Halictidae), principalmente aquellos con grandes cargas de polen sobre su cuerpo, específicamente en el tercer par de patas, parte ventral del tórax y abdomen (Tabla 4). Las abejas grandes (Xylocopa sp.) se posan sobre los poros de las anteras y la vibración de sus cuerpos ocasiona que el polen se deposite sobre las patas traseras y parte ventral principalmente, garantizando el contacto posterior del polen con el estigma. Dialictus sp. (Halictidae) y Trigona testaceicornis (Apidae) mostraron menor número de granos de polen sobre su cuerpo (Tabla 4). Cabe destacar que las abejas pequeñas remueven más fácilmente el polen de las flores hermafroditas y permanecen comparativamente mayor tiempo sobre la flor (observación no cuantificada). Centris (Hemisiella) trigonoides y C. inermis tuvieron un comportamiento similar a Xylocopa frontalis al visitar la flor, por lo que pueden ser considerados polinizadores efectivos aún cuando no se pudo cuantificar la carga de polen sobre su cuerpo. Dos especies de coleópteros se capturaron alimentándose de las partes florales y portando polen muy escaso y disperso en todo el cuerpo, por lo que fueron catalogados como visitantes florales.

Asignación de biomasa floral
Los promedios del peso seco de las flores y de las partes florales se señalan en la Tabla 5. El peso seco de las flores hermafroditas fue significativativamente mayor comparado con el de las flores masculinas (Anexo 1). Para ambos tipos florales la biomasa masculina (androceo) fue significativamente mayor que la biomasa femenina (gineceo); este mismo patrón se encontró al comparar la biomasa de atracción (corola) con la de soporte (cáliz), y la reproductiva (androceo + gineceo) con la vegetativa (cáliz + corola). Aun cuando entre morfos los valores de biomasa fueron significativamente mayores en las flores hermafroditas y la biomasa masculina fue similar, las relaciones de biomasa reproductiva/biomasa vegetativa, biomasa de atracción/biomasa de soporte y biomasa masculina/biomasa femenina fueron mayores en las flores masculinas (Tabla 5).

Eficiencia reproductiva
El número promedio de frutos/infrutescencia fue significativamente menor comparado con el número promedio de flores/inflorescencia, lo que indica que algunas flores abortan antes de producir frutos (Tabla 6). Este mismo patrón se encontró a nivel de semillas por fruto (incluidas las abortivas) comparado con el número de óvulos por flor para ambos morfos florales, es decir, hay una gran cantidad de óvulos que no llega a formar semillas. Esta tendencia se mantiene al comparar los altos valores de óvulos/inflorescencia con el bajo número de semillas/infrutescencia (Tabla 6). La proporción de aborto a nivel de flores-frutos inmaduros fue 34,5%, y 50,8% a nivel de óvulos; la proporción de semillas abortadas fue baja (2,1%); todo esto se traduce en una baja eficiencia reproductiva (31,6%).

El peso seco del fruto fue 0,050 (DS = 0,026). Se encontró una relación peso fruto/peso flor de 8,32. El peso seco de la semilla fue 0,0014 g. La relación peso semillas/fruto fue 0,031 y la de pericarpo/semilla fue 0,61.
Diseminación y depredación
El fruto de S. gardneri es una baya globosa con cáliz persistente y pubescente, de 1,00 cm largo y 0,89 cm diámetro, de color rojo cuando maduro, con superficie lisa y brillante. Estas características del fruto sugieren que el síndrome de dispersión es la ornitocoria. Asociados con las semillas de S. gardneri se encontraron insectos de la familia Anobiidae (Coleoptera), que se consideraron como depredadores predispersión produciendo daño casi total de las semillas afectadas y dejando sólo el borde intacto. El porcentaje de depredación predispersión encontrado fue 26,66%, observándose un sólo orificio de salida en las paredes del fruto lo que indica la presencia de un insecto por fruto.
DISCUSIÓN
Biología floral y polinización
Aunque las solanáceas presentan diversos modos de polinización (Bowers 1975; Voss et al. 1979; Coleman & Coleman 1982; Sullivan 1984; Fernández 1988; Cocucci 1991; Galetto 1991; Grases & Ramírez 1996; Vesprini & Galetto 2000; Stehmann & Semir 2001), las características florales de Solanum gardneri revelan adaptaciones a la polinización entomófila, específicamente la melitofilia: antesis diurna, flores pleomórficas, de color blanco y anteras amarillas, e inflorescencias más o menos densas, atributos comunes en especies polinizadas por abejas (Faegri & van der Pijl 1971; Proctor & Yeo 1973; Harborne 1977; Dafni & Neal 1997). Esto concuerda con lo reportado por Symon (1979) y Smith & Knapp (2002), quienes sostienen que las abejas son los principales agentes polinizadores en el género Solanum.
La extracción del polen de las anteras poricidas es realizada por la vibración de las abejas. Además, la orientación de las flores facilita la llegada de los polinizadores y la deposición del polen sobre ellos. Es importante señalar que las abejas pequeñas tienen mayor probabilidad de remover polen de las flores con estilo largo sin tocar el estigma, por lo que no serían los polinizadores más efectivos, a diferencia de las abejas grandes que aún cuando portan menor cantidad de polen, la probabilidad de contactar el estigma con la parte ventral del abdomen es mayor como resultado de la forma de posarse sobre la flor. No obstante, la carga de polen transportada ventralmente por los insectos grandes y pequeños excede la capacidad máxima del área estigmática de esta especie y garantiza la deposición de suficiente polen sobre el estigma. Sin embargo, las abejas colectan grandes cantidades de polen en sus patas, el cual pocas veces estará disponible para la polinización. Probablemente, la alta producción de polen en flores hermafroditas, además del polen producido en flores masculinas, promuevan cantidades adecuadas de polen depositadas ventralmente sobre el cuerpo del polinizador.
El incremento en la relación P/O asociado con la andromonoecia se ha interpretado como un sistema favorecido cuando los sistemas de polinización requieren grandes cantidades de polen (Heithaus et al. 1974). La alta relación P/O que resulta de la condición andromonoica puede ser una respuesta a un sistema de polinización entomófilo en el que parte del polen puede perderse con el movimiento de los polinizadores al visitar la flor, donde además, la planta ofrece polen como única recompensa a los agentes visitantes. Tomando en consideración el carácter andromonoico de la especie, esta relación será mucho mayor. Cabe destacar que los bajos niveles de granos de polen germinados son suficientes para fertilizar los óvulos. Además, el exceso de polen depositado sobre el estigma garantiza la fertilización.
El presente estudio reveló que Solanum gardneri posee un sistema de autoincompatibilidad. La dehiscencia poricida de las anteras, la ubicación de anteras y estigmas a diferentes niveles, y una alta relación P/O son características que promueven el entrecruzamiento. Esta especie es catalogada, de acuerdo con la clasificación de Cruden (1977), como xenógama obligada y concuerda con el hecho que las abejas son indispensables para la polinización.
La ausencia de producción de frutos en cruces intra- e inter-morfos florales en flores con estilo corto demuestra la condición masculina de este tipo de flores y, por lo tanto, S. gardneri puede ser considerada como una especie andromonoica en la cual la producción de flores masculinas es aproximadamente 50%. La condición masculina de las flores con estilo corto es reforzada por la presencia de estilos y estigmas reducidos, menor número y tamaño de óvulos, gineceo de menor tamaño y biomasa con relación a las flores con estilo largo. Una situación similar se reporta para varias especies de Solanum (Anderson & Symon 1989; Forni-Martins et al. 1998) y para Bauhinia ungulata (Caesalpiniaceae) (Ramírez et al. 1984), donde las flores con estilo corto tienen óvulos abortivos. Aparentemente, los cambios morfológicos observados en el gineceo de las flores con estilo corto en Solanum parecen representar algunas de las alteraciones que acompañan la evolución de la andromonoecia.
La evolución de la andromonoecia ha sido muy discutida (Heithaus et al. 1974; Ruiz-Zapata & Arroyo 1978; Bawa & Beach 1981; Bertin 1982; Coleman & Coleman 1982; Willson 1983; Solomon 1987) y, en general, se ha atribuido a presiones selectivas que promueven el entrecruzamiento e incrementan el despliegue floral (Lloyd 1979; Primack & Lloyd 1980). En S. gardneri las características florales podrían representar no sólo una especialización del papel sexual sino también una adaptación para promover el entrecruzamiento. Así, el papel reproductivo de las flores estaminadas en esta especie involucra mayor donación de polen y/o atracción de polinizadores. Schlessman et al. (2004) señalan que la función principal de las flores estaminadas de Pseudocymopterus montanus (Apiaceae) es la de donantes de polen.
Asignación de biomasa floral
La teoría de asignación sexual predice que el éxito femenino podría estar limitado por la disponibilidad de recursos para la producción de frutos y semillas, mientras que el éxito masculino podría estar limitado por la capacidad de atraer polinizadores (Charnov 1982). La menor asignación de recursos a óvulos o a partes vegetativas, podría considerarse una estrategia de producción de granos de polen más pequeños o un incremento en la cantidad de los mismos (Willson 1979). En S. gardneri la mayor asignación de recursos a la biomasa reproductiva, específicamente a la función masculina, en ambos tipos de flores, así como la biomasa asignada a las estructuras de atracción, indican un desbalance entre los verticilos florales y concuerdan con la condición alógama de esta especie autoincompatible.
Además, se ha señalado que la producción de mayor número de flores donantes de polen incrementa la capacidad de atracción de la planta a los polinizadores y, por tanto, refuerza la función masculina (Lloyd 1979; Bertin 1982; Solomon 1987). Los altos valores de la relación biomasa masculina/biomasa femenina evidencian una mayor asignación a la biomasa masculina comparada con la biomasa femenina a nivel de la flor. Las mayores relaciones de asignación de biomasa encontradas en las flores masculinas respecto a las flores hermafroditas, específicamente la de biomasa masculina/biomasa femenina, podrían estar asociadas con la especialización de las flores masculinas.
Se ha reportado que la andromonoecia es muy variable y puede estar determinada por variaciones en los recursos a nivel del individuo, por cambios ambientales, o estar genéticamente establecida (Diggle 1993, 1994). La ventaja de la andromonoecia puede reflejarse por el incremento del número de flores masculinas y/o por la reducción de la proporción de flores que producen óvulos (Bertin 1982). Solomon (1986) señala que la reducción en el número de flores que producen semillas puede conducir a ahorro de energía que puede ser invertida en el mantenimiento vegetativo, en la reproducción o en la producción de más flores estaminadas. En Solanum gardneri la producción de flores masculinas es una ventaja ya que el incremento en la cantidad de polen garantiza la producción de semillas. Considerando que las flores funcionalmente masculinas poseen estructuras femeninas que en algunos casos son casi vestigiales, puede suponerse que la andromonoecia en esta especie ha sido derivada de la condición hermafrodita, como ha sido sugerido por varios autores (Lewis 1942; Bertin 1982; Willson 1983).
Eficiencia reproductiva y niveles de aborto
Es muy frecuente que las plantas hermafroditas alógamas presenten una baja relación fruto-flor (Willson & Burley 1983; Sutherland & Delph 1984; Sutherland 1986a, b, 1987). La producción de frutos en S. gardneri es relativamente alta. Este resultado concuerda con la hipótesis de la función masculina (Sutherland 1986a), en la cual se produce un exceso de flores hermafroditas que funciona sólo como donante de polen; las flores con estilo corto cumplen esta función. Además, las flores funcionalmente masculinas contribuyen a la atracción de los polinizadores y a la donación de polen. La condición andromonoica contribuye a la aptitud masculina, al producir grandes cantidades de polen.
En Solanum gardneri la alta relación peso fruto/peso flor indica una baja similitud entre las estructuras reproductivas, donde el costo de producción del fruto es alto comparado con el costo de las flores masculinas. La alta inversión requerida para la formación del fruto carnoso (comparada con la de la flor) es producida a niveles relativamente elevados de óvulos y flores-frutos abortados. Desde el punto de vista energético, es conveniente que el aborto ocurra a nivel de flores y óvulos y no malgastar energía en la formación de frutos y semillas que posteriormente aborten (semillas abortadas). Probablemente, hay selección a favor de la calidad de la progenie que se forma a expensas de un mayor aborto a nivel de óvulos los cuales, aparentemente, determinan la baja eficiencia global de S. gardneri. El alto aborto de óvulos puede estar asociado con la condición autoincompatible de esta planta. En las especies autoincompatibles, el aborto de óvulos puede ser interpretado como la primera respuesta a la diversidad de granos de polen que llegan al estigma (Lloyd 1980).
El número de granos de polen transportado por los polinizadores y las cargas de polen depositadas sobre los estigmas en S. gardneri fueron mayores al número de óvulos. Probablemente, los altos niveles de óvulos abortados respondan a características diferentes a la limitación de polinizadores en S. gardneri y pudieran ser asociados con fallas en la polinización y en consecuencia en la fertilización, con aborto selectivo o limitaciones de recursos (Sutherland 1986a, b, 1987). Ramírez & Berry (1993) señalan que los mayores niveles de aborto de óvulos en especies con numerosas semillas están asociados a una mayor eficiencia en la utilización de los recursos maternos. La relación pericarpo/semilla aporta un estimado del costo de empaquetamiento de semillas por fruto, es decir, de la inversión de biomasa en pericarpo (inversión materna) relativa a la inversión de biomasa en progenie. En S. gardneri esta relación indica que existe una menor inversión de biomasa en la producción de semillas que en el pericarpo del fruto. Esto, asociado con un gran número de semillas por fruto probablemente esté relacionado con la estrategia de diseminación, es decir, en frutos carnosos tipo baya el éxito reproductivo se garantiza a través de un gran número de semillas por fruto (Ramírez & Berry 1993). Snow (1981) reporta que un incremento en el número de semillas se refleja en un aumento en la biomasa del fruto, y esto puede mejorar la atracción del fruto puesto que la producción de frutos ornitocóricos requiere de una gran inversión materna (frutos costosos) donde las unidades dispersantes deben tener ciertas características morfológicas (tamaño y forma) y bioquímicas (color, olor, composición nutricional de la recompensa) que permitan la atracción, el consumo, y consecuentemente, la dispersión de las semillas. En la especie estudiada parece establecerse un compromiso entre el costo de empaquetamiento de las semillas y la estrategia de diseminación; es decir, el fruto carnoso garantiza la protección, dispersión y diseminación de un elevado número de semillas por fruto.
En conclusión, el dimorfismo floral en S. gardneri corresponde a una condición andromonoica en la cual las flores funcionalmente masculinas presentan diferencias morfológicas notorias con relación a las flores hermafroditas. Solanum gardneri mostró valores intermedios en la producción de frutos, asociados a un sistema de autoincompatibilidad, lo cual responde a la condición andromonoica. La eficiencia global, semillas por óvulo por inflorescencia, fue baja, principalmente asociada a altos niveles de óvulos abortados.
AGRADECIMIENTOS
El primer autor agradece al Laboratorio de Biología Reproductiva, Instituto de Biología Experimental, UCV, por facilitar las instalaciones para el desarrollo de la parte experimental de este trabajo. Agradecemos a la Lic. Celia Gil por la elaboración de los esquemas botánicos. A los árbitros por sus comentarios y sugerencias. Este trabajo es el producto de la asignatura Biología Reproductiva de Angiospermas, Postgrado en Botánica, Universidad Central de Venezuela.
BIBLIOGRAFIA
1. Anderson, G.J. 1979. Dioecious Solanum species of hermaphroditic origin as an example of a broad evolutionary convergence. Nature, Lond. 282: 836-838. [ Links ]
2. Anderson, G.J. & D.E. Symon. 1989. Functional dioecy and andromonoecy in Solanum. Evolution 43: 204-219. [ Links ]
3. Bawa, K.S. & J.H. Beach. 1981. Evolution of sexual systems in flowering plants. Ann. Missouri Bot. Gard. 68: 254-274. [ Links ]
4. Benítez, C.E. 1981. Notas sobre especies de Solanum en Venezuela (II). Ernstia 7: 18–19. [ Links ]
5. Bertin, R.I. 1982. The evolution and maintenance of andromonoecy. Evol. Theor. 6: 25-32. [ Links ]
6. Bezerra, E.L. de & I.C. Machado. 2003. Biologia floral e sistema de polinização de Solanum stramonifolium Jacq. (Solanaceae) en remanescente de Mata Atlântica, Pernambuco. Acta Bot. Bras. 17(2): 247-257. [ Links ]
7. Bowers, K.A.W. 1975. The pollination ecology of Solanum rostratum (Solanaceae). Amer. J. Bot. 62: 633-638. [ Links ]
8. Buchmann, S.L. 1983. Buzz pollination in Angiosperms. In: Handbook of experimental pollination biology (Jones, C.E. & R.J. Little, eds.), pp. 73-113. New York. [ Links ]
9. Charlesworth, D. & B. Charlesworth. 1987. The effect of investment in attractive structures on allocation to male and female functions in plants. Evolution 41: 948-968. [ Links ]
10. Charlesworth, D. & M.T. Morgan. 1991. Allocation of resources to sex functions in flowering plants. Philos. Trans., Ser. B 332: 91-102. [ Links ]
11. Charnov, E.L. 1982. The theory of sex allocation. Princeton Univ. Press, Princeton, NJ. [ Links ]
12. Cocucci, A.A. 1991. Pollination biology of Nierembergia (Solanaceae). Pl. Syst. Evol. 174: 17-35. [ Links ]
13. Coleman, J.R. & M.A. Coleman. 1982. Reproductive biology of an andromonoecious Solanum (S. palinacanthum Dunal). Biotropica 14: 69-75. [ Links ]
14. Cruden, R.W. 1977. Pollen-Ovule ratios: A conservative indicator of breeding systems in flowering plants. Evolution 31: 32-46. [ Links ]
15. Dafni, A. & P.R. Neal. 1997. Sex and shape in floral advertisement: measurement, concepts and implications. Acta Hort. 437: 121-140. [ Links ]
16. Diggle, P.K. 1993. Developmental plasticity, genetic variation, and the evolution of andromonoecy in Solanum hirtum (Solanaceae). Amer. J. Bot. 80: 967-973. [ Links ]
17. Diggle, P.K. 1994. The expression of andromonoecy in Solanum hirtum (Solanaceae): Phenotypic plasticity and ontogenetic contingency. Amer. J. Bot. 81: 1354-1365. [ Links ]
18. Ewel, J.J., A. Madriz & J.A. Tosi Jr. 1976. Zonas de vida de Venezuela. Estado Sucre. Caracas (Fondo Nacional de Investigaciones Agropecuarias, Caracas). [ Links ]
19. Faegri, K. & L. van der Pijl. 1971. The principles of pollination ecology. Pergamon Press, New York. [ Links ]
20. Fernández, S.E. 1988. Biología floral de Solanum sessiliflorum Dun. var. sessiliflorum, na região de Manaus, AM. Acta Amazon. 18: 55-65. [ Links ]
21. Forni-Martins, E.R., M.C. Mendes Marques & M.R. Lemes. 1998. Biología floral e reprodução de Solanum paniculatum L. (Solanaceae) no estado de São Paulo, Brasil. Revista Brasil. Bot. 21: 117-124. [ Links ]
22. Galetto, L. 1991. Sobre el néctar y los nectarios de algunas especies de Nicotiana (Solanaceae). Kurtziana 21: 165-176. [ Links ]
23. Grases, C. & N. Ramírez. 1996. Biología reproductiva de especies ornitófilas de un bosque deciduo secundario tropical. Trabajo Especial de Grado. Escuela de Biología. Facultad de Ciencias. Universidad Central de Venezuela. Caracas. Venezuela. [ Links ]
24. Harborne, J.B. 1977. Introduction ecological biochemistry. Academic Press, London. [ Links ]
25. Heithaus, E.R., P.A. Opler & H.G. Baker. 1974. Bat activity and pollination of Bauhinia pauletia: Plant pollinator coevolution. Ecology 55: 412-419. [ Links ]
26. Küppers, H. 1979. Atlas de los colores. Editorial Blume, Barcelona. [ Links ]
27. Leppik, E.E. 1977. Floral evolution in relation to pollination ecology. Today & Tomorrow's Printers and Publishers, New Delhi. [ Links ]
28. Lewis, D. 1942. The evolution of sex in flowering plants. Biol. Rev. 17: 46-67. [ Links ]
29. Lloyd, D.G. 1965. Evolution of self-compatibility and racial differentiation in Leavenworthia (Cruciferae). Contr. Gray Herb. 195: 3-134. [ Links ]
30. Lloyd, D.G. 1979. Evolution towards dioecy in heterostyly plants. Pl. Syst. Evol. 131: 71-80. [ Links ]
31. Lloyd, D.G. 19870. Sexual strategies in plants. I. An hypothesis of serial adjustment of maternal investment during one reproductive session. New Phytol. 86: 69-79. [ Links ]
32. Lloyd, D.G. 1987. Allocation to pollen, seeds and pollination mechanisms in self-fertilizing plants. Funct. Evol. 1: 83-89. [ Links ]
33. Primack, R.B. & D.G. Lloyd. 1980. Andromonoecy in the New Zealand montane shrub Manuka, Leptospermum scoparium (Myrtaceae). Amer. J. Bot. 67: 361–368. [ Links ]
34. Proctor, M. & P. Yeo. 1973. The pollination of flowers. W. Collins Sons & Co. London. [ Links ]
35. Ramírez, N. 1992. Las características de las estructuras reproductivas, niveles de aborto y semillas producidas. Acta Ci. Venez. 43: 167-177. [ Links ]
36. Ramírez, N. & P. Berry. 1993. Producción y costo de frutos y semillas relacionados a los tipos morfológicos de frutos, unidad de dispersión y síndromes de dispersión. Ecotropicos 6: 43-60. [ Links ]
37. Ramírez, N., C. Sobrevila, N.X. de Enrech & T. Ruiz-Zapata. 1984. Floral biology and breeding system of Bauhinia ungulata L. (Leguminosae), a bat-pollinated tree in Venezuelan "Llanos". Amer. J. Bot. 71: 273-280. [ Links ]
38. Ruiz-Zapata, T. & M.T.K. Arroyo. 1978. Plant reproductive ecology of a secondary deciduous tropical forest in Venezuela. Biotropica 10: 221-230. [ Links ]
39. Schlessman, M.A., N. Underwood, T. Watkins, L.M. Graceffa & D. Cordray. 2004. Functions of staminate flowers in andromonoecious Pseudocymopterus montanus (Apiaceae, Apioideae). Pl. Spec. Biol. 19: 1-12. [ Links ]
40. Smith, S.D. & S. Knapp. 2002. The natural history of reproduction in Solanum and Lycianthes (Solanaceae) in a subtropical moist forest. Bull. Nat. Hist. Mus. Lond. (Bot.) 32: 125-136. [ Links ]
41. Snow, D.W. 1981. Tropical frugivorous birds and their food plants: A world survey. Biotropica 13: 1-14. [ Links ]
42. Sobrevila, C. & M.T.K. Arroyo. 1982. Breeding systems in a montane tropical cloud forest in Venezuela. Pl. Syst. Evol. 140: 19-37. [ Links ]
43. Sokal, R.R. & F.J. Rohlf. 1981. Biometry. 2nd ed. W.H. Freeman, New York. [ Links ]
44. Solomon, B.P. 1986. Sexual allocation and andromonoecy: Resource investment in male and hermaphrodite flowers of Solanum carolinense (Solanaceae). Amer. J. Bot. 73: 1215-1221. [ Links ]
45. Solomon, B.P. 1987. The role of male flowers in Solanum carolinense: pollen donors or pollinator attractors?. Evol. Trends Pl. 1: 89-93. [ Links ]
46. Stehmann, J.R. & J. Semir. 2001. Biologia reproductiva de Calibrachoa elegans (Miers) Stehmann & Semir (Solanaceae). Revista Brasil. Bot. 24: 43-49. [ Links ]
47. Sullivan, J.R. 1984. Pollination biology of Physalis viscosa var. cineracens (Solanaceae). Amer. J. Bot. 71: 815-820. [ Links ]
48. Sutherland, S. 1986a. Patterns of fruit-set: What controls fruit-flower ratios in plants?. Evolution 40: 117-128. [ Links ]
49. Sutherland, S. 1986b. Floral sex ratios, fruit-set, and resource allocation in plants. Ecology 67: 991-1001. [ Links ]
50. Sutherland, S. 1987. Why hemaphroditic plants produce many more flowers than fruits: Experimental tests with Agave mckelveyana. Evolution 4: 750-759. [ Links ]
51. Sutherland, S. & L.F. Delph. 1984. On the importance of male fitness in plants: Patterns of fruit-set. Ecology 65: 1093-1104. [ Links ]
52. Symon, D.E. 1970. Dioecious Solanums. Taxon 19: 909–910. [ Links ]
53. Symon, D.E. 1979. Sex forms in Solanum (Solanaceae) and the role of pollen collecting insects. In: The biology and taxonomy of Solanaceae (Hawes, J.G., R.N. Lester & A.D. Skelding, eds.), pp. 385–397. Academic Press, London, U.K. [ Links ]
54. van der Pijl, L. 1972. Principles of dispersal in higher plants. Second edition. Springer-Verlag, New York. [ Links ]
55. Vesprini, J.L. & L. Galetto. 2000. The reproductive biology of Jaborosa integrifolia (Solanaceae):Why its fruits are so rare?. Pl. Syst. Evol. 225: 15-28. [ Links ]
56. Voss, R., M. Turner, R. Inouye, M. Fisher & R. Cort. 1979. Floral biology of Markea neurantha Hemsley (Solanaceae), a bat-pollinated epiphyte. Amer. Midl. Naturalist 103: 262-268. [ Links ]
57. Willson, M.F. 1979. Sexual selection in plants. Amer. Naturalist 113: 777-790. [ Links ]
58. Willson, M.F. 1983. Plant Reproductive Ecology. John Wiley & Sons, New York. [ Links ]
59. Willson, M.F. & N. Burley. 1983. Mate choice in plants: Tactics, mechanisms, and consequences. Princeton Univ. Press, Princeton. [ Links ]
60. Yamplosky, C. & H. Yamplosky. 1922. Distribution of sex forms in the phanerogamic flora. Biblioth. Genet. 3: 1-62. [ Links ]

