











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkErnstia
versión impresa ISSN 0252-8274
Ernstia v.17 n.2 Caracas dic. 2007
ESTRUCTURA Y FLORÍSTICA DE UN BOSQUE RIBEREÑO DE MONTAÑA, PARQUE NACIONAL HENRI PITTIER, ESTADO ARAGUA
Alfonso Cardozo López1 y Dumas Conde M.2
1 Instituto de Botánica Agrícola, Facultad de Agronomía, Universidad Central de Venezuela. Venezuela. 2 Instituto Nacional de Tierras. Venezuela.
cardozoaj@hotmail.com, dconde@inti.gob.ve
COMPENDIO
El estudio de los bosques ribereños de montaña es una de las tareas prioritarias pendientes, ya que éstos actúan como corredores biológicos de especies vegetales que se encuentran aguas arriba y se hallan bajo el peligro latente de desaparición por las quemas periódicas a que anualmente se encuentran sometidos los bosques nublados en el Parque Nacional Henri Pittier (PNHP). Río del Medio es una quebrada del sector oriental del PNHP. A pesar de que sólo es una mínima superficie la que se considera (1/10 ha), no se tienen conocimientos sobre este tipo de formación vegetal en la selva nublada del PNHP. Se halló una especie que representa un nuevo registro para el PNHP, Metteniusa nucifera (Icacinaceae), la cual nunca ha sido registrada en el lado occidental del parque y sin embargo, fue la que obtuvo el segundo mayor valor de IVI (Índice de valor de Importancia) en el lugar estudiado. Igualmente se encontró que Chrysophyllum scalare (Sapotaceae) es una especie propia de selvas nubladas muy húmedas, la cual obtuvo el mayor valor de IVI del área estudiada. Esta especie también ha ocupado posición importante en bosques nublados estudiados en el lado occidental del parque (Pico Periquito). Las familias con mayor importancia florística en el lugar fueron las Moraceae y las Guttiferae, destacando Chrysochlamys clusiaefolia (Guttiferae) como una especie indicadora de selvas nubladas muy húmedas.
PALABRAS CLAVE: Selvas nubladas, bosques ribereños de montaña, Parque Nacional Henri Pittier.
ABSTRACT
The study of the riparian mountain forests is one of the pending high-priority tasks, since these act like biological corridors for vegetal species that are waters above and they are under the latent danger of disappearance by the periodic fires to which the cloud forests of the National Park Henri Pittier (PNHP) are annually submitted. Río del Medio is a canyon of the eastern sector of the PNHP, whose riparian forests are neither floristically not structurally known. Although it is only a small surface the one that is considered (1/10 ha), there is no knowledge about are not had on this type of vegetal formation in the cloudy forest of the PNHP. A species was found that represents a new register for the PNHP, Metteniusa nucifera (Icacinaceae), which never has been reported on the western side of the park; nevertheless, it was obtained the second greatest value of IVI (Index of value of Importance) in the area studied. Also Chrysophyllum scalare (Sapotaceae) a typical species of very humid cloud forests, obtained the greatest value of IVI of the area studied. This species has also occupied an important position in cloud forests studied on the western side of the park (Pico Periquito). The Moraceae and the Guttiferae are the families with greater floristic importance in the area studied. Chrysochlamys clusiaefolia standing out as an indicator species of very humid cloud forests.
KEY WORDS: Cloud forest, Montane Riparian Forests, Henri Pittier National Park.
INTRODUCCIÓN
Los bosques ribereños son franjas boscosas que crecen al lado de los ríos, quebradas, lagos y que están sujetos a períodos de aumento o disminución del volumen de agua.
Según Huber y Alarcón (1988) cuando el curso de agua fluye a lo largo de una zona no boscosa (sabana), el bosque desarrollado en sus orillas se denomina bosque de galería. Cuando crecen sobre topografía inclinada, se denominan bosques de galería de montaña o también bosques de vaguada.
Steyermark y Huber (1978) los llamaron bosques de galería residual por «tratarse de bosques relictos de la vegetación forestal original, ampliamente distribuida antes de la intervención humana». Según estos mismos autores, en dichos bosques «especies vegetales propias de las regiones más elevadas, pue- den encontrar condiciones favorables para su establecimiento a niveles altitudinales mucho más bajos».
Estos bosques han venido disminuyendo en los últimos años, la mayoría fragmentados debido a la ubicación de fincas, conucos y vegas en las márgenes de los ríos (Paredes-Bastidas 1997). El grado de amenaza de los bosques ribereños es grande en varias regiones del centro del país, donde se presentan asentamientos humanos densos a lo largo de los ríos.
La red hidrográfica del Parque Nacional Henri Pittier (PNHP) que vierte sus aguas en el Mar Caribe está constituida por varias quebradas y ríos, tales como San Miguel, Ocumare, Cata, Aroa, Choroní, Chuao, El Tigre, Apamate, Guarapito, Uricaro, entre otras; todos nacen en diferentes puntos de la selva nublada del PNHP, con su respectivo bosque ribereño en sus márgenes; dichas corrientes de agua constituyen la única fuente de abastecimiento de agua de los asentamientos urbanos del litoral aragüeño.
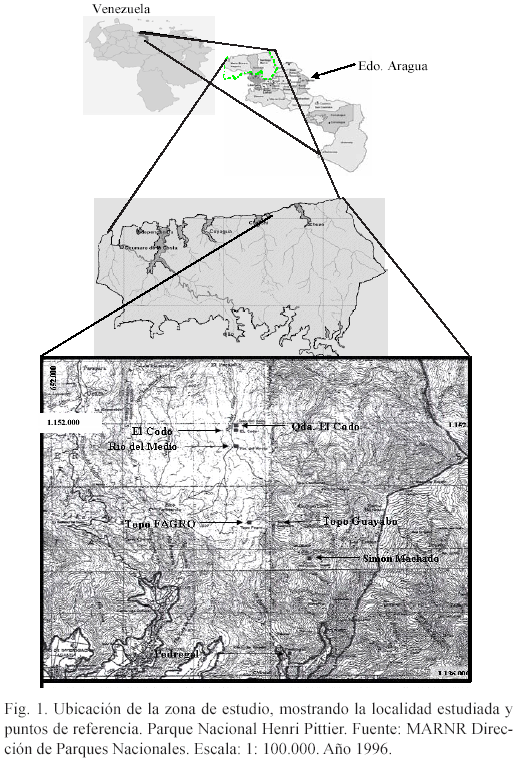
Río del Medio es uno de los cauces de agua que se cruza cuando se recorre el sendero Turmero-Chuao en la vertiente norte del lado oriental del parque. Realmente se trata de un tributario del verdadero Río del Medio (Fig. 1). Dicha corriente de agua se localiza en el centro-oriente del PNHP, el cual vierte sus aguas sobre el río Tamaira poco antes de formarse el río Chuao por la confluencia del río Tamaira con las quebradas Maestra y Sinamaica.
La vegetación que crece en los márgenes de Río del Medio en las zonas montañosas es prístina, se trata de bosques ribereños de montaña, con poca o ninguna intervención humana. Acerca de la vegetación que se localiza en esos lugares, se sabe muy poco. En esas áreas usualmente se presentan cambios debido a la dinámica natural del terreno, observándose deslaves, quemas por relámpagos, caída de árboles u otros fenómenos naturales que dejan grandes claros, razón por la cual se justifica el estudio y conocimiento de la vegetación que crece en las diferentes formaciones boscosas de sus márgenes como una forma de contribuir al conocimiento de la flora del parque antes de que ocurran cambios debido a la intervención antrópica, los cuales aunque puntuales, pueden alterar drásticamente el resto de la vegetación.
Los bosques ribereños de montaña actúan como corredores biológicos y no es raro encontrar especies propias de la selva nublada superior creciendo en los márgenes de ríos, en zonas mucho más bajas que donde usualmente se encuentran. La vegetación de estos bosques es una mezcla de especies, tanto del lugar que se estudia, como de especies de áreas aguas arriba. Así por ejemplo, Hirtella triandra (Chrysobalanaceae) propia de la selva nublada baja en Pico Periquito (± 1350 msnm) en el PNHP, es común en las márgenes de la Quebrada Los Araguatos (500 msnm) en la vertiente sur del mismo parque, así como en los márgenes del río Paya (± 400 msnm), vertiente sur del PNHP, pero en el lado Oriental del parque. Igualmente, Jiménez (1997) la registró en los márgenes de un afluente del río Guataparo (± 600 msnm), estado Carabobo. Otra especie, Prunus myrtifolia, es un árbol de la familia Rosaceae que ha sido visto y colectado en la cumbre de Pico Guacamaya (1850 msnm) del PNHP, la cual es frecuente en los márgenes de la quebrada Guacamaya aguas abajo (800 msnm) en la vertiente sur del mencionado parque.
Esta situación de especies de diferentes regiones altitudinales conviviendo juntas, se observa en todos los bosques ribereños de montaña, aún los cercanos a bosques deciduos. Bastardo (1994) observó en el bosque ribereño de la quebrada Los Araguatos, que la vegetación del lugar era una mezcla de vegetación xerófita con vegetación propia de áreas ubicadas más arriba del lugar estudiado; en el área se registraron varias especies de Acacia, tales como Acacia tortuosa, A. paniculata, y A. macracantha, indicadoras de zonas que sufren quemas, creciendo con Porcelia venezuelensis y Parinari pachyphila, propias de zonas húmedas y más elevadas altitudinalmente. La vegetación xerofítica busca refugio de las quemas periódicas y es común verla cercana o incluso en los márgenes del cauce y por otro lado, la corriente de agua actuando como un corredor biológico trae semillas y/o frutos; estas diásporas debido a la humedad relativa persistente todo el año germinan de manera que en estos ecosistemas pueden conseguirse asociadas especies normalmente ubicadas en pisos altitudinales muy diferentes.
El objetivo de este trabajo es el estudio de la composición florística y estructural de un sector de un bosque ribereño de montaña del PNHP ubicado muy cerca de las nacientes de un tributario de la Quebrada Río del Medio en la selva nublada baja en la vertiente norte del PNHP.
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
Se seleccionó una superficie ubicada en la orilla de una quebrada llamada Río del Medio (realmente se trata de un tributario de dicho río o el ramal oriental del mismo, el cual luego formará Río del Medio con otro tributario). Se encuentra a una altura de 1032 msnm. La topografía del terreno es plana a ligeramente inclinada y se localiza como a 12-15 m sobre el lecho de la mencionada quebrada. Las coordenadas UTM de un punto sobre un árbol del lugar fueron: 662.470 E y 1.150.585 N del huso 19 (las respectivas coordenadas geográficas son: Oeste 67º 32 35,17" y Norte 10º 24 4,25").
De acuerdo con Fernández (2000), Río del Medio nace en el Pico Cambural, no obstante, Río del Medio aparece denominado como tal en el mapa luego de la unión de dos tributarios o ramales, uno de ellos por el este (el cual nace en Pico Cambural) y el otro por el oeste (el cual se forma por la confluencia de tres quebradas pequeñas), que nace en Pico Palmarito (Fig.1). El ramal occidental es mucho más caudaloso que el ramal oriental y al unirse ambos constituyen Río del Medio (Fig. 1). El bosque estudiado en este trabajo se halla al margen del ramal oriental, un poco antes de su unión más abajo con el ramal occidental. Este ramal o tributario oriental, usualmente es denominado Río del Medio (Fig. 2) por las personas que hacen la travesía Turmero-Chuao por la montaña.

El acceso al área de estudio se logra como a cinco o seis horas de camino desde el puesto de guardaparques Simón Machado, el cual se ubica como a una hora (en vehículo de doble tracción) o cinco horas caminando desde el puesto de guarda-parques Pedregal. Este último lugar se encuentra como a 15- 20 minutos (en vehículo) de la población de Turmero (Fig. 1).
METODOLOGÍA
Se delimitaron 10 subparcelas de 10 x10 m2 se tomó nota del diámetro a la altura del pecho (DAP), altura, nombre de la especie y de la familia de cada uno de los individuos presentes en las 10 subparcelas cuyo DAP fuese mayor o igual a 2,5 cm (DAP ³ 2,5 cm) siguiendo la metodología del 1/10 ha (Gentry, 1982). Con estos valores se determinó la frecuencia, abundancia, área basal e Índice de valor de importancia (IVI) absolutos y relativos de todas las especies presentes en la superficie estudiada.
La frecuencia relativa (FR) se calculó multiplicando por 100 el número de veces en las que aparece cada especie en las 10 subparcelas, dividida entre la sumatoria de las frecuencias de todas las especies (FR = Nº subparcelas en que aparece cada especie x 100 / Ó de las frecuencias de todas las especies).
La abundancia relativa (AR) o densidad relativa de cada especie se calculó multiplicando por 100 el número de individuos de cada especie (Ai), dividida entre el número total de individuos de todas las especies (AR = Nº individuos de Ai x 100 / Nº total de individuos).
El área basal absoluta (ABA) de cada especie = å Áreas Basales de cada individuo de la especie; el área basal de cada individuo se obtiene mediante la fórmula S = pR2, en donde R = DAP/2. El área basal relativa (ABR) de cada especie se obtiene multiplicando por 100 la sumatoria Áreas Basales de todos los individuos de la especie / sumatoria Áreas Basales de todas las especies (Área Basal Relativa = å Áreas Basales de todos los individuos de la especie x 100/ Área Basal total).
En los casos de individuos multicaules (Palmae, Moraceae, Guttiferae), cada diámetro que se observó por encima de 2,5 cm se registró separadamente y para determinar las áreas basales de esas especies, se siguió el método propuesto por Franco-Rosselli et al. (1997), según el cual, el diámetro total (Dt) de las especies multicaules se obtiene mediante la fórmula Dt = (4At/p)1/2, en donde At es el área total de cada especie, la cual se calcula sumando las áreas individuales de todos los brotes de esa especie(Ari), es decir, At = SAri y el área de cada brote se calcula mediante la fórmula Ai = S(DAP)1/2
El IVI se determinó mediante la sumatoria de la frecuencia relativa, abundancia relativa y área basal relativa de cada especie.
Para este tipo de trabajo, tal como lo señala Kelly et al. (1994) se requiere estar familiarizado con la vegetación del lugar, ya que la mayoría de las veces el material colectado está en fase vegetativa.
La colección de los individuos se realizó usando la metodología común en estos casos, consistente en cortar ramas de cada uno mediante tijeras o gancho decopador. En el caso de los árboles se utilizaron los servicios de un escalador profesional. Las muestras fueron prensadas y empapadas en alcohol isopropílico para su conservación y posterior secado.
Aunque no se dispone de datos sobre temperatura, precipitación y humedad relativa, el lugar estudiado es muy húmedo. Si se compara con el bosque de los alrededores de la Estación Biológica, es mucho más húmedo que este último. El sólo hecho de crecer al lado de una corriente continua de agua (Fig. 2), ya lo ubica como un bosque muy húmedo. Tal como se señaló, en el sitio de trabajo son muy frecuentes especies indicadoras de ambientes muy húmedos, como Geonoma tenuis (palma enana, unifoliolada), Anthurium longissimum, Chrysochlamys clusiaefolia, Chrysochlamys membranacea, Besleria disgrega, Besleria clivorum, Chrysophyllum scalare, entre otras.
Muestras de suelo del sitio estudiado fueron analizadas en los laboratorios del Instituto de Edafología de la Facultad de Agronomía – UCV y las mismas resultaron ser características de un suelo muy ácido, con pH 4,3 y alto contenido de materia orgánica, de textura franco-arenosa (medianamente grueso), lo cual incide en una alta permeabilidad del mismo. El alto contenido de materia orgánica ayuda a una mayor retención de agua en el suelo. A pesar de ser ácido, los contenidos de Ca++ y K+ son medios y el de Mg++ es muy alto. El contenido de P es bajo, pero se compensa por la acción de las micorrizas que forman un sistema intrincado.
RESULTADOS
En la superficie estudiada se registraron 267 individuos pertenecientes a 72 especies y 31 familias de espermatofitas.
Las familias con mayor número de especies fueron: Moraceae (seis spp.), Guttiferae (cinco spp.), Palmae y Euphorbiaceae (cuatro spp. c/u). Hubo seis familias representadas por tres especies, las cuales fueron: Lauraceae, Leguminosae, Malpighiaceae, Sapindaceae, Solanaceae y Rubiaceae. Con dos especies figuran Apocynaceae, Araceae, Icacinaceae, Marcgraviaceae, Melastomataceae, Myrtaceae, y Sapotaceae y con una especie se tuvieron las familias Actinidaceae, Boraginaceae, Burseraceae, Chloranthaceae, Chrysobalanaceae, Lecythidaceae, Myristicaceae, Myrsinaceae y Olacaceae.
Dentro del material estudiado hubo nueve individuos cuyas familias no pudieron ser determinadas por falta de material reproductivo, pertenecientes a cinco morfo-especies.
De las 72 especies, hubo cuatro representadas por trepadoras, pertenecientes a las familias Araceae, Malpighiaceae, Marcgraviaceae y Sapindaceae, y sólo una especie con hábito epífito (Clusia multiflora).
En la Tabla I se aprecian las especies más abundantes entre las que destacan Metteniusa nucifera y Drypetes variabilis (33 y 17 individuos respectivamente); luego siguen Cordia allartii y Chrysochlamys membranacea (15 y 13 individuos c/u), Chrysochlamys clusiaefolia y Tovomita stigmatosa (12 y 10 individuos respectivamente), Cecropia peltata, Ficus sp. (A.Cardozo 3048) y Posoqueria coriacea (nueve individuos c/u).

Estas nueve especies, constituyen el 47,7% del total de individuos registrados en la superficie estudiada, tres de ellas de la familia Guttiferae (Clusiaceae). Hubo dos especies que presentaron multicaulía pero cada individuo multicaule se considera como uno solo; así, a pesar de que Bactris setulosa exhibió 28 tallos o brotes, los mismos correspondieron a cuatro individuos macolladores, mientras que Chrysochlamys clusiaefolia mostró 21 tallos que correspondieron a 12 individuos.
Tomando en cuenta las morfo-especies, hubo 29 especies representadas por un solo individuo, mientras que 10 especies se hallaron representadas por dos individuos.
Las especies más frecuentes (Tabla II) fueron: Metteniusa nucifera (presente en nueve subparcelas), Cordia allartii y Chrysochlamys clusiaefolia (presentes en ocho subparcelas), Tovomita stigmatosa, Drypetes variabilis, Chrysochlamys membranacea y Ficus sp. (presentes en siete subparcelas).

Las especies con mayor área basal fueron (Tabla III): Chrysophyllum scalare, Brosimum utile, Eschweilera beebei, Metteniusa nucifera, Dictyocaryum fuscum, Cecropia peltata y Richeria grandis. Las tres primeras especies, con sólo nueve individuos, representan el 57,14% del área basal total de los individuos muestreados en la superficie estudiada. Chrysophyllum scalare, con tres individuos (dos de ellos con 120 y 79 cm de DAP respectivamente) fue la especie con abrumadora mayoría en área basal, mientras que Brosimum utile, con dos individuos (43,8 y 91 cm de DAP respectivamente) fue la segunda especie con mayor área basal; esta especie exhibió uno de los individuos más altos (aproximadamente 50 m) y Eschweilera beebei, con cuatro individuos, de los cuales sólo uno con 90 cm de DAP, los restantes se hallaban entre el sotobosque y el segundo estrato. Otras especies arbóreas como Dictyocaryum fuscum, Cecropia peltata y Richeria grandis presentaron áreas basales relativas menores de 5%. Metteniusa nucifera y Chrysochlamys clusiaefolia, con áreas basales relativas ligeramente por encima del 5%, deben este valor a su abundancia, a diferencia de Dictyocaryum fuscum, que con sólo cinco individuos, presentó área basal relativa de 3,32%.

Las especies con mayores valores de IVI fueron (Tabla IV): Chrysophyllum scalare (36,25%), Metteniusa nucifera (22,08%), Brosimum utile (17,56%), Eschweilera beebei (15,93%), Drypetes variabilis (10,80%), Cordia allartii (10,30%), Chrysochlamys clusiaefolia (9,88%), Chrysochlamys membranacea (9,48%) y Tovomita stigmatosa (9,25%); luego sigue un grupo de cinco especies con valores de IVI entre 9% y 7% (Richeria grandis, Cecropia peltata, Posoqueria coriacea, Ficus sp.y Dictyocaryum fuscum).

Chrysophyllum scalare resultó la especie más importante de la comunidad estudiada debido a su alto valor de área basal, ya que sólo se observaron tres individuos; igual acontece con Brosimum utile y Eschweilera beebei, mientras que Metteniusa nucifera y Chrysochlamys clusiaefolia deben su ubicación como segunda y séptima especie respectivamente más importantes a la frecuencia, la abundancia y al área basal. Otras especies tales como Drypetes variabilis y Cordia allartii, deben su ubicación en el grupo de especies más importantes a la abundancia y a la frecuencia.
En el bosque estudiado, se establecieron cinco clases de altura y se tabuló el número de individuos que se encontraron en relación con su altura en cada clase (Fig. 3). El primero corresponde al estrato herbáceo y/o subarbustivo en el cual se localizan, además de las especies herbáceas propias del sotobosque, individuos juveniles y pequeños de especies arbustivas y arbóreas con DAP ³ 2,5 cm. Se ubicaron en este estrato todos los individuos cuya altura es inferior a 5 m. En este primer estrato se encuentra una cantidad (69) importante de individuos pertenecientes a 30 especies. Entre las especies arbustivas de poca altura destacan: Cordia allartii, Bactris setulosa, Cybianthus venezuelanus, Geonoma interrupta y Drypetes variablis. Entre los elementos herbáceos, subarbustivos y arbustivos presentes en el sotobosque se encontraron Geonoma tenuis (palma de hoja simple, la cual sólo se observa en ambientes muy húmedos), Anthurium longissimum (especie epífita, pero que también crece en el suelo por la caída de ramas), Besleria clivorum, Aphelandra micans, Besleria disgrega, Asplundia caput-medusae. Piper trigonum e individuos juveniles de especies arbóreas.

El segundo estrato corresponde a los individuos cuya altura varía entre 5 y 9,5 m; en la tercera clase se ubicarían los individuos cuya altura oscila entre 10 y 16,5 m; en la cuarta clase o dosel se ubican los individuos cuya altura varía entre 17 y 25 m de altura y los individuos sobresalientes o emergentes correspondientes al quinto estrato estarían por encima de los 25 m.
La mayoría de los individuos (135) se localizan en la segunda clase y una menor proporción se halla en la cuarta (15 individuos) y quinta (seis individuos) clases. Sólo se observaron seis individuos por encima de los 25 m de altura, que correspondieron a Chrysophyllum scalare y Eschweilera beebei (35 m c/u), Saurauia excelsa (33 m), Otoba parvifolia (26 m) y dos individuos de Brosimum utile (30 y 50 m respectivamente) (Fig. 3).
En el segundo estrato del perfil vertical (5-9,5 m), se aprecian como es lógico, individuos de las especies más abundantes, entre las que destacan: Metteniusa nucifera, (15 individuos), Cordia allartii, Chrysochlamys membranacea y Drypetes variabilis (11, 11 y ocho individuos respectivamente).
En el tercer piso, entre 10-16,5 m, el número de individuos es menor, (casi la mitad del número observado en el segundo), pertenecientes a las especies Metteniusa nucifera, Cecropia peltata, Richeria grandis, Eugenia sp. 2 y Drypetes variabilis.
En el dosel o cuarta clase (17-25 m) se observaron pocos individuos (15) pertenecientes a pocas especies (10), entre las que se hallan Dictyocaryum fuscum, con tres individuos y Chrysophyllum scalare con dos individuos. Entre los emergentes destacan las especies Brosimum utile (Moraceae), Chrysophyllum scalare (Sapotaceae), Saurauia excelsa (Actinidaceae), Eschweilera beebei (Lecythidaceae) y Otoba parvifolia (Myristicaceae), de los cuales Brosimum utile presentó un individuo con un poco más de 50 m de altura.
DISCUSIÓN
La especie más importante en la estructura del bosque (Chrysophyllum scalare) fue señalada por Huber (1986) como la especie típica, indicadora de selva nublada propiamente dicha; en esa oportunidad fue erróneamente determinada como Ecclinusa abreviata, posteriormente Pennington (1990) la publicó como especie nueva.
Esta especie es común en áreas húmedas o muy húmedas de la selva nublada baja del PNHP. Ha sido vista en Pico Periquito (1350 msnm), cumbre de Rancho Grande (1450 msnm) y Paso Portachuelo (1100 msnm), todos éstos, lugares húmedos a muy húmedos de la región occidental del PNHP y en varias localidades por el lado oriental en la vertiente Norte del PNHP (El Codo y Quebrada El Codo (1080 y 1100 msnm). Usualmente se asocia con especies arbóreas, sub-arbustivas, epífitas y herbáceas propias de ambientes húmedos, entre las cuales destacan: Anthurium longissimum (Araceae), Geonoma spinescens (Palmae), Besleria disgrega y Besleria clivorum, (Gesneriaceae), entre otras.
La especie más abundante, más frecuente y la segunda con mayor valor de IVI, Metteniusa nucifera, es una Icacinaceae que hasta ahora no había sido reportada del PNHP a pesar de ser conocida de Distrito Federal, Miranda, Carabobo y Falcón. Numerosas exploraciones realizadas en la zona occidental del PNHP no la habían reportado, sin embargo, resulta sorprendente que sea una de las especies más importante en uno de los bosques húmedos del sector oriental en la vertiente Norte. Esta especie es relativamente común en los bosques ribereños de montaña del lado oriental ya que ha sido vista en otra quebrada de menor cauce (Quebrada El Codo), y como ocurre con Río del Medio, mantiene siempre su curso de agua, dicha quebrada dista como una hora de camino del área estudiada. Probablemente se encuentre en las proximidades de algunas quebradas en la vertiente Norte por el lado occidental, pero hasta ahora no existe registro de la misma. Los ríos y quebradas en ésta última zona se localizan en áreas de difícil acceso, sobre terrenos escarpados y tal vez ésta sea la razón por la cual no ha sido registrada de esta área.
Hubo 11 especies cuyos valores de IVI estuvieron por encima de 8%, las cuales fueron en orden decreciente de valor de IVI las siguientes: Chrysophyllum scalare (36,25%), Metteniusa nucifera (22,08%), Brosimum utile (17,56%), Eschweilera beebei (15,93%), Drypetes variabilis (10,8%), Cordia allartii (10,3%), Chrysochlamys clusiaefolia (9,88%), Chrysochlamys membranacea (9,48%), Tovomita stigmatosa (9,25%), Richeria grandis (9,03%) y Cecropia peltata (8,78%). Son especies muy importantes en la parcela estudiada, representan casi la mitad de los individuos (47,7%) y la sumatoria de sus valores de IVI constituye un alto porcentaje del total.
Algunas son importantes en los niveles superiores de la estructura vertical del bosque (dosel o emergentes), las cuales usualmente están representadas por pocos individuos. Tres especies, Chrysophyllum scalare, Brosimum utile y Eschweilera beebei dominaban en el dosel o como emergentes y en conjunto presentaron nueve individuos. De los tres individuos de Chrysophyllum scalare, dos se encontraban en el nivel entre 17-25 m y el tercero como emergente (35 m). Los dos individuos de Brosimum utile fueron emergentes y de los cuatro individuos de Eschweilera beebei, sólo uno era emergente, otros dos estaban en el estrato 5-9,5 m y uno en el inferior, por debajo de los 5 m.
Otras especies son importantes en los niveles medio y/o bajo, es decir, en los niveles entre 10-16,5 m o entre 5-9,5 m tales como Metteniusa nucifera, Cordia allartii, Drypetes variablis, Chrysochlamys clusiaefolia, C. membranacea, y Tovomita stigmatosa, entre otras. En el caso de Metteniusa nucifera de los 33 individuos registrados, la mayoría se ubicaron en los estratos medio y bajo, quince (15) de los individuos de esta especie se localizaron en el nivel 5-9,5 m y once (11) de ellos se hallaban en el estrato 10-16,5 m; sólo un individuo se halló en el cuarto estrato (18 m) y seis por debajo de los 5 m. Desde el punto de vista de la distribución horizontal en la superficie estudiada, esta especie presentó la distribución más homogénea de todas, ya que se halló presente en 9 de las 10 subparcelas estudiadas y desde el punto de vista de su distribución vertical también fue la especie más homogéneamente distribuida debido a que estuvo presente en cuatro de los cinco niveles; y sólo ausente como emergente, pero éstas son características inherentes a la especie y probablemente sea la máxima altura que dicha especie alcance. En general, las especies de Icacinaceae en el PNHP son árboles pequeños a medianos.
Chrysochlamys clusiaefolia presentó un individuo con tres tallos en el cuarto nivel (22-24 m) y otro individuo con dos tallos en el tercer nivel (14-15 m), los restantes ocho individuos se ubicaron en niveles más bajos. Otras especies como Cordia allartii, Drypetes variabilis y Chrysochlamys membranacea tenían la mayoría de sus individuos en el segundo o tercer nivel y el resto por debajo de los 5 m. Igual sucede con Tovomita stigmatosa, Posoqueria coriacea y Richeria grandis, pero estas últimas especies estaban representadas en el dosel (cuarto nivel) por un individuo en cada caso.
Existe un grupo de especies que ocupan una posición no menos importante en la comunidad estudiada, ya que sus valores de IVI se hallan entre 7% - 8%. No son abundantes, pero tampoco son escasas, debido a que en general la mayoría estuvo representada por pocos individuos y sólo 15 tenían más de cinco individuos, las restantes 57 especies estuvieron representadas por 1-4 individuos. Estas especies son Dictyocaryum fuscum (7,34%), Eugenia sp. 2 (7,45%) y Ficus sp. (7,56%).
Algunos autores (Carlquist, 1974; Burger, 1981) citados en Gentry (1982) utilizan la relación especies/género como un indicador de la diversidad de una localidad. En lugares húmedos a muy húmedos estudiados por Gentry (1982) esta relación oscila entre 1,2-1,3 especies por género, mientras que en lugares secos esta relación oscila entre 1,5-1,8 especies por género. El cociente especies/ género obtenido en Río del Medio fue de 1,14, es decir, propio de lugares con alta diversidad, lo cual es un indicador más de que el bosque estudiado en Río del Medio presenta una alta diversidad. Tal como señala Gentry (1982), es de esperarse que a medida que aumenta la precipitación (aumenta la humedad), aumenta la diversidad ya que se van agregando nuevos géneros de plantas a la comunidad vegetal.
No existe un lugar del PNHP con el que se pueda comparar los resultados obtenidos en Río del Medio. Lo único conocido del parque es una comunidad ubicada en el lado occidental, en Pico Periquito a 1350 msnm. Como su nombre lo indica, la superficie estudiada se halla sobre un topo y laderas empinadas de dicha cumbre. La comunidad boscosa de Pico Periquito ha sido considerada por algunos investigadores como característica de selva nublada propiamente dicha (Huber 1986) o de selva nublada baja (Cardozo 1999). En Pico Periquito no se pudieron trazar parcelas de 10x10 m² de allí que no son comparables los dos lugares estudiados. Además Río del Medio se halla en un valle o vaguada entre cerros, mientras que Pico Periquito es una cumbre, ambos lugares son muy húmedos, con condiciones ambientales y climáticas diferentes. Sin embargo, sólo con fines comparativos entre dos lugares muy húmedos del PNHP, conviene señalar similitudes y diferencias entre dos comunidades boscosas del PNHP.
El número de familias, especies e individuos encontrados en Pico Periquito (PP) fue mayor que en Río del Medio, expresado en 38 familias, 90 especies y 440 individuos vs. 31 familias, 72 especies y 267 individuos. La menor riqueza florística en Río del Medio se debe a que el área estudiada en PP atraviesa varios pisos altitudinales a diferencia de Río del Medio. En Río del Medio las parcelas estudiadas se hallaban a la misma altura, mientras que en Pico Periquito debido a dificultades para establecer parcelas de 10x10 m2, se hicieron transectas de 50x2 m2, lo que trajo como consecuencia que hubo que trabajar en diferentes zonas altitudinales y por ende con mayor diversidad de nichos y ambientes distintos. Este efecto de beta diversidad produjo como resultado mayor riqueza florística en Pico Periquito. Otra consideración importante es que el área de trabajo realizado en Pico Periquito oscila entre 1300 - 1350 msnm y como se sabe (Gentry 1982), en los bosques tropicales la diversidad aumenta con la altura hasta los 1500 msnm, y Río del Medio se halla a una altura de 1032 msnm.
Las familia dominante en Pico Periquito fue Rubiaceae (diez spp.), seguida por Palmae y Guttiferae (siete spp. c/u), luego Lauraceae, Melastomataceae y Euphorbiaceae (cinco spp. c/u), mientras que en Río del Medio las familias dominantes fueron Moraceae y Guttiferae (seis y cinco spp. c/u), luego Palmae y Euphorbiaceae (cuatro spp. c/u).
Las especies más abundantes en Pico Periquito fueron Ladenbergia muzonensis y Psychotria costanensis (Rubiaceae), mientras que en Río del Medio fueron Metteniusa nucifera (Icacinaceae) y Drypetes variabilis (Euphorbiaceae). Las dos especies de Rubiaceae más abundantes en Pico Periquito son dos árboles pequeños o arbustos (2-5 m altura), mientras que las dos especies más abundantes en Río del Medio son dos árboles del estrato medio y del dosel.
Las especies con mayores valores de área basal también fueron diferentes en las dos localidades, en Pico Periquito fueron Eschweilera perumbonata, Chrysophyllum scalare, Aspidosperma fendleri, Micropholis crotonoides y Beilschmiedia pendula, y en Río del Medio fueron Chrysophyllum scalare, Metteniusa nucifera, Brosimum utile, Eschweilera beebei y Dictyocaryum fuscum.
Las especies con mayor IVI en Pico Periquito fueron las mismas que dominan en área basal, pero se incluye entre las cinco primeras a Ladenbergia muzonensis, mientras en Río del Medio además de las dominantes en área basal se incluyen Drypetes variabilis y Cordia allartii.
Ambas localidades sólo tienen en común a Chrysophyllum scalare entre las especies más importantes. Sólo coinciden en que una de las especies con mayor área basal e IVI en Pico Periquito fue la especie con mayor área basal e IVI en Río del Medio, lo cual permite sacar como conclusión que esta especie es característica de selvas húmedas o muy húmedas del Parque Nacional Henri Pittier.
Es importante destacar que Ladenbergia muzonensis (=Cinchona henleana) siendo una especie tan abundante y fácil de distinguir en el campo en Pico Periquito y alrededores, sea tan rara o ausente en Río del Medio. Esto mismo se aplica para Metteniusa nucifera, la cual en Río del Medio es abundante y frecuente (fácil de distinguir hasta vegetativamente), pero nunca ha sido vista en Pico Periquito y sus alrededores. Esto corrobora la teoría expuesta en Tanner et al. (1994) de que cada bosque montano tropical es sustancialmente diferente de cualquier otro, aún si éste se halla próximo. Estas dos parcelas no están tan próximas entre sí, pero si se considera el parque en el entorno de Venezuela, pertenecen a la misma formación denominada bosque submontano ombrófilo (Huber 1998), dos asociaciones de una misma formación, la cual si se observa en el mapa se ve pequeña.
Sería muy interesante comparar florística y estructuralmente la selva nublada baja estudiada en Pico Periquito con la que se encuentra en el Picacho Rancho Grande, los cuales se hallan uno frente al otro en el lado occidental del parque, pero no existe un estudio de este tipo sobre la cumbre de Rancho Grande. En el futuro sería igualmente interesante comparar la comunidad boscosa de Río del Medio con una comunidad situada a la misma altura y con condiciones climáticoedáficas similares.
REFERENCIAS BIBLIOGRÁFICAS
1. Bastardo, X. 1994. Inventario florístico de un bosque ribereño en la quebrada Los Araguatos, de la cuenca del río Güey, Parque Nacional Henri Pittier. Trabajo de Grado. Facultad de Agronomía. Universidad Central de Venezuela. 268 p. [ Links ]
2. Cardozo, A. 1999. Comparación florística y estructural entre la selva nublada baja y la selva nublada superior, Parque Nacional Henri Pittier, estados Aragua y Carabobo. Venezuela. Tesis Doctoral. Facultad de Ciencias. Universidad Central de Venezuela. 366 p. [ Links ]
3. Franco-Rosselli, P., J. Betancour y J. Fernández-Alonso. 1997. Diversidad florística en dos bosques subandinos del sur de Colombia. Caldasia 19: 205-234. [ Links ]
4. Gentry, A. 1982. Patterns of neotropical plant diversity. Evol. Biol. 15:1-84. [ Links ]
5.Huber, O. (Ed.). 1986. Las selvas nubladas de Rancho Grande: Observaciones sobre su fisonomía, estructura y fenología. La selva nublada de Rancho Grande, Parque Nacional Henri Pittier. Fondo Editorial Acta Científica Venezolana. Caracas. 288 p. [ Links ]
6. Huber, O. y C. Alarcón. 1988. Mapa de vegetación de Venezuela. Escala 1:2000000. M.A.R.N. Caracas. Venezuela. [ Links ]
7. Jiménez, E. 1997. Inventario florístico de un bosque de galería y sus adyacencias en un afluente del río Guataparo, Haciendas Guataparo, Valencia, estado Carabobo. Trabajo de Grado. Facultad de Agronomía. Universidad Central de Venezuela. 172 p. [ Links ]
8. Kelly, D., E. Tanner, E. Niclughadha and V. Kapos. 1994. Floristic and biogeography of a rain forest in the Venezuelan Andes. Journal of Biogeography 21: 421-440. [ Links ]
9. Paredes-Bastidas, A. 1997. Importancia de los bosques ribereños en el pie de monte andino y los llanos altos occidentales de Venezuela. Biollania 13: 47-66. [ Links ]
10. Steyermark, J. y O. Huber 1978. Flora del Ávila. Sociedad Venezolana de Ciencias Naturales, Vollmer Foundation y Ministerio del Ambiente y de los Recursos Naturales Renovables. Caracas. 971 p. [ Links ]
Anexo





