










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Facultad de Ciencias Veterinarias
versión impresa ISSN 0258-6576
Rev. Fac. Cienc. Vet. v.53 n.1 Maracay jun. 2012
Caracterización morfológica e histoquímica del hígado de la baba (Caiman Crocodilus Crocodilus)
Sonia Alvarado-Rico*,1, Gisela García*, Raquel Céspedes**, Martha Casañas*** y Albert Rodríguez****
*Cátedra de Histología. Facultad de Ciencias Veterinarias.Universidad Central de Venezuela
**Cátedra de Anatomía Facultad de Ciencias Veterinarias.Universidad Central de Venezuela
***Postgrado de la Facultad de Ciencias Veterinarias.Universidad Central de Venezuela
****Pregrado de la Facultad de Ciencias Veterinarias. Universidad Central de Venezuela. Apartado 4563, Maracay, 2101A, estado Aragua, Venezuela. Correo-E:alvaradorico8@gmail.com
Resumen
La morfología, organización y los componentes intracitoplasmáticos del hepatocito de la baba (Caiman crocodilus crocodilus) son aspectos que se han estudiado parcialmente hasta el momento. Con el fin de generar información sobre la morfología microscópica del hígado de esta subespecie, se realizó un estudio descriptivo de las características morfológicas e histoquímicas del hígado de la baba, lo que permitirá la comprensión de algunas funciones hepáticas en estos ejemplares. De esta manera, se podrán realizar en el futuro, investigaciones utilizando a dicho órgano como modelo experimental para el estudio de las rutas metabólicas de los metabolitos procesados a este nivel. Se tomaron ocho muestras de hígado, se fijaron en formol al 10% y se procesaron por los métodos histológicos de coloraciones de rutina (Hematoxilina y Eosina). Asimismo, se utilizaron las coloraciones especiales de Tricrómico de Gallegos, el método de Wilder y el aceite rojo O. Los hallazgos del presente trabajo revelaron la presencia de un hígado bilobulado, ubicado en la cavidad celómica caudal, con los lóbulos derecho e izquierdo unidos por un istmo. El órgano se encuentra rodeado por una delgada cápsula de tejido conectivo, su parénquima se dispone en cordones tortuosos con un patrón acinar, con presencia de células pigmentadas compatibles con melanóforos, dispersas en el tejido hepático, y con gotas lipídicas intracitoplasmáticas de diversos tamaños, que en algunos casos ocupaban casi la totalidad del citoplasma de los hepatocitos.
(Palabras clave: Caiman (crocodilus crocodilus), hígado, anatomía animal, histología animal, Aragua)
Morphological and Histochemical Characterization of the Liver of the Spectacled Cayman (Caiman crocodilus crocodilus)
Abstract
The morphology, organization and intracytoplasmic components of the liver of the spectacled cayman (Caiman crocodilus crocodilus) are aspects that have been partially studied so far. A descriptive study of the morphological and histochemical characteristics of the liver of this subspecies was performed. This investigation would enable researchers to understand the liver functions in these animals, thus allowing future research using this organ as an experimental model. Eight liver samples were fixed in 10% v/v formalin and processed by conventional histology using routine hematoxylin and eosin staining. Additionaly, special stains, such as the Trichrome of Gallegos method and the red oil O and Wilder, were also used. The findings revealed the presence of a bilobed liver located in the midline of the caudal coelomic cavity, surrounded by a thin capsule of connective tissue; the parenchyma showed the layout of the classic hepatic lobule, the hepatocytes were arranged in tortuous cords and an acinar pattern. The presence of pigment-laden cells compatible with melanophores scattered in liver tissue was evidenced. Intracytoplasmic lipid droplets of various sizes occupying almost the entire cytoplasm of the hepatocytes were observed.
(Key words: Caiman (crocodilus crocodilus), liver, animal morphology, animal histology, Aragua)
Recibido: 28/11/11 - Aprobado: 13/07/12
Introducción
La baba, pertenece al orden Crocodylia, el cual está compuesto por 22 especies y dividido en tres grupos básicos considerados como las subfamilias: Crocodylinae y Gavialinae, y la subfamilia Alligatorinae, donde se encuentra la subespecie Caiman crocodilus crocodilus (Ross y Magnusson, 1989). Estos reptiles forman un grupo de animales muy antiguos cuyos fósiles se remontan a unos 200 millones de años (Britton, 2002).
En Venezuela, existen al menos cinco especies de crocodilidus, entre las cuales se encuentran el caimán del Orinoco (Crocodilus intermedius) y el caimán de la Costa (Crocodilus acutus), el babo morichalero (Paleosuchus palpehosus), caimanes enanos (Paleosuchus trigonatus) y la baba (Caiman crocodilus crocodilus; Brazaitis, 1973), subespecie objeto de este estudio. La baba se ubica en nuestro país principalmente en las zonas llaneras inundables de los estados Apure, Barinas, Cojedes, Guárico y Portuguesa, y en la Cuenca y el Delta del Orinoco (Velasco y Ayarzagüena, 1992).
Las especies Crocodilus intermedius, Crocodilus acutus y la sub especie Caiman crocodilus crocodilus son consideradas como las de mayor potencial biológico y económico tanto en la industria peletera, como para el consumo de su carne, huevos, hemipene, huesos, grasas y vísceras. Con respecto al Caiman crocodilus crocodilus, su explotación y cría en cautiverio se han practicado desde hace dos décadas (Velasco et al., 2003). Con el objetivo de consumir su carne como fuente proteica animal, en la actualidad su captura se hace en forma indiscriminada y sin ningún control; además, estos animales son considerados productos de exportación, sin tomar en cuenta los aspectos sanitarios básicos. Por lo antes expuesto, es importante generar información sobre los aspectos nutricionales, conductuales, anatomofisiológicos, metabólicos y reproductivos de la baba, a fin de mejorar el manejo sanitario a nivel de zoocriaderos y de solventar las posibles patologías que pudieran presentarse. El manejo sanitario adecuado permitirá su explotación racional controlada y garantizará la restitución de los ejemplares a su ambiente natural.
Se han realizado estudios relacionados con las características anatómicas, histológicas e histoquímicas del sistema digestivo de la baba (Rossini, 2003); características anatómicas e histológicas del aparato genital masculino del Caiman crocodilus crocodilus (Cabrera y García, 2004; Cabrera et al., 2007), estudios acerca de la presencia de mucinas en el aparato genital masculino de la baba (Cabrera y García, 2007); y descripción morfológica de las células sanguíneas del Caiman crocodilus crocodilus (Rossini y García, 2010).
Desde el punto de vista anatómico, el hígado es una glándula simple, que constituye del 2 al 5% del peso corporal en un individuo adulto (Banks, 1986). Este órgano recibe irrigación especial de aporte y funcional. La irrigación de aporte proviene del tronco celíaco, la primera rama visceral colateral de la arteria aorta abdominal, mediante la arteria hepática, la cual irriga al hígado. La irrigación funcional ocurre por la vena porta hepática, a través de la cual ingresa sangre venosa proveniente del tracto gastrointestinal. Esta sangre contiene la mayoría de las sustancias que han sido absorbidas por el intestino y que en su trayecto hacia la circulación sanguínea principal, son captadas por la célula hepática. Estas sustancias intervienen en múltiples procesos metabólicos, de excreción y endocrinos (Geneser, 2001).
Cabe destacar, que en el hígado se realizan entre otras funciones, la producción de sustancias de naturaleza hormonal, procesos metabólicos como la glucólisis y gluconeogénesis, metabolismo lipídico, procesos de transporte, destoxificación, síntesis de vitaminas; y durante el desarrollo embrionario, la función hematopoyética (Geneser, 2001).
Basado en lo anteriormente expuesto, el propósito de este trabajo fue el de caracterizar morfológica e histoquímicamente el hígado del Caiman crocodilus crocodilus.
Materiales y Métodos
Los animales utilizados en esta investigación, fueron tratados respetando las normas éticas que rigen el empleo de animales de investigación (AVECAL, 2008). El material biológico empleado en este trabajo consistió de ocho babas adultas, cinco hembras y tres machos, clínicamente sanos. Los animales provinieron de la laguna de oxidación de la Facultad de Agronomía de la Universidad Central de Venezuela (UCV), Maracay, estado Aragua, y fueron capturados mediante el empleo de anzuelo con carnada. Como protocolo anestésico, se utilizó 0,5 g de Tiopental sódico (SM Pharma, Venezuela), a una dosis de 200 mg/kg de peso corporal, por vía intraperitoneal, hasta alcanzar un plano anestésico satisfactorio. El abordaje de la cavidad celómica, se realizó colocando al animal en decúbito dorsal y se efectuó una incisión a nivel de la línea media, luego se disecaron la piel y los músculos abdominales. Al abordar la cavidad, se observó la anatomía macroscópica del hígado y su relación con el resto de los órganos presentes se tomaron fotografías correspondientes, con una cámara digital Nikon Coolpix 950 de 2,1 mega píxeles. Posteriormente, se procedió a la eutanasia, inoculando una sobredosis de tiopental sódico vía intravenosa (vena coxígea), para seguidamente realizar la toma de muestras. Se obtuvieron muestras de 2 cm2 de ambos lóbulos hepáticos y se fijaron en formol al 10% v/v, a pH 7,1. El procesamiento histológico se llevó a cabo en el Laboratorio de Histoquímica de la Cátedra de Histología de la Facultad de Ciencias Veterinarias de la UCV. Las muestras de hígado fueron incluidas en parafina, según los métodos histológicos convencionales (Prophet et al., 1995). De los bloques obtenidos, se realizaron cortes de 4 micrómetros de grosor, utilizando un microtomo de deslizamiento marca JUNG. Las muestras se tiñeron usando la coloración de rutina hematoxilina-eosina (H-E), y se hicieron cortes seriados para la observación de la trayectoria de los cordones de hepatocitos, así como también, de los vasos sanguíneos y los conductillos hepáticos. Para evidenciar las fibras colágenas, se utilizó la coloración especial de Tricrómico de Gallegos (TG) y para evidenciar las fibras reticulares, se empleó el método de Wilder (MW). Asimismo, se realizó la toma de muestras de 2 cm2 para cortes por congelación, los cuales se realizaron en un criostato Leica modelo CM1510-3, utilizándose la tinción especial de aceite rojo O (ARO) para la visualización de lípidos (Prophet et al., 1995).
Los cortes histológicos fueron observados a través del microscopio óptico y las áreas de interés se fotografiaron bajo un fotomicroscopio Nikon Eclipse E400 con cámara Nikon Coolpix 950 de 2,1 mega píxeles, obteniéndose las fotografías para el estudio morfológico.
Resultados y Discusión
Los resultados del presente estudio morfológico del hígado de la baba, revelaron que este órgano está conformado por dos lóbulos, derecho e izquierdo, unidos por un istmo, tal como lo reporta Rossini (2003). Este órgano se encuentra ubicado anatómicamente en la línea media de la cavidad celómica caudal (Figura 1).

Figura 1. Hígado del Caiman crocodilus crocodilus, LD lóbulo derecho, LI lóbulo izquierdo
Con la coloración H-E se observó un parénquima, en el cual la lobulación clásica del lobulillo hepático, se encuentra ausente (Figura 2). Esta observación coincide con lo reportado por Rossini (2003), quien no pudo detectar la presencia del lobulillo hepático en esta misma subespecie. Los hepatocitos se describen como células globosas con núcleo en posición basal (Figura 3), similar a lo observado en el Scleropus occidentalis biseriatus (Ells, 1954). En la subespecie estudiada, se evidenció la presencia de un citoplasma eosinofílico con espacios claros, atribuibles a la presencia de gotas lipídicas. Estas observaciones coinciden con las reportadas en otros reptiles, tales como el Scleropus occidentalis biseriatus y el Caiman crocodilus crocodilus, las cuales vinculan la presencia de contenido lipídico con los espacios claros intracitoplasmáticos de los hepatocitos (Ells, 1954; Rossini 2003); sin embargo, Orós (2002), relacionó los espacios claros de los hepatocitos con la presencia de agua intracitoplasmática. El núcleo del hepatocito es vesiculoso, con presencia de uno o dos nucléolos, lo cual coincide con lo señalado por Orós (2002).
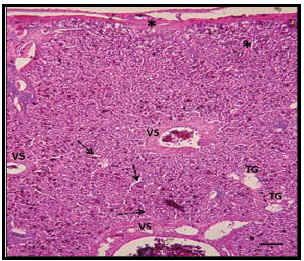
Figura 2. Micrografía del hígado de baba. Hematoxilina-Eosina. VS: Vasos sanguíneos (vena central o vena tributaria). TG: tejido graso. Sinusoides hepáticos (flechas punteadas). Tejido conectivo (*). Barra 50m.
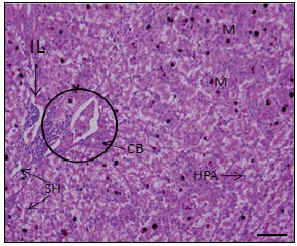
Figura 3. Aspecto microscópico del hígado de baba. Hematoxilina-Eosina. HPA: Hepatocitos en patrón acinar. SH: Sinusoide hepático. CB: Conductillo biliar. M: Macrófagos cargados de melanina. IL: infiltrado linfocitario. Tríada portal (circulo). Barra 50 mm
Del estudio de los cortes seriados, se pudo evidenciar que los cordones hepáticos presentan un trayecto tortuoso, similar a lo observado en el hígado del Scleropus occidentalis biseriatus, según lo reportado por Ells (1954), encontrándose a los hepatocitos enfrentados por su superficie apical, creando un canalículo biliar y por su superficie basal, se relacionan con los vasos sinusoidales, lo cual les confiere una disposición acinar en orientación transversal (Figura 3).
En estudios realizados en otras especies, como en gallos, en cortes de hígados efectuados transversal y tangencialmente, las células aparecen en forma vagamente triangular, agrupadas en número de cuatro a seis, alrededor de los canalículos biliares (Delhon et al., 1984), coincidiendo con los resultados de este estudio. Esta disposición acinar de los hepatocitos, podría estar relacionada con la secreción exocrina de bilis del órgano, tal como lo cita Orós (2002) quien trabajó en otras especies de reptiles como Python molurus.
Con relación a la estructura de la red vascular, se notó la presencia de vasos con una luz amplia, con paredes muy delgadas, rodeados de hepatocitos, siendo similares a los observados en el Scleropus occidentalis biseriatus, (Ells, 1954) y a los vasos sinusoidales de los mamíferos (Geneser, 2001). Además, se encontraron venas con luz amplia, similares a las descritas por Ells (1954), a las cuales les denominó venas tributarias, que se corresponden funcionalmente, pero no morfológicamente con las venas centrales en mamíferos. En estas venas tributarias hepáticas confluyen numerosas ramas de un reticulado sinusoidal (Figura 2). Asimismo, se observan áreas equivalentes a los espacios porta de los mamíferos, dispersas en el parénquima hepático que contienen ramas venosas, ramas arteriales muy delgadas y uno o dos conductillos biliares dispuestos transversalmente, los cuales se evidencian por la presencia de un epitelio cúbico simple. Todo este espacio porta está rodeado por tejido conectivo. En todos los hígados estudiados, se observó un infiltrado linfocitario perivascular moderado, cercano a los espacios porta (Figura 3).
En la Figura 3, se muestran numerosas estructuras de aspecto redondeado y de color pardo oscuro, dispersas en todo el parénquima, compatibles con melanóforos, macrófagos cargados de pigmento, muy frecuentes en la circulación sistémica, comunes en las diversas especies de reptiles y peces estudiadas por Ells (1954) y Lagler et al. (1990).
El hígado se encuentra cubierto por una membrana extremadamente delgada de tejido conectivo, la cual se corresponde con la cápsula hepática de los mamíferos. Este tejido penetra al órgano superficialmente, formando trabéculas incompletas, lográndose observar además, una delgada membrana a nivel de la vena central o tributaria y áreas portales (Figura 2).
Con la coloración especial de TG, en el tejido hepático se observan las fibras colágenas de color verde intenso, confirmando la presencia de tejido conectivo rodeando al parénquima hepático, el cual penetra en forma de trabéculas incompletas; además, las fibras colágenas se disponen finamente entre los cordones de hepatocitos, haciéndose mucho más gruesas cuando rodean a los vasos sanguíneos y a los conductillos biliares (Figura 4). Adicionalmente, se demostró la trama reticular del tejido hepático con el MW, observándose que las fibrillas reticulares se disponen en forma paralela y muy delgada, rodeando a los hepatocitos. Las áreas con mayor densidad de fibras reticulares, indican la presencia de vasos sanguíneos y canalículos biliares (Figura 5).

Figura 4. Micrografía del hígado de la baba. Tinción: Tricrómico de Gallegos. FC: Fibras colágenas. Fibras colágenas que se disponen finamente entre los cordones de hepatocitos (*) PH: Parénquima hepático. Barra 50 mm

Figura 5. Micrografía del hígado de la baba. Método de Wilder. FR: Fibras reticulares, (flechas) Barra 50mm
Con respecto a éstas dos últimas observaciones, no se encontraron estudios previos relacionados con coloraciones especiales que evidencien el estroma hepático de reptiles.
Con la coloración especial de ARO, se confirmó la presencia de gotas lipídicas intracitoplasmática, las cuales han sido consideradas como el almacén principal de lípidos en las células eucariotas (Martin y Parton, 2006). Dichas gotas se observan de diversos tamaños y en algunos casos ocupan casi la totalidad del citoplasma de los hepatocitos, lo cual está acorde por lo citado por Ells (1954) y Rossini (2003), y en contraposición con los hallazgos en reptiles, en los cuales se ha observado la presencia de agua intarcitoplamática (Orós, 2002). Otras investigaciones han revelado, mediante microscopía electrónica, la presencia de abundantes gotas lipídicas citoplasmáticas en los hepatocitos del hígado del Scleropus javorri, como lo refiere Goldberg (1972). En este estudio, también se evidenció la presencia de lípidos, incluso en los vasos sinusoidales y las células de los conductillos hepáticos (Figura 6). La presencia de este material lipídico podría asociarse a los hábitos alimenticios de estos reptiles, consumen gran cantidad de alimento, hasta más del 100% de su propia masa corporal y se mantienen en ayunas hasta por 18 meses; los reptiles que consumen alimento poco frecuente tienen proporcionalmente mayores respuestas reguladoras a la alimentación (Uriona et al., 2005). Otro aspecto importante que debe ser tomado en cuenta son los cambios estacionales en el hábitat y la abundancia o escasez de oferta de alimento que afectan la dieta de estos reptiles (Allsteadt y Vaughan-Dickhaunt, 1994). Estas subespecies, en la época de abundancia de alimento, almacenan lípidos en el órgano graso; y en el período de sequía en el que hay escasez de alimento, movilizan los lípidos almacenados desde el órgano graso hacia el hígado donde son metabolizados, similar a lo que ocurre en caso de ayuna prolongada de las babas, que conlleva a la movilización de los lípidos presentes en el órgano graso, lo cual explica la abundancia de material lipídico de las muestras de hígados tomadas, lo cual coincidió con la época de sequía. Otro aspecto a considerar, según G. García (comunicación personal), se relaciona con las necesidades energéticas de la subespecie, ya que la presencia de insectos tipo coleópteros en babas estudiadas de diferentes tallas, se relaciona con la necesidad del aporte energético proveniente de las grasas presentes en los insectos y su posterior almacenamiento en hígado y órgano graso. Estas subespecies se encuentran en espacios con una alta oferta y escasez de alimentos durante los períodos de lluvia y sequía respectivamente, por lo que se sugiere realizar estudios no solo morfológicos, sino bioquímicos y fisiológicos que aclaren las observaciones presentadas en este estudio. Asimismo, la presencia de estas gotas lipídicas no es excluyente de la presencia de los componentes subcelulares típicos de un hepatocito, ya que la presencia del lípido presupone el desarrollo de un retículo endoplasmático liso y de mitocondrias asociados al metabolismo de grasas, lo cual no interfiere con las funciones del hepatocito.

Figura 6. Micrografía del hígado de la baba, coloración aceite rojo O. PH: Parénquima hepático, a. Barra 20 mm. Canalículo biliar (*) b. Barra 50mm
Conclusión
El hígado de la baba, está ubicado en la línea media de la cavidad celómica caudal, es un órgano bilobulado, con los lóbulos derecho e izquierdo unidos por un istmo y está rodeado por una delgada cápsula de tejido conectivo que penetra al tejido de forma incompleta. El parénquima hepático no presenta la disposición del lobulillo hepático clásico y los hepatocitos se disponen en cordones tortuosos y con un patrón acinar sobre un estroma de fibras reticulares. Adicionalmente, existen melanóforos dispersos en el tejido hepático y hepatocitos cargados de gotas lipídicas intracitoplasmáticas de diversos tamaños, y cantidades, las cuales pueden explicarse por los hábitos alimenticios y las condiciones metabólicas de almacenaje y movilización posterior de grasas de estas subespecies, además de las condiciones ambientales de su entorno en relación a la disposición de alimento. En tal sentido, se podrán realizar en el futuro, investigaciones, utilizando el hígado como modelo experimental con el fin de realizar estudios de las diferentes rutas metabólicas que ocurren en este órgano.
Agradecimientos
Este estudio fue financiado a través del proyecto N° PI 035/2009-PI-11-7296-2008/1, otorgado por el CDCH. Se agradece al Señor Francisco García, del Laboratorio de Histoquímica de la Cátedra de Histología de la FCV-UCV, por el procesamiento de las muestras y, especialmente, al Dr. Jesús Rojas Urbina, por la revisión de este manuscrito.
Referencias
1. AVECAL. 2008. Manual para la Producción y Uso Ético de los Animales de Laboratorio. Gobierno Bolivariano de Venezuela. Ministerio del Poder Popular para la Ciencia y Tecnología. 93 p. [ Links ]
2. Allsteadt, J.; Vaughan-Dickhaut, C. 1994. Food habits of Caiman crocodile in Caño Negro, Costa Rica. Vida Silvestre Neotropical, 3:24-29. [ Links ]
3. Banks, W. 1986. Histología Veterinaria Aplicada. El Manual Moderno. México, D.F. 730 p. [ Links ]
4. Brazaitis, P. 1973. The identification of living crocodilians. Zoológica, 58:59-101. [ Links ]
5. Britton, A. 2002. Crocodilian Biology Database. Página en línea. Disponible en:<http://www.flmnh.ufl.edu/natsci/herpetology/brittoncrocs/cnhc.html>. Consultada el 20 de Octubre 2008. [ Links ]
6. Cabrera, F.; García, G. 2004. Características anatómicas del aparato genital masculino del Caiman crocodilus crocodilus. Rev. Fac. Cs. Vets. UCV, 45:55-63. [ Links ]
7. Cabrera, F.; García, G.; González-Vera, M. Rossini, M. 2007. Características histológicas del aparato genital masculino del Caiman crocodilus crocodilus. Rev. Cien. FCV-LUZ, XVII:123-130. [ Links ]
8. Cabrera, F.; García, G. 2007. Mucinas del aparato genital masculino de la baba (Caiman crocodilus crocodilus) Rev. Fac. Cs. Vets. UCV, 48:77-84. [ Links ]
9. Delhon, G.; Fernández, J.; Gangna, G.; Lacolla, D.; Von Lawzewitsh, I. 1984. Lecciones de Histología Veterinaria. Aparato Digestivo Comparado: Estómago de Monogástricos, preestómagos, aves, glándulas anexas. Vol 5. 3era ed., Hemisferio Sur S.A. Argentina, 111p. [ Links ]
10. Ells, H. 1954. The gross and microscopic anatomy of the liver and gall bladder of the lizard Scleropus occidentalis biseriatus (Hallowell) Anat. Rec., 119:213-229. [ Links ]
11. Geneser, F. 2001. Histología sobre las Bases Moleculares. 3ª Ed. Médica Panamericana. Buenos Aires. 813 p. [ Links ]
12. Goldberg, S. 1972. Seasonal weight and cytological changes in the fat bodies and liver of the iguanid lizard Scleropus jarrovi. Copeia, 2:227-232. [ Links ]
13. Lagler, K.; Bardach, J.; Millers, R.; May Passino, D. 1990. Ictiología. 1era ed. en Español. AGT. México. 489 p. [ Links ]
14. Martin, S.; Parton, R. 2006.Lipid droplets: a unified view of a dynamic organelle. Cell Biology, 7:373-378. [ Links ]
15. Orós, J. 2002. Aparato digestivo en línea. http://www.5.ulpgc.es/servidores/histología/reptiles/Cosultada el 28 de enero 2010. [ Links ]
16. Prophet, E.; Milis, B.; Arrington, J.; Sobín, L. 1995. Métodos Histotecnólógicos. Instituto de Patología de las Fuerzas Armadas de los Estados Unidos de América (AFIP).Washington, D.C. 400 p. [ Links ]
17. Ross, C.; Magnuson, W. 1989. Living Crocodilians. In: Crocodiles and Alligators. Facts on File, New York, EEUU. C. A. Ross CA (ed.). pp. 58-63. [ Links ]
18. Rossini, M. 2003. Características macroscópicas, histológicas e histoquímicas del sistema digestivo de la baba (Caiman crocodilus). Tesis de Maestría, Postgrado en Medicina Veterinaria, mención Patología. FCV UCV-. Maracay estado Aragua, Venezuela. 119 p. [ Links ]
19. Rossini, M.; García, G. 2010. Descripción morfológica de las células sanguíneas de la baba (Caiman crocodilus crocodilus) en vida silvestre. Rev. Fac. Cs. Vets. UCV, 51:63-70. [ Links ]
20. Uriona, T.; Framer, C.; Dazely, J.; Clayton, F.; Moore J. 2005. Structure and function of the esophagus of American alligator (Alligator mississippiensis). J. Exp. Biol., 208:3047-3053. [ Links ]
21. Velasco, A.; Ayarzagüena, J. 1992. Population status of the spectacled caiman (Caiman crocodilus) in the Venezuelan plain. In: Crocodiles. Proc. 11th Working Meeting. Crocodiles Specialist Group. IUCN/ The World Conservation Union. Gland, Switzerland. Vol. II. pp. 184-199. [ Links ]
22. Velasco, Á.; Colomine, G.; De Sola, R. 2003. Effects of sustained harvests on wild populations of Caiman crocodilus crocodilus in Venezuela. INCI., 28:544-548. [ Links ]
