










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkGaceta Médica de Caracas
versión impresa ISSN 0367-4762
Gac Méd Caracas v.113 n.1 Caracas ene. 2005
La lámina basal trofoblástica y su relación con la ultraestructura del trofoblasto
Dr. Olivar C Castejón*
*Coordinador general del CIADANA. Prof. Titular en biología celular. Fac. Cs de la Salud, Universidad de Carabobo.
RESUMEN
Con el propósito de evaluar la lámina basal trofoblástica en vellosidades placentarias a término se tomaron biopsias de cinco placentas inmediatamente después del parto vaginal normal para ser procesadas según las técnicas convencionales de microscopia electrónica de transmisión y el método Gaboul para determinar polianiones con el azul alcián.
Los resultados muestran una banda densa de variable grosor, homogénea, íntimamente asociada a la membrana plasmática del trofoblasto estando ausente la lámina lúcida. Fibrillas de colágeno contactan la lámina densa por la superficie estromal. En zonas de intensa actividad de biosíntesis del trofoblasto y de incrementada transferencia de sustancias, la lámina densa sufre notables cambios, pierde su uniformidad y emite prolongaciones en ambas superficies. El método Gaboul determinó la ausencia de grupos aniónicos en la lámina densa indicando el pasaje libre de macromoléculas hacia el compartimiento fetal capilar. La actividad del trofoblasto parece cambiar el grosor y la estructura de la lámina densa.
Palabras clave: Lámina basal trofoblástica. Vellosidad. Placenta humana.
SUMMARY
To evaluate the trophoblastic basement membrana in placental villi at term specimens were taken from five placentas inmediately after normal vaginal delivery to be processed according to conventional transmission electron microscopy and the Gaboul method to determine polyanionic sites with the alcian blue. The findings showed a densa lamina of variable trickness, uniform, closely associated to the trophoblastic plasma membrane. The lucid lamina was absent. The fibrils of collagen contact the dense lamina throughout the stromal suface. In regions of increased trophoblastic biosynthetic acti-vity the dense lamina suffers important changes, lose uniformity and emits extensions in both surfaces. The Gaboul method determined absence of anionic sites in the dense lamina indicating a free transport of macromolecules to the fetal capillary compartment. This trophoblastic activity seems to change the thickness and the structure of the dense lamina.
Key words: Trophoblastic basement membrane. Villi. Human placenta.
INTRODUCCIÓN
La lámina basal (LB) trofoblástica separa el epitelio trofoblástico del estroma velloso y está constituida por colágeno IV, laminina, heparán sulfato y puede asociarse con otras proteínas de la matriz extracelular como fibronectina y tenascina (1).
La estructura, composición, desarrollo y las inclusiones de esta lámina, en general, han sido brevemente descritas con relación a la ultraestructura de la placenta humana normal (2). La mayoría de las descripciones tratan con el número de sus capas en las etapas tempranas del embarazo y el pasaje de sustancias a través de ella (3).
Entre los epitelios y la matriz extracelular del estroma de otros tejidos, mediante la microscopia electrónica de transmisión (MET), se ha observado que la LB se compone de varias capas o láminas. La lámina densa, una banda oscura de 30-100 µm de grosor. Entre esta y la célula asociada hay una zona clara llamada lámina lúcida, de aproximadamente 60 µm de ancho. Por el otro lado de la lámina densa hay una capa de variable grosor denominada lámina fibrorreticular la cual está asociada a proteínas fibrosas de la matriz extracelular (4).
La estructura vista con la microscopia de luz, teñidas con ácido paramino salicílico y nitrato de plata (5), referida como membrana basal, es una combinación de todas estas láminas. El término LB en este trabajo se refiere a la lámina densa descrita ultraestructuralmente y mencionada antes. Los términos membrana basal y lámina basal serán usados aquí indistintamente.
En condiciones normales, según la revisión bibliográfica, el grosor promedio de la LB está entre 150-500 µm. El estudio de los cambios morfológicos que experimenta esta estructura en momentos de elevada transferencia de fluido o de incrementada actividad metabólica orienta nuestra atención con el objeto de encontrar si hay correlación entre la dinámica funcional del trofoblasto y la barrera física representada por la LB que lo sostiene. Esto contribuiría a nuestro entendimiento del comportamiento de esta estructura en las anomalías del trofoblasto (5). El trofoblasto molar es de elevada actividad biosintética y será de interés correlacionar su ultraestructura y la de su LB con la finalidad de obtener información sobre la transformación maligna. Para lograr estos objetivos la técnica de estudio ultraestructural será seguida por un método de histoquímica ultraestructural.
Se conoce que la LB trofoblástica (6) presenta proteglucanos polianiónicos que son responsables de impedir, como una barrera, la entrada de macromoléculas cargadas negativamente hacia el estroma. Con el uso del método Gaboul (7) que emplea azul alcián a pH 2,5 podemos detectar si la LB presente en vellosidades placentarias a término, demuestran estos grupos aniónicos. La no reacción del azul alcián con los componentes de carga negativa nos indicaría su ausencia de la LB y por tanto que en esta clase de vellosidades terminales del final del embarazo, el pasaje de importantes macromoléculas estaría permitido. Observar y evaluar secciones de vellosidades a término mostrando la LB en condiciones normales con la técnica de MET y el método Gaboul constituyen el propósito de este estudio.
MATERIALES Y MÉTODOS
Placentas de cinco embarazos a término, entre 37 a 40 semanas de gestación, fueron recolectadas inmediatamente después del parto vaginal normal y se tomaron especimenes de la región central-parabasal y periférica de la placa basal. Estos segmentos de tejidos se lavaron brevemente en solución salina para remover la sangre materna procesándose según la técnica de MET de acuerdo con trabajo previo (8). Otras biopsias se emplearon para determinar polianiones en la LB según el método Gaboul (7) de vellosidades cercanas a la placa basal. El conjunto de procedimientos en el cual el material se fija en glutaraldehido, posteriormente teñido en azul alcián, posfijado en solución de tetraóxido de osmio y doblemente teñido con acetato de uranilo y citrato de plomo ha sido denominado método Gaboul. Las secciones tomadas para estudio se seleccionaron perpendiculares a la superficie del trofoblasto o eje longitudinal del mismo, fueron examinadas en un microscopio electrónico Hitachi H-500 a 80 kV, provisto de una apertura de 50 µm en el objetivo. Las mediciones realizadas en la LB corresponden a la lámina densa únicamente.
Las micrografías fueron tomadas en un rango de 2 000 x a 60 000 x, de zonas pertenecientes a membranas vasculo-sincitiales, placas epiteliales o zonas de elevada actividad de transferencia placentaria localizadas en vellosidades terminales.
RESULTADOS
Técnica de MET: la lámina densa mostró variabilidad en su grosor el cual se expresa en un rango de 0,05-0,79 µm y un promedio de 0,42 µm. En algunos casos se observa dividida en bandas. La lámina lúcida está casi ausente y a veces no se observa. La lámina densa se observó uniforme, de aspecto gránulo-filamentosa, de variable grosor (Figura 1).
En numerosas micrografías se nota que la uniformidad de la lámina densa como una banda, se modifica y numerosas prolongaciones de ella sobresalen en ambas superficies hacia el trofoblasto o hacia el estroma (Figura 2). Estos cambios coinciden con amplias cisternas de retículo endoplasmático rugoso (RER) cuyo contenido subcisternal es electrón opaco y pertenecen al trofoblasto que descansa sobre ella. Sustancias electrón opacas, de diversos tamaños, que cruzan la lámina densa se observaron en momentos en que hay una correspondencia con las variaciones de su aspecto o grosor (Figura 3). En algunas observaciones se notaron fragmentos de lámina lúcida en el estroma de la vellosidad (Figura 4).

Figura 1. Región del citotrofoblasto que descansa sobre la lámina basal cuya zona más ancha es producto del corte. La flecha señala microfibrillas de colágeno en región fibrorreticular. Barra: 0,5 µm.
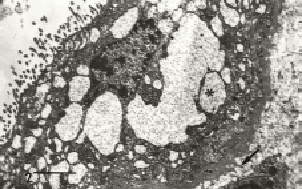
Figura 2. Retículo endoplásmico rugoso con un contenido electrón opaco gránulo-filamentoso se observa al momento en que la lámina basal presenta proyecciones de lámina densa (flecha). Barra: 2 µm.

Figura 3. Gránulos electrón opacos (flecha) de diversos tamaños cruzan la lámina densa. Algunos quedan atrapados entre "dedos" del sincitio. Barra: 1 µm.

Figura 4. Una zona de membrana plasmática sincitial que no constituye prolongaciones filiformes (flecha) se observa cercana a otra con prolongaciones (cabeza de flecha). Franjas de lámina densa se localizan en el estroma. Barra: 2 µm.
A lo largo de la LB, se observan dos regiones muy distintas de organización de la membrana plasmática del sincitio o del citotrofoblasto. En una se nota lisa, sin pliegues, descansando sobre la LB. En otra región a corta distancia, se exhibe con numerosos pliegues filiformes, paralelos, semejando podocitos contactando la lámina densa (Figura 4, 5).
Estas regiones parecen estar cumpliendo una función diferente. Por otro lado, numerosas prolongaciones del sincitio o trofoblasto celular fueron vistas incorporadas a la estructura de la lámina densa. Hay imágenes donde las prolongaciones del citotrofoblasto recorren cierta distancia en su espesor (Figura 6). No se logró observar prolongaciones del trofoblasto que sobrepasen el espesor y contacten prolongaciones de células estromales o viceversa como ocurre, por ejemplo, entre células foliculares y el ovocito II, en el ovario humano, separados por una LB de notable grosor.
Prolongaciones de células de Hofbauer o de miofibroblastos cercanas a la lámina fibrorreticular no parecen atravesarla.
Método Gaboul: la LB se mostró con un componente granulofilamentoso aún más fino que el observado con la técnica de MET convencional. Las regiones de membrana plasmática sincitial o celular, incorporadas a la lámina densa se notan como una línea punteada irregularmente electrón opaca. Estos granulos irregulares son reactivos al azul alcián y por tanto alcianofílicos (Figura 7).
La región de la LB, la fibrorreticular, que mira hacia el estroma presenta numerosas microfibrillas de colágeno con el punteado alcianofílico. Todo el resto de la lámina densa permanece no reactiva al azul alcián, de manera homogénea (Figura 8). Vesículas de pinocitosis, vesículas cubiertas, huecos o depresiones cubiertas en las membranas del trofoblasto cercanas a la lámina lúcida son indicativas de una dinámica en la transferencia de fluidos (Figura 7).

Figura 5. Las regiones A y B muestran zonas de prolongaciones filiformes, asociadas a una lámina basal de aspecto reticulado en B, con una imagen que semeja liberación de segmentos de lámina densa hacia el estroma (flecha). Barra: 2 µm.
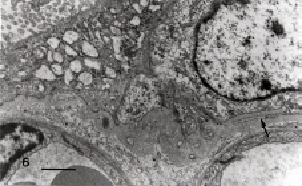
Figura 6. La flecha señala una prolongación del citotrofoblasto que recorre cierto trayecto en el espesor de la lámina densa exhibiendo la micrografía otros cortes a lo largo de su recorrido. Barra: 1 µm.

Figura 7. Corte de membranas plasmáticas del citotrofoblasto se muestran alcianofílicas (flecha curva) en el espesor de la lámina densa. Vesículas cubiertas se observan en el trofoblasto celular (cabezas de flecha). Método Gaboul. Barra: 0,5 µm.

Figura 8. La región de la lámina basal (LB) se nota negativa al azul alcián. Fibrillas de colágeno exhiben un punteado alcianofílico. El filipodio del macrófago se incrusta en la LB sin atravesarla (flecha). Método Gaboul. Barra: 0,5 µm.
DISCUSIÓN
La LB trofoblástica ha sido evaluada con la idea de que una elevada transferencia de sustancias a través de su espesor y de una incrementada actividad de biosíntesis del trofoblasto son factores que modifican la ultraestructura de la misma y que las variaciones de su espesor y estructura son de interés para el entendimiento de ellas en la anomalía del trofoblasto molar. En los primeros estudios ultra-estructurales (3), donde se discute la transferencia de sustancias, se pensó que la ausenta de vesículas o plaquetas de lípidos era producto en la LB, de la ausencia de circulación en las etapas tempranas del embarazo, confirmada también en los casos de mola hidatidiforme.
Durante el transcurso del embarazo la LB incrementa su grosor, quedando conformada por capas membranosas, perdiéndose en parte su permeabilidad a término. El grosor depende de la etapa del embarazo, el tamaño de la vellosidad y la oblicuidad de la sección en relación al eje longitudinal de la vellosidad estando en un rango de 0,22 µm, en la etapa embrionaria de 13 µm hasta 0,95 µm a término (10). El valor promedio del grosor acá encontrado en la lámina densa, está por debajo del rango medido por otros (4) pero como estas mediciones han sido hechas en especímenes fijados en aldehídos durante el proceso, sufren una contracción o arrugamieno del 30 % (11) que provoca también la desaparición de la lámina lúcida. Estos valores en realidad están por debajo del valor real.
Se requiere actualizar estas mediciones utilizando técnicas criogénicas combinadas con MET. La MET de alta resolución revela que la lámina densa trofoblástica es una malla poligonal de colágeno IV que tiene polígonos de 18 µm de lado, aislada de la placenta humana mediante tratamiento con pepsina (11). Fibrillas de colágeno desde el conjuntivo del estroma estabilizan la lámina densa (12).
Las inclusiones o depósitos de glóbulos de grasa, lípidos, gránulos de calcio o hierro han llamado la atención de numerosos investigadores cuando se hace referencia a la LB en la revisión de la ultraestructura placentaria (2). Morfológica e histoquímicamente se han demostrado depósitos de hierro en la LB (13-15) en forma de gránulos y cuya presencia en exceso, durante el segundo y tercer trimestre del embarazo es pronóstico de anomalía del desarrollo fetal (14). La MET asociada con microanálisis de rayos X nos da una inequívoca diferenciación de la naturaleza de estos depósitos. El análisis de abundantes plaquetas de lípidos que se liberan desde el trofoblasto hacia la LB, con estas técnicas revelaron espectros de rayos X que contienen picos de hierro durante la séptima semana de gestación (15). La LB es así un reservorio temporal de sustancias, mientras los capilares recién formados del estroma vellositario tienen poca habilidad para transportar el hierro hacia la circulación fetal, en las primeras semanas del desarrollo. Al activarse los capilares se dificulta la observación del pasaje de sustancias en la lámina densa.
El mecanismo de tinción del azul alcián, una ftalocianina de cobre, se fundamenta en que su molécula posee cuatro grupos positivos isotiouronium que al combinarse con los grupos aniónicos existentes en la estructura a teñir, fijada en tetraóxido de osmio origina un precipitado negro, a pH 2,5, si es utilizado con este grado de acidez. Debido a que la membrana plasmática del trofoblasto contiene sobre su superficie grupos carboxílicos del ácido siálico, las cargas negativas de éstas reaccionan con el azul alcián dando precipitados en forma punteada (7). Por otro lado, como las prolongaciones de membrana plasmática del trofoblasto penetran la LB es por esto que se observa la línea punteada en el espesor de la lámina densa; mientras que ésta permanece no reactiva al colorante. Esto nos está indicando que la LB carece de grupos aniónicos, el heparán sulfato de la LB está ausente o disminuido a término. Este glucosaminoglucano se localiza en la lámina densa entre 7-12 semanas de gestación (6) y es la macromolécula que principalmente contribuiría a dar reactividad con el colorante. Nuestros resultados sugieren que la LB de la vellosidad placentaria, a diferencia de la LB del glomérulo renal, se adapta al pasaje de sustancias al final del embarazo coincidiendo con lo reportado por King (6) con el rojo rutenio.
Dado que polipéptidos, proteínas, anticuerpos y otras macromoléculas tienen libre paso y un epitelio fuertemente secretor como lo es el trofoblasto, en una etapa del desarrollo fetal donde los intercambios materno-fetales son más rápidos e intensivos, es natural que la lámina densa se vea afectada en su organización como lo indican los resultados. Si bien la placenta ha llegado a la etapa final de su ciclo de vida todavía hay un transporte intracelular intenso y descarga de productos secretorios provenientes de la actividad de síntesis del RER del trofoblasto; actividades que pudieran ser factores de desestabilización de la LB o de sus cambios de forma. Esto estaría confirmado por los restos de lámina densa encontrados en el estroma vellositario. El elevado peso fetal alcanzado a término amerita una intensa actividad de transporte que en ambos sentidos termina por desorganizar, en parte, la lámina densa lo que traería como consecuencia una situación de edema.
Durante las etapas más tempranas de la gestación estos cambios fisiológicos están incrementados y las constantes remodelaciones provocadas por la ramificación del árbol velloso (16) impactan sobre el papel mecánico de sostén de la lámina densa que se asocia al esqueleto fibrilar colagenoso estromal el cual conforma la matriz extracelular, sostén del trofoblasto (17). El colágeno, aquí observado, en la región fibrorreticular, reactivo al azul alcián, muy probablemente por su asociación con heparán sulfato, es parte de esta matriz extracelular que ha sido recientemente visualizada en forma tridimensional con microscopia electrónica de barrido en vellosidades a término (18).
Las zonas de numerosas prolongaciones del citoplasma del trofoblasto, descansando como estructuras en forma de dedos sobre la LB, en paralelo, han sido correlacionadas con el movimiento de fluidos, al igual que los túbulos renales pero a diferencia de estos las prolongaciones no contienen mitocondrias (13). La observación de gránulos complejos de hierro atrapados en la lámina densa y rodeados por estas estructuras filiformes pudieran expresar además un rol distinto al de la transferencia de fluido (13); como el filtraje de grandes partículas en vellosidades a término. En los resultados vistos hemos podido apreciar (Figura 3) la participación de estas estructuras en su rol de filtro. El aumento de superficie de membrana por aumento de prolongaciones, trae como consecuencia una mayor disponibilidad de cargas en la membrana trofoblástica que pueden electrostáticamente atraer cualquier partícula que traspasa la LB y contribuyendo así a la acción de filtrado en cualquiera de las dos direcciones.
En vista de que la laminina o el colágeno IV pueden ser producidos, tanto por el trofoblasto celular como por los miofibroblastos estromales, se trató de observar si habían contactos entre prolongaciones del trofoblasto y la de alguna célula en el interior de la vellosidad con la finalidad de evidenciar algún intercambio de información que estuviese en correlación con la deposición o elaboración de componentes de la LB. Estos contactos quedaron a ser demostrados.
La relación colágeno IV/laminina que resulta elevada en la preeclampsia refleja disminución en la concentración de laminina (19) lo que pudiera debilitar la unión del trofoblasto a la lámina densa, modificando las propiedades de intercambio y permeabilidad de la vellosidad. Si aplicaramos las técnicas de inmunofluorescencia y radioinmunoensayo con las cuales se encontró esta relación a placentas de pacientes con preeclampsia (20) posiblemente podría encontrarse una explicación al retardo del crecimiento intrauterino que suele notarse en ellos (21).
Como la LB es una estructura dinámica que constantemente se cataboliza y se redeposita, es de nuestro interés conocer su integridad en los casos de elevada producción hormonal por el trofoblasto, en cuya entidad es necesario dilucidar si la interrupción de la membrana basal, por ejemplo, es producto del pasaje de sustancias por su incrementada actividad de biosíntesis o bien por la transformación maligna. De esta manera se conoce que un componente de la LB, la laminina M, bajo regulación hormonal (22) ha proporcionado información sobre el carácter neoplásico (5). La laminina-1 además está implicada en la etiología y estructura de la mola hidatidiforme (23). Los resultados reportados en este trabajo expresan que la transferencia de sustancias o macromoléculas a través de la lámina densa en vellosidades a término provocan modificaciones en su grosor o aspecto en momentos en que la actividad del trofoblasto está metabólicamente incrementada.
REFERENCIAS
1. Benirschke K, Kaumann P. Pathology of the human placenta. 4ª edición. New York: Springer-Verlag; 2000. [ Links ]
2. Jones CJ, Fox H. Ultrastructure of the normal human placenta. Electron Microsc Rev. 1991;4:129-178. [ Links ]
3. Knoth M. Ultrastructure of chorionic villi from a four somite human embryo. J Ultrastruct Res. 1968;25:423-440. [ Links ]
4. Stevens A, Lowe JS. Human Histology. 2ª edición. Londres: Mosby; 2000. [ Links ]
5. Castejón OC, Scucces MG, Rivas AE, Graterol IJ. La membrana basal en caso de mola hidatidiforme completa. Rev Obstet Ginecol Venez. 2002;62:207-213. [ Links ]
6. King BF. Distribution and characterization of anionic sites in trophoblast and capillary basal laminas of human placental villi. Anat Rec. 1985;212:63-68. [ Links ]
7. Castejón OC, Belouche CR, Morett de Castejón V. Demostración histoquímica ultraestructural de carbohidratos en el trofoblasto de la placenta humana con el azul alicián. Gac Méd Caracas. 1998;106:7-12. [ Links ]
8. Castejón OC, Morett de Castejón V. Desmosomas intracitoplasmáticos en células trofoblásticas-x de la placa basal de la placenta humana a término. Gac Méd Caracas. 1988;96:385-391. [ Links ]
9. Castejón OJ, Castejón HV. Application of alcian blue and Os-Dmeda in electronhistochemical study of the cerebellar cortex. II. Os-Dmeda staining. Rev Micr Electr. 1972;1:227-238. [ Links ]
10. Hamilton WJ, Hamilton DV. Development of the human placenta. En: Philipp EH, Barnes J, Newton M, editores. Scientific foundations of obstetrics and gynaecology. 2ª edición. Chicago: A Heinemann medical books publication; 1977. [ Links ]
11. Adachi E, Takeda Y, Nakazato K, Muraoka M, Iwata M, Sasaki T, et al. Isolated collagen IV retains the potencial to form an 18-µm sided polygonal meshwork of the lamina densa. J Electron Microsc. 1997;46:233-241. [ Links ]
12. Adachi E, Hayashi T. Anchoring of epithelia to underlying connective tissue: Evidence of frayed ends of collagen fibrils directly merging with meshwork of lamina densa. J Electron Microsc. 1994;43:264-271. [ Links ]
13. Salazar H, Gonzalez-Angulo A. The fine structure of human chorionic villa and placental transfer of iron in late pregnancy. Am J Obstet Gynecol. 1967;97:851-865. [ Links ]
14. Drachenberg CB, Papadimitriou JC. Placental iron deposits: Significance in normal and abnormal pregnancies. Human Pathol. 1994;25:379-385. [ Links ]
15. Hamasaki K, Fujimoto S, Yamamoto K, Okamura Y. Iron-containing granules in the syncytiotrophoblast of the human chorionic villi. Acta Anat. 1985;121:194-196. [ Links ]
16. Castellucci M, Kosanke G, Verdenelli F, Huppertz B, Kaufmann P. Villous sprouting: Fundamentals mechanisms of human placental development. Human Reprod Update. 2000;6:485-494. [ Links ]
17. Vizza E, Goranova V, Heyn R, Correr S, Mota PM. The extracellular fibrillar matriz architecture of human placental villa at term. Ital J Anat Embryol. 2001;106:317-323. [ Links ]
18. Vizza E, Goranova V, Heyn R, Correr S, Mota PM. The extracellular matriz of normal human placental villa: A scanning electron microscopy study. Ital J Anat Embryol. 1998;103:301-307. [ Links ]
19. Risteli J, Foidart JM, Risteli L, Boniver J, Goffinet G. The basement membrane proteins laminin and type IV collagen in isolated villi in pre-eclampsia. Placenta. 1984;5:541-550. [ Links ]
20. Zerpa E, de Gavaller B E, Agüero O. La placenta en toxemia del embarazo. Rev Obstet Ginecol Venez. 1954;14:42-64. [ Links ]
21. Kingdom J, Huppertz B, Seaward G, Kaufmann P. Development of the placental villous tree and its consecuences for fetal growth. Eur J Obstet Gynecol Reprod Biol. 2000;92:35-43. [ Links ]
22. Reyna VE, Torres MM, Reyna VN, Mejías MJ. Estructura y función de la matriz extracelular de las membranas fetales humanas. Rev Obstet Ginecol Venez. 2003;63:19-30. [ Links ]
23. Iwahashi M, Muragaki Y, Ooshima A, Nakano R. Increased laminin-1 expression in hydatidiform mole. Mol Hum Reprod. 1997;3:1107-1111. [ Links ]
AGRADECIMIENTOS
Al personal técnico Sr. Raul Colina y Milagros Díaz, del Lab. de microscopia electrónica, del Instituto de Investigaciones Odontológicas, Fac. Odontología, UCV-Caracas, por brindar la infraestructura de la microscopia electrónica de transmisión (MET). Este trabajo ha sido financiado en parte por el fondo fijo institucional de la Fac. Cs de la Salud de la dirección administrativa del Núcleo Aragua, Universidad de Carabobo, para el CIADANA.
Correspondencia:
Laboratorio de microscopia electrónica. CIADANA. Fac. Cs de la Salud. Universidad de Carabobo. Núcleo Aragua. Maracay. Apdo.: 1911.
