











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.28 n.7 Caracas jul. 2003
SEASONAL DYNAMICS OF RUMINAL CRUDE PROTEIN DIGESTION OF BROWSE SPECIES FROM BAJA CALIFORNIA SUR, MEXICO
Rafael Ramírez-Orduña, Roque G. Ramírez-Lozano, Marco V. Gómez-Meza, Juan A. Armenta-Quintana, Juan M. Ramírez-Orduña, Ramón Cepeda-Palacios and Juan M. Ávila-Sandoval
Rafael Ramírez-Orduña. M.C. in Animal Nutrition. Professor, Department of Zootechnics, Universidad Autónoma de Baja California Sur (UABCS), Mexico. Address: Apartado Postal 676. La Paz Baja California Sur. 23080, México.
Roque G. Ramírez-Lozano. Doctor in Animal Sciences. Professor, School of Biology, Universidad Autónoma de Nuevo León, (UANL). Address: Apartado Postal 142, Sucursal F, San Nicolás de los Garza, N.L. 66450, México. e-mail: roqramir@fcb.uanl.mx.
Marco V. Gómez-Meza. Doctor in Statistics. Professor, School of Economics, UANL. Address: Apartado Postal 130, Monterrey, N.L. 65450, México.
Juan A. Armenta-Quintana. M.C. in Animal Nutrition. Professor, Department of Zoothecnics, UABCS, Mexico.
Juan M. Ramírez-Orduña. Doctor in Biothecnology. Professor, Department of Zoothecnics, UABCS, Mexico.
Ramón Cepeda-Palacios. Doctor in Animal Sciences. Professor, Department of Zoothecnics, UABCS, Mexico.
Juan M. Ávila-Sandoval. M.C. in Animal Nutrition. Professor, Department of Zoothecnics, UABCS, Mexico.
Summary
Ten native browse species from Baja California Sur, Mexico, were nutritionally evaluated in each season of three consecutive years measuring crude protein (CP), protein fraction in neutral detergent fiber (NDIP) and in acid detergent fiber (ADIP), and rate and extent of crude protein degradability. Five legumes and five non-legumes were collected from 16 linear 30m transects. All plants in each transect were identified and measured to estimate Shannons diversity index, species richness, aerial cover and relative frequency, dominance, density and importance. Medicago sativa L. hay was used as reference forage with high nutritional quality. To estimate the effective degradability of crude protein (EDCP), duplicate nylon bags (10x15cm; 50µm pore size) containing 4g of each plant species from each season and year were incubated in the ventral part of the rumen in four cannulated beef steers. In general, CP content, NDIP, ADIP and EDCP were significantly higher in legumes. Neutral detergent soluble CP was higher in non-legumes during spring and summer, and CP and EDCP were higher in autumn. Legume EDCP was significantly lower in summer, possibly due to reduction of available cell wall protein. Only shrubs such as Opuntia cholla, Prosopis sp. and Cercidium floridium had EDCP values comparable to or higher than alfalfa hay. Cyrtocarpa edulis had the lowest EDCP. Legume CP was more degraded in the rumen than that of non-legumes. A high quality forage period in autumn and winter, and a low quality one in spring and summer, could be distinguished.
Resumen
Diez arbustos nativos de Baja California Sur, México, fueron nutricionalmente evaluados en cada estación durante tres años consecutivos, midiendo proteína cruda (PC), fracción proteica en la fibra detergente neutro (FPDN) y en la fibra detergente ácido (FPDA), y velocidad y grado de degradabilidad de PC. Cinco leguminosas y cinco no-leguminosas fueron colectadas de 16 transectos lineales de 30m. Todas las plantas de cada transecto fueron identificadas y medidas para estimar el índice de diversidad Shannon, riqueza de las especies, cobertura aérea y frecuencia relativa, dominancia, densidad e importancia. Heno de Medicago sativa L. fue usado como forraje de referencia de alta calidad nutritiva. Para estimar la degradabilidad efectiva de PC (DEPC), se usaron por duplicado bolsas nylon (10x15cm; poro de 50¼m) conteniendo 4g de cada especie de cada estación y año, incubadas en la parte ventral del rumen de cuatro becerros de carne canulados. En general, el contenido de PC, FPDN, FPDA y DEPC fueron significativamente mayores en leguminosas. La PC soluble en la fibra detergente neutro fue mayor en las no-leguminosas en primavera y verano, y PC y DEPC fueron mayores en otoño. La DEPC en leguminosas fue significativamente menor en verano, posiblemente por reducción en la proteína disponible de la pared celular. Solo especies como Opuntia cholla, Prosopis sp. y Cercidium floridium tuvieron valores de DEPC comparables o mayores a la alfalfa. Cyrtocarpa edulis tuvo la menor DEPC. La PC de leguminosas fue más degradada en el rumen que las no-leguminosas. Se distinguen dos períodos de calidad nutritiva, uno de alta calidad en otoño e invierno y otro de baja en primavera y verano.
Resumo
Dez arbustos nativos da Baixa Califórnia Sul, México, foram nutricionalmente avaliados em cada estação durante três anos consecutivos, medindo proteína crua (PC), fração protéica na fibra detergente neutro (FPDN) e na fibra detergente ácido (FPDA), e velocidade e grau de degradabilidade de PC. Cinco leguminosas e cinco não leguminosas foram coletadas de 16 transectos lineares de 30 m. Todas as plantas de cada transecto foram identificadas e medidas para estimar o índice de diversidade Shannon, riqueza das espécies, cobertura aérea e freqüência relativa, dominância, densidade e importância. Feno de Medicago sativa L. foi usado como forragem de referência de alta qualidade nutritiva. Para estimar a degradabilidade efetiva de PC (DEPC), se usaram por duplicado sacolas nylon (10 x 15 cm; poro de 50 ìm) contendo 4 g de cada espécie de cada estação e ano, incubadas na parte ventral do rúmen de quatro bezerros de carne canulados. Em geral, o conteúdo de PC, FPDN, FPDA e DEPC foram significativamente maiores em leguminosas. A PC solúvel na fibra detergente neutro foi maior nas não leguminosas na primavera e no verão, e PC e DEPC foram maiores no outono. A DEPC em leguminosas foi significativamente menor no verão, possivelmente por redução na proteína disponível da parede celular. Somente espécies como Opuntia cholla, Prosopis sp. e Cercidium floridium tiveram valores de DEPC comparáveis ou maiores à alfafa. Cyrtocarpa edulis teve a menor DEPC. A PC de leguminosas foi mais degradada no rúmen que as não leguminosas. Distinguem-se dois períodos de qualidade nutritiva, um de alta qualidade no outono e no inverno e outro de baixa na primavera e no verão.
KEYWORDS / Native Shrubs / Sonora Desert / Crude Protein / Cell Wall Proteins / Degradability of Crude Protein /
Received: 03/20/2003. Modified: 06/13/2003. Accepted:06/19/2003
Introduction
The State of Baja California Sur, in Northern Mexico, is considered as an extremely arid zone (COTECOCA, 1975). Of the vegetation, 92% is mainly composed by shrubs 1-3m in height. Most farmers are traditional small holders. However, the use of shrubs as food resources for livestock has been mostly supported by farmers experience and the knowledge that some shrubs are palatable for animals (Arriaga and Cancino, 1992).
Many browse plants that grow in Northeastern Mexico have been used as fodder for ruminants, especially legume species (Ramírez, 1996). In other arid and semiarid regions of the world, the use of browse species has been promoted due to severe food shortages, harsh and prolonged drought periods, low animal performance, mortality and uneconomic production (Devendra, 1990). However, the low quality and seasonal nature of the forage supply, together with low intake by animals and poor digestibility of forage, are the major factors contributing to the low productivity of ruminants feeding on browse species (Remenyi and McWilliams, 1986).
In ruminants, where feed is fermented in the rumen, this fermentation has confounded the prediction of animal performance from dietary ingredients. Therefore, a mechanistic understanding of fermentation is needed for sustained development of animal production. Mechanistic models require quantitative estimates of carbohydrates and protein digestion (Satter, 1980; Russell et al., 1992; Sniffen et al, 1992).
The object of this study was to evaluate and compare seasonally, during three consecutive years, the crude protein content (CP), the CP composition of the cell wall, and the rate and extent of CP digestion in leaves of five legumes and five non-legumes growing in a shrubland from Baja California Sur, Mexico. Medicago sativa hay was included as reference forage of high nutritional quality.
Materials and Methods
Site and collection area
This study was conducted in the Palmar de Abajo ranch (800ha), located in La Paz, Baja California Sur, Mexico, at 23º38'40''N and 110º18'07''W. The climate of the region is arid with annual mean temperature of 21.2ºC. Rainfall is generally present during the summer (July, August and September) and winter (January and February) seasons. Total year rainfall during the study period was 177, 47 and 61.7mm for 1993, 1994 and 1995, respectively. The soils are mainly of alkaline, regosol, eutric and calcareous types, which are very permeable (Flores, 1998).
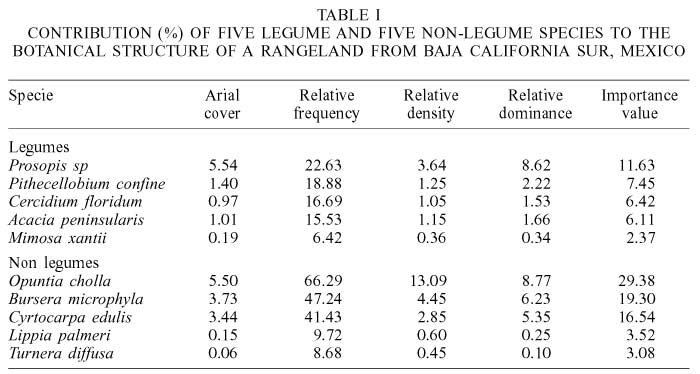
Branches from the legumes Acacia peninsularis (Brit. L. Rose) Standley; Cercidium floridium (Benth); Mimosa xantii, Gray; Pithecellobium confine (Standley) and Prosopis sp. (Torr) and the non-legumes Bursera microphylla Gray (Burceraceae); Cyrtocarpa edulis (Brand) Stand (Anacordiaceae); Lippia palmeri, S. Wats (Verbenaceae); Opuntia cholla, Weber (Cactaceae) and Turnera diffusa Wild (Turneraceae) were collected from 16 randomly located linear transects of 30m long. These browse species are reported to be consumed selectively by cattle and goats in Baja California Sur (Arriaga and Cancino, 1992). Collections were carried out during winter (February 23), spring (May 23), summer (August 23) and autumn (November 23), in 1993, 1994 and 1995. Branches of each species were bulked in triplicate samples for each season and year, and were allowed to dry under shade. Leaves were removed manually, oven dried (45ºC) for 72h, ground in a Willey mill (1mm). Alfalfa hay (Medicago sativa L.) obtained from a commercial store was considered as reference forage. Within each transect the species were identified and individuals from each specie were counted. The length of intercepts occupied by individuals touching the transect were recorded. The Shannon diversity index (Shannon, 1948) and the species richness index (Hart, 1985) were determined.
Chemical and in situ digestibility analyses
Duplicate samples were subjected to chemical analysis to determine crude protein (CP; AOAC, 1980), neutral detergent insoluble protein (NDIP) and acid detergent insoluble protein (ADIP; Van Soest et al., 1991). ADIP is considered insoluble protein (Pichard and Van Soest, 1977) because it is associated with lignin, tannin and Maillard compounds (Krishnamoorthy et al, 1982). Insoluble protein in neutral detergent but soluble in acid detergent was calculated as NDIP-ADIP (Goering and Van Soest, 1970; Krishnamoorthy et al., 1982).
The nylon bag technique was used to estimate the rate (kd, %/h) and extent of CP loss (%). Nylon bags of 10x15cm and 50µm pore size containing 4g of each plant species were incubated in the ventral part of the rumen of 4 cannulated beef steers (250 ±5.3kg body weight). During the trial steers were fed alfalfa hay ad libitum (16% CP). Duplicate nylon bags were incubated in each steer for 1, 2, 3, 4, 8, 12, 24, 48, 72 and 100 hours. Zero-time disappearance was determined from non-incubated bags. After incubation, the bags were dried at 60ºC in an oven for 48h. The in situ disappearance of crude protein (ISDCP) for every period of incubation was calculated as
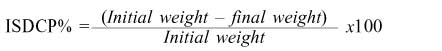
To estimate the non-linear characteristics of CP, in situ disappearance values from nylon bags, in each incubation period were used in the following equation (Ørskov and McDonald, 1979)
p = a + b (1-e-kd.t)
where p: percentage of disappearance of the CP at time t, a: soluble fraction of CP that is lost during washing, b: insoluble fraction of CP that is degraded in the rumen, kd: constant rate of disappearance of fraction b, and t: incubation time.
The non-linear parameters a, b and kd and effective degradability of the CP (EDCP= (a+b)kd/(kd+kp)[e-(kd+kp)T], where kp: outflow rate from the rumen and T: lag time (period in h that rumen microbes take to initiate fermentation), were all calculated using the Neway program (McDonald, 1981). The EDCP values of browse species and M. sativa hay were estimated using a rumen outflow rate of 2%/h.
Statistical analysis
Data of CP, CP fractions in the cell wall, degradation rate of CP and EDCP were analyzed using a general lineal models procedure (SPSS, 1999). Mean values of legumes and non-legumes species were compared using orthogonal contrasts. The season and year effect for each plant type (legumes and non-legumes) was determined by a two way analysis of variance model. Values of acid detergent lignin (ADL) and condensed tannins (CT) reported by Ramírez-Orduña et al. (2003) were used to estimate simple linear correlation coefficients between ADL, CP and EDCP, NDIP, ADIP.
Results and Discussion
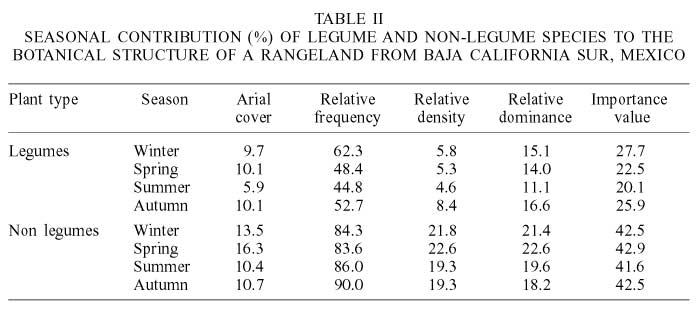
The Shannon diversity index of the study area was 1.0 %, species richness was 10.62% and the total aerial cover was 62.05%. Of the legume species, Prosopis sp. and M. xantii were the most and least important species, whereas among the non-legumes O. cholla was the most important species, and L. palmeri and T. diffusa were the least important species (Table I). In the different seasons (Table II), legumes and non-legumes covered roughly the same area (about 10%). Non-legumes contributed almost twice in importance value than legumes in all seasons because of their higher relative frequency, density and dominance; however, both plant types were similar in all seasons with respect to covered area. Non-legumes decreased all these variables from winter to autumn, whereas legumes were lower in summer and higher in autumn than other seasons.
Chemical composition
The year was the principal factor affecting EDCP, soluble and slowly degraded fractions, whereas the interaction year x plant type affected mainly the NDIP-ADIP content in plants. Moreover, plant type was the main factor that affected the CP, NDIP, ADIP and CP degradation rate (Table III).
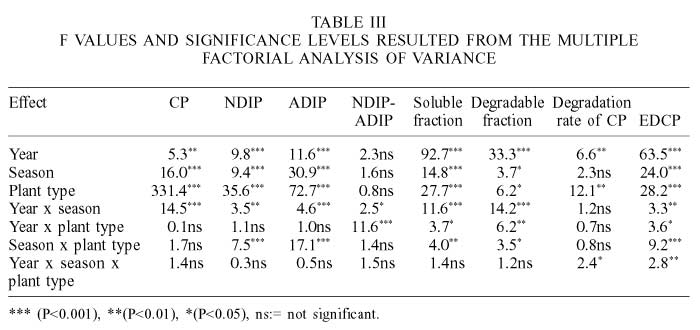
The shrub O. cholla (6.0 ±2.8 annual mean ±standard error) had the lowest CP content (Table IV) while Prosopis sp. had the highest (19.4 ±3.4), even higher than alfalfa hay (16.1 ±2.7). Legume species were significantly higher in CP (15.8 ±3.9) than non-legumes (8.7 ±3.5). In autumn, non-legumes had a higher CP content, and in legumes it was higher in winter. The CP values for alfalfa hay were comparable to those values reported for alfalfa hay mid bloom (16.5% DM) by Sniffen et al. (1992). Only non-legume species were significantly affected by the year; in years when rainfall was lower, the CP in non-legumes was also lower (7.7 ±0.4%).
According to Sniffen et al. (1992) NDIP is the fraction that includes both available and unavailable proteins linked to cell wall. Legumes (27.7 ±8.0) had lower NDIP (%CP) than non-legumes (35.1 ±13.6). Non-legumes had the higher NDIP content during spring and summer; but legumes were not significantly affected by season (Table IV). NDIP in legumes was significantly higher in 1994 than in 1993 and 1995, and in alfalfa hay it was lower (14.1 ±5.9) than in all the shrubs evaluated in this study. L. palmeri had the highest (45.4 ±15.4) and Prosopis sp. the lowest NDIP values. According to Sniffen et al. (1992) this fraction includes both buffer soluble proteins that are rapidly degraded and buffer insoluble, but neutral, detergent soluble proteins that are intermediately degraded and may be degraded in the rumen depending on the relative rates of degradation and passage. This means that the CP proportion of native legume that could be rapidly to intermediately degraded in the rumen species remains high throughout the year and is higher than in non-legumes. However, in non-legumes this fraction might be reduced in spring and summer. Also, CP in cell content of alfalfa hay (85.8 ±5.9), evaluated in this study, had a higher proportion of total CP as compared to all browse species, and was intermediate to the corresponding values (74.8 and 92.1% CP) reported for mid bloom alfalfa hay by Sniffen et al. (1992) and Elizalde et al. (1999a), respectively. Evidence from numerous feeding studies indicate that excessive ruminal protein degradation could be the most limiting nutritional factor in higher quality temperate legume forages such as alfalfa hay (Broderick, 1995).
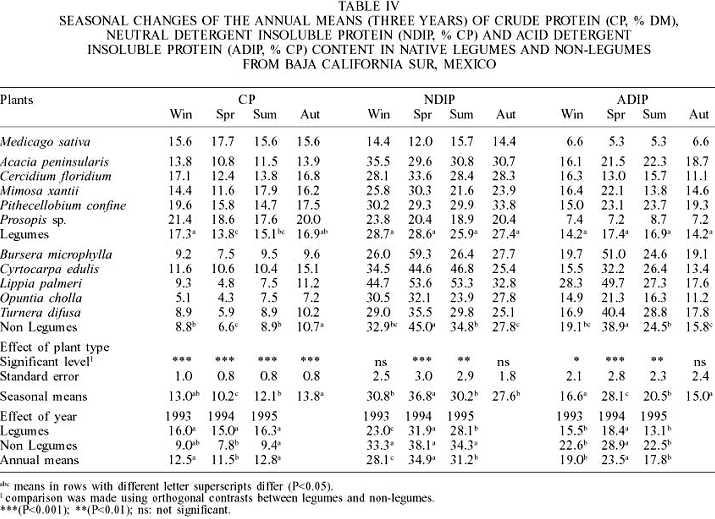
The ADIP (% CP) content in legume species (15.7 ±6.9) was significantly lower than in non-legumes (24.6 ±13.6). Moreover, ADIP in non-legumes was higher during spring and summer seasons, whereas ADIP in legumes was not affected by season. However, with exception of autumn, there were significant differences among seasons between legumes and non-legumes (Table IV). Legumes and non-legumes increased significantly (P<0.05) their ADIP in 1994. Alfalfa hay (6.0 ±1.7) had the lowest ADIP concentration. This fraction is the unavailable protein bound to lignin, tannin-protein complexes or Maillard products. Therefore, this fraction is highly resistant to ruminal and mammalian enzymes activity (Krishnamoorthy et al., 1983) and it would not provide aminoacids post ruminally (Krishnamoorthy et al., 1982).
During the summer months (Table V), legumes significantly reduced the NDIP-ADIP (%CP), which also showed a significant year effect, increasing in 1994 (13.4 ±0.9) and 1995 (15.0 ±1.0), three years of lowest precipitation. On the other hand, non-legumes were neither different among seasons nor among years. Non-legumes such as B microphylla (7.9 ±7.6), M. xantii (8.6 ±4.9), and T. diffusa (8.1 ±5.8) had comparable NDIP-ADIP contents to those of alfalfa hay (8.1 ±4.9). L. palmeri had with the highest value (18.4 ±13.2).
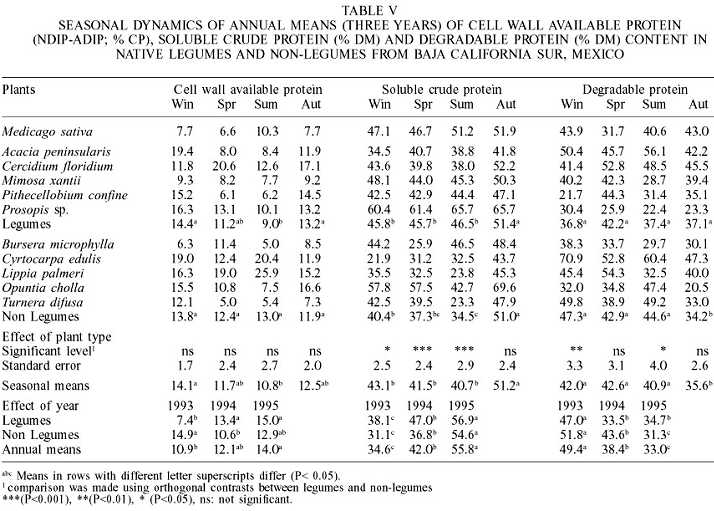
The available protein in cell wall (NDIP-ADIP) is considered as slowly degraded in the rumen, for being in the cell wall (Van Soest et al., 1981; Krishnamoorthy et al., 1983), and their extent of degradation depends on the degradation and passage rates of the fibrous fraction (Sniffen et al., 1992). It appears that if the passage rate is very fast or the degradation rate is very slow, then this fraction would escape degradation in the rumen. The NDIP-ADIP values for alfalfa hay in this study are comparable to the values reported by Elizalde et al. (1999a) and Sniffen et al. (1992), of 7.8 and 11.2% CP, respectively.
Degradability of crude
protein
In non-legumes, the soluble fraction (a) increased in autumn, after the rainy season, but was similar in the other seasons. (Table V). With exception of autumn, legumes were higher in this fraction than non-legumes. Legumes were similar to alfalfa hay, however the values obtained for the latter were higher than those values reported by Elizalde et al. (1999b) for mid vegetative (40.2%) to late flowering (41.3%) alfalfa hay, but were similar to the results of Hoffman et al. (1993) for late bud (44.8%) to midbloom (48.2%) alfalfa hay.
The slowly degraded fraction (b) was similar among seasons in legume species. In non-legumes, however, this fraction was significantly lower during winter, when the soluble fraction was highest. During winter and summer fraction (a) was higher in legumes than in non-legumes; however both plant types were affected by the year (Table V). Legumes in this study had a lower slowly degraded fraction than alfalfa hay in all seasons except in spring. Alfalfa hay values were lower than those reported by Elizalde et al. (1999b) for mid vegetative (53.5%) to late flowering (47.1%) alfalfa hay, but were similar to the results of Hoffman et al. (1993) for late bud (41.1%) to midbloom (30.8%) alfalfa hay.
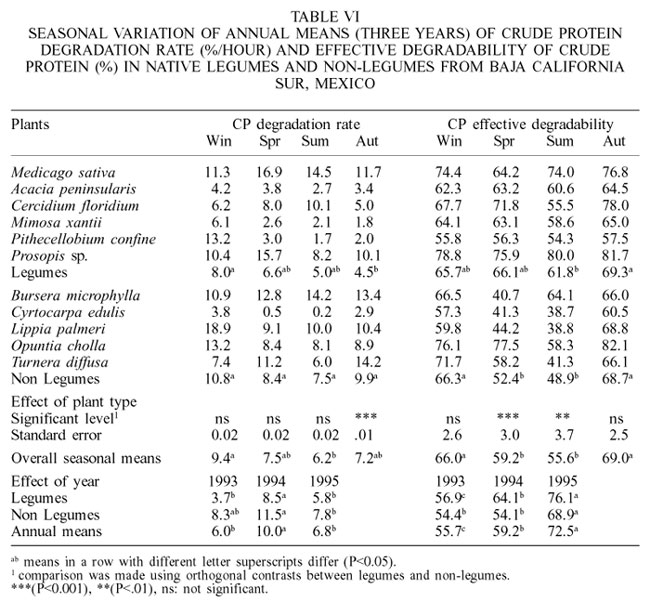
Only during the autumn months, legumes had significantly lower CP degradation rates than non-legumes species (Table VI). The CP of Prosopis sp., B. microphylla and O. cholla was rapidly degraded; however, A. peninsularis, C. edulis, M. xantii and P. confine were slowly degraded in rumen of cattle. The CP of alfalfa hay was degraded faster than all native shrubs evaluated in this study. High NDSP values (Table II) in alfalfa would have positively influenced the CP degradation rate in rumen of cattle. Conversely, low NDSP in browse plants would have reduced the digestion rate of CP.
Only during the spring and summer seasons, legumes had EDCP significantly higher values than non-legumes. Species such as O. cholla, Prosopis sp. and C. floridium had the highest EDCP values and were comparable to those in alfalfa hay. C. edulis and L. palmeri had the lowest values (Table VI). The EDCP values for alfalfa hay obtained in this study are comparable to those reported by Elizalde et al. (1999b) and Hoffman et al. (1993) of 72.9 and 72.1%, respectively. In this study, browse plants had higher EDCP values during 1995 than other years, and were also higher in autumn and winter than in other seasons.
Ramírez et al. (2000a,b) and Moya-Rodríguez et al. (2002) reported that lignin content and condensed tannins negatively influenced the EDCP in leaves of 22 browse species growing in Northeastern Mexico. They also found that during the winter months the EDCP was higher than in other seasons. They concluded that in some situations, because of warm temperatures and wet climate that occurred at the end of winter, native shrubs from Northeastern Mexico could develop new vegetative growth-producing foliage with highly soluble CP. Thus, this would explain why microbes in the rumen of sheep, in previous studies, and of cattle in this study, digested to a greater extent the CP in browse species during winter than during other seasons.
Lignin content in browse species evaluated by Ramírez-Orduña et al. (2003) was negatively correlated with EDCP in legumes (r= -0.55, P<0.001) and non-legumes (r= -0.31, P<0.001), but positively correlated with NDIP (r= 0.36 and 0.40, P<0.001, in legumes and non-legumes, respectively) and with ADIP (r= 0.61 and 0.59, P<0.001). Condensed tannins in legumes were positively correlated with NDIP (r= 0.42, P<0.001) and with ADIP (r= 0.32, P<0.001).
These results indicate that lignification in browse plants might be the most important factor in determining the protein distribution in the cell wall and, perhaps, protein fermentation. Moreover, condensed tannins in legume species might be a contributing factor in determining the protein availability of protein in the cell wall. However, it may be recognized that the structural features of lignin have a greater effect on ruminal cell wall degradation than the concentration of individual cell wall components (Jung and Deetz, 1993; Van Soest, 1994). Similarly, research on tannins in forage legumes has determined their variable effect on protein digestion (Reed, 1995; Butter et al., 1999; Mueller-Harvey, 1999). However, more research on tannin structure in relation to digestion of specific proteins is needed.
Ruminants browsing shrub species such as those evaluated in this study, possibly metabolize nitrogen in a better way than those animals feeding only on alfalfa hay, as most of the protein from the latter may be more rapidly degraded. Thus, if ruminal microbes cannot utilize all of the aminoacids and ammonia released, it is suggested that more protein would have been degraded than synthesized. Otherwise, more concentrate has to be provided to maximize capture of the degraded forage protein for ruminal protein synthesis (Broderick, 1995). Conversely, nitrogen browse plants with lower NDSP and EDCP may pass through the rumen as aminoacids to the lower gut, and eventually be absorbed in the small intestine (Van Soest, 1994).
Even though there were variations in the seasonal dynamics of protein concentration, composition and degradation rates between legume and non-legume species, the protein content and effective degradation of CP of native legumes were highly influenced by the year (Table III). Also, during the three years evaluation of nutritional dynamics of the ten most important browse species from Baja California Sur, two periods could be detected: 1) a good forage quality period during the autumn and winter seasons, and 2) a low forage quality period in the spring and summer seasons. It seems that in non-legume species the main limiting factors of forage quality were the lower CP content and cell wall available CP (NDIP-ADIP), because during the low forage quality period the low concentration of CP negatively influenced the soluble fraction as well as EDCP, and increased the insoluble fractions (NDIP and ADIP).
In this study, O. cholla, a non-legume species, with the lowest CP content in all seasons and years, had the highest EDCP and potentially degradable CP, which means that its CP is very soluble and degradable in the rumen of cattle (Table II). Other reports (Vadiveloo and Fadel, 1992; Hoffman et al., 1993) have found a wide variation in protein degradation and composition between and within plant species, and have concluded that the species and maturity interactions is wide, so that the conventional classification of forages may not be a good reflection of the compositional and degradability attributes.
Conclusions
Two periods could be detected in the three years nutritional evaluation of the ten most important native browse species from Baja California Sur: 1) a good quality forage period, represented by the nutritional values of the legume species, in the autumn and winter seasons, and 2) a low quality forage period, which can be represented by the non-legumes, in the spring and summer seasons. During the worst quality forage seasons (spring and summer), legumes almost double non-legumes in CP and degradable CP content. Moreover, legumes such as C. floridium and Prosopis sp. were the most important legumes in the area, and are largely fermented in the rumen of steers, even more so than alfalfa hay. Conversely, non-legume species had low CP content and CP fermentation values. Only the non-legumes O. cholla and C. edulis had CP contents that were highly fermented in the rumen of steers. Therefore, it is recommended that legumes be conserved and promoted in these arid regions, because of their CP and CP in the cell wall that can be largely fermented and, consequently, could improve performance of animals feeding on them and sustain a good development of range ruminants.
References
1. AOAC (1980) Official Methods of Analysis. 13th ed. Association of Official Analytical Chemists. Arlington, VA. USA. 830-850 pp. [ Links ]
2. Arriaga L Cansino J (1992) Prácticas pecuarias y caracterización de especies forrajeras en la selva Baja caducifolia. In Uso y Manejo de los Recursos Naturales en la Sierra de la Laguna, Baja California Sur. Alfredo Ortega Ed. La Paz, México. pp. 155-184. [ Links ]
3. Broderick GA (1995) Desirable characteristics of forage legumes for improving protein utilization in ruminants. J. Anim. Sci. 73: 2760-2767. [ Links ]
4. Butter NL, Dawson JM, Buttery PJ (1999) Effects of dietary tannins on ruminants. In Caygill JC, Mueller-Harvey I (Eds.) Secondary Plant Products Anti nutritional and beneficial actions in animal feeding. Nottingham University Press. Nottingham, UK. pp. 51-70. [ Links ]
5. COTECOCA (1975) Coeficientes de agostadero de la República Mexicana: Estado de Baja California, Sur. Secretaría de Agricultura y Ganadería. México. p. 67. [ Links ]
6. Devendra C (1990) The use of shrubs and tree fodders by ruminants. In Devendra C (Ed.) Shrubs and Tree Fodders for Farm Animals. Proceedings of a workshop in Denpasar Indonesia. International Development Research Center. Ottawa, Canada. pp 51-70. [ Links ]
7. Elizalde JC, Merchen NR, Faulkner DB (1999a) Fractionation of fiber and crude protein in fresh forages during the spring growth. J. Anim. Sci. 77: 476-481. [ Links ]
8. Elizalde JC, Merchen NR, Faulkner DB (1999b) In situ dry matter and crude protein degradation of fresh forages during the spring growth. J. Dairy Sci. 82: 1978-1983. [ Links ]
9. Flores EZ (1998) Geosudcalifornia Geografía Agua y Ciclones. Universidad Autónoma de Baja California Sur. La Paz, Baja California Sur, México. pp. 155-208. [ Links ]
10. Goering HK, Van Soest PJ (1970) Forage fiber analyses (apparatus, reagents procedures and some applications). Agric Handbook Nº379. ARS/USDA. Washington DC, USA. pp. 29-33. [ Links ]
11. Hart RD (1985) Conceptos básicos sobre agro ecosistemas. Serie de Materiales de Enseñanza Nº 1. CATIE. Turrialba, Costa Rica. 160 pp. [ Links ]
12. Hoffman PC, Sievert SJ, Shaver RD, Welch DA, Combs DK (1993) In situ dry matter protein and fiber degradation of perennial forages. J. Dairy Sci. 76: 2632 [ Links ]
13. Jung HG, Deetz DA (1993) Cell wall lignification and degradability. In Jung HG, Buxton DR, Hatfield RD, Ralph J (Eds.) Forage Cell Wall Structure and Digestibility. ASA-CSSA-SSSA. Madison, WI, USA. pp 315-339. [ Links ]
14. Krishnamoorthy UC, Muscato TV, Sniffen CJ, Van Soest PJ (1982) Nitrogen fractions in selected feedstuffs. J. Dairy Sci. 65: 217-221. [ Links ]
15. Krishnamoorthy UC, Sniffen CJ, Stern MD, Van Soest PJ (1983) Evaluation of a mathematical model of digesta and in-vitro simulation of rumen proteolysis to estimate the rumen undergraded nitrogen content of feedstuffs. Br. J. Nutr. 50: 555-560. [ Links ]
16. McDonald I (1981) A revised model for estimation of protein degradability in the rumen. J. Agric. Sci. 96: 251-255. [ Links ]
17. Moya-Rodríguez JG, Ramírez RG, Foroughbakhch R (2003) Nutritional value and effective degradability of crude protein in browse species from northeastern Mexico. J. Appl. Anim. Res. In Press. [ Links ]
18. Mueller-Harvey I (1999) Tannins: Their nature and biological significance. In Caygill JC, Mueller-Harvey I (Eds.) Secondary Plant Products, Antinutritional and beneficial actions in animal feeding Nottingham University Press. Nottingham, UK. pp 17-40. [ Links ]
19. Ørskov ER, McDonald I (1979) The estimation of protein degradability in the rumen from incubation measurements weighed according to rate of passage. J. Agric. Sci. (Camb). 92: 499-504. [ Links ]
20. Pichard DG, Van Soest PJ (1977) Protein solubility of ruminant feeds. Proc. Cornell Nutr. Conf. Ithaca, NY, USA. p. 91. [ Links ]
21. Ramírez RG (1996) Feed value of browse. VI International Conference on Goats International Academic Publishers. Beijing, China. pp 510-527. [ Links ]
22. Ramírez-Orduña R, Ramírez RG, Gómez-Meza MV, Armenta-Quintana JA, Ramírez-Orduña JM, Cepeda-Palacios R, Ávila-Sandoval JM (2003) Seasonal Dynamics of Organic Matter Digestion in browse species from Baja California Sur Mexico. J. Appl. Anim. Res. In press. [ Links ]
23. Ramírez RG, Neira-Morales RR, Torres-Noriega JA (2000a) Digestión ruminal de la proteína de siete arbustos nativos del nordeste de México. Int. J. Exp. Bot. (FYTON) 67: 29-35.
24. Ramírez RG, Neira-Morales RR, Torres-Noriega JA, Mercado-Santos AC (2000b) Seasonal variation of chemical composition and crude protein digestibility in seven shrubs of NE Mexico. Int. J. Exp. Bot. (FYTON) 68: 77-82.
25. Reed JD (1995) Nutritional toxicology of tannins and related polyphenols in forage legumes J. Anim. Sci. 73: 1516-1520. [ Links ]
26. Remenyi JV, McWilliams JR (1986) Ruminant production trends in Southeast Asia and the South Pacific, and the need for forages. In Blair GJ, Ivory DA, Evans TR (Eds.) Forages in Southeast Asian and South Pacific Agriculture. Australian Center for International Agricultural Research. Canberra, Australia. pp 1-6. [ Links ]
27. Russell JB, O'Connor JD, Fox DG, Van Soest PJ, Sniffen CJ (1992) A net carbohydrate and protein system for evaluating cattle diets: I. Ruminal fermentation. J. Anim. Sci. 70: 3551-3557. [ Links ]
28. Satter L (1980) A Metabolizable Protein System Keyed to Ruminal Ammonia Concentration The Wisconsin System In Owen FN (Ed.) Protein Requirements for Cattle. Oklahoma State Univ. USA. p. 245. [ Links ]
29. Shannon CE (1948) A mathematical theory of comunication. Bell System Tech. J. 27: 379-423. [ Links ]
30. Sniffen CJ, OConnor JD, Van Soest PJ, Fox DG, Russell JB (1992) A net carbohydrate and protein system for evaluating cattle diets: II Carbohydrate and protein availability. J. Anim. Sci. 70: 3562-3568. [ Links ]
31. SPSS (1999) Statistical package for Social Sciences. Users Manual (Release 9.0). pp. 120-133. [ Links ]
32. Vadiveloo J, Fadel JG (1992) Compositional analyses and rumen degradability of selected tropical feeds. Anim. Feed Sci. Technol. 37: 265-270. [ Links ]
33. Van Soest PJ (1994) Nutritional ecology of the ruminant. 2nd ed. Cornell Univ Press. Ithaca, NY. USA. pp 256-290. [ Links ]
34. Van Soest PJ, Sniffen CJ, Mertens DR, Fox DG, Robinson PH, Krishnamoorthy UC (1981) A net protein system for cattle: The rumen submodel for nitrogen. In Owens FN (Ed.) Protein Requirements for Cattle. Div. of Agriculture. Oklahoma State Univ. USA. p. 265. [ Links ]
35. Van Soest PJ, Robertson JB, Lewis BA (1991) Methods for dietary fiber neutral detergent fiber and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74: 3583-3590. [ Links ]
