











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.29 n.4 Caracas abr. 2004
SPATIAL PARTITIONING OF FISHES IN THE RÍO PARAGUAY, PARAGUAY
BARRY CHERNOFF, PHILIP W. WILLINK and ANTONIO MACHADO-ALLISON
Barry Chernoff. Ph.D., University of Michigan, USA. Professor, Departments of Biology and Earth and Environmental Sciences, Wesleyan University, USA. Address: Wesleyan University, Middletown, CT 06459, USA. e-mail: bchernoff@wesleyan.edu
Philip W. Willink. Ph.D., University of Michigan, USA. Assistant Collection Manager, Field Museum of Natural History, Chicago, USA. Address: Departament of Zoology, Field Museum of Natural History, 1400 S. Lakeshore Drive, Chicago, IL 60605, USA. e-mail: pwillink@fmnh.org
Antonio Machado-Allison. Biologist, Universidad Central de Venezuela (UCV). Ph.D., The George Washington University, USA. Professor, UCV. Address: Laboratorio de Sistemática de Peces, Instituto de Zoología Tropical, UCV. Apartado 47058. Caracas 1041A, Venezuela. e-mail: amachado@strix.ciens.ucv.ve
Abstract:
The 173 species of freshwater fishes collected during the AquaRAP expedition to the Río Paraguay were analyzed to determine if distributional patterns existed within the region. The region was divided into five subregions and into eight macrohabitats. Two null hypotheses were tested and rejected: i) that fishes are randomly distributed with respect to subregion, and ii) that fishes are randomly distributed with respect to macrohabitat. The results show a strong subregional effect, such that two distributional elements were present. One consists of an association between the Río Paraguay and Río Negro subregions. The other consists of Río Apa and Riacho La Paz subregions. The analysis of distributions with respect to macrohabitats also reveals two components. The first relates the beach and main channel faunas to the macrohabitats that are inundated during seasonal flooding, such as flooded forests and lagoons. The assemblage associated with the flooding cycle accounts for more than 75% of the fishes collected. The second component comprises habitats found within Río Apa and Riacho La Paz (e.g. clear water, rapids, etc.). This component has a relatively sharp boundary with respect to Río Paraguay, with more than a 50% turnover in fauna. These results show that core conservation areas must be set up within situations in which the seasonal flooding cycle is unimpeded and the area of inundation is relatively unmodified. Also, Río Apa and Riacho La Paz represent a highly threatened area because of high rates of land conversion, aridity, and the fact that much of the fauna cannot be recolonized from nearby water sources.
KEYWORDS: Conservation; Distribution; Fishes; Freshwater; Paraguay
LA REPARTICIÓN ESPACIAL DE LOS PECES EN EL RÍO PARAGUAY, PARAGUAY
Resumen:
Las 173 especies de peces dulceacuícolas colectados durante la expedición AquaRAP al Río Paraguay fueron analizadas para determinar si existen patrones de distribución dentro de la región. La región fue dividida en cinco subregiones y en ocho macrohábitats. Dos hipótesis nulas fueron examinadas y rechazadas: i) que los peces están distribuidos al azar con respecto a las subregiones; y, ii) que los peces están distribuidos al azar con respecto a los macrohábitats. Los resultados muestran un fuerte efecto subregional, tal que dos elementos distribucionales estaban presentes. Uno consiste de una asociación entre las subregiones del Río Paraguay y el Río Negro. El otro consiste de una asociación entre las subregiones de Río Apa y Riacho La Paz. El análisis de distribuciones con respecto a los macrohábitats también revela dos componentes. El primero relaciona las faunas presentes en la playa y el canal principal con los macrohábitats que son inundados periódicamente durante la estación lluviosa, tales como los bosques inundables y lagunas. El ensamblaje asociado con los ciclos de inundación corresponde a más del 75% de los peces colectados. El segundo componente comprende hábitats encontrados dentro del Río Apa y el Riacho La Paz (p.e. aguas claras, rápidos, etc.). Este componente posee un límite relativamente marcado con respecto al Río Paraguay con más del 50% de recambio en fauna. Estos resultados muestran que áreas básicas para conservación deben incluir situaciones en que el ciclo estacional de inundación ocurra y el área de inundación esté relativamente sin modificaciones. También, Río Apa y Riacho La Paz representan áreas con alta amenaza debido a altas tasas de conversión de la tierra, aridez y el hecho que mucha de su fauna no puede ser recolonizada a partir de fuentes ícticas de agua cercanas.
A REPARTIÇÂO ESPACIAL DOS PEIXES NO RIO PARAGUAI, PARAGUAI
Resumo:
As 173 espécies de peixes doceaqüícolas recolhidos durante a expedição AquaRAP ao Rio Paraguai foram analisadas para determinar se existem padrões de distribuição dentro da região. A região foi dividida em cinco sub-regiões e em oito macro-hábitats. Duas hipóteses nulas foram examinadas e recusadas: i) que os peixes estão distribuídos ao azar com respeito às subregiões; e, ii) que os peixes estão distribuídos ao azar com respeito aos macro-hábitats. Os resultados mostram um forte efeito sub-regional, tanto que dois elementos distribuicionais estavam presentes. Um consiste de uma associação entre as sub-regiões do Rio Paraguai e o Rio Negro. O outro consiste de uma associação entre as sub-regiões do Rio Apa e Riacho La Paz. A análise de distribuições com respeito aos macrohábitats também revela dois componentes. O primeiro relaciona as faunas presentes na praia e o canal principal com os macro-hábitats que são inundados periodicamente durante a estação de chuvas, tais como os bosques inundáveis e lagos. A ensamblagem associada com os ciclos de inundação corresponde a mais de 75% dos peixes recolhidos. O segundo componente compreende hábitats encontrados dentro do Rio Apa e o Riacho La Paz (p.e. águas claras, rápidos, etc.). Este componente possui um limite relativamente marcado com respeito ao Rio Paraguai com mais do 50% de recambio em fauna.
Received: 04/10/2003. Modified: 02/19/2004. Accepted: 02/24/2004.
The fishes of the Río Paraguay comprise a rich assemblage with more than 250 species documented from the entire river basin (Bonetto, 1986; Lowe- McConnell, 1987). Toledo-Piza et al. (2001) collected 173 species in the Río Paraguay between Concepción and the Río Negro. Based on observations made during a conservation survey (AquaRAP), the area of collections was divided into five subregions: Río Negro, Upper Río Paraguay, Lower Río Paraguay, Río Apa and Riacho La Paz (Figure 1). Furthermore, a number of macrohabitats (e.g. beaches, flooded forests, rapids, etc.) were recognized within which the fishes were captured. There are many trenchant threats to the aquatic ecosystems of the Río Paraguay (e.g. Hidrovia, land conversion, etc.) and this paper will explore what the consequences of the threats might be, given the distributions of fishes with respect to the subregions and macrohabitats.

A pattern of heterogeneous distribution by the fishes within the Río Paraguay with respect to subregions and macrohabitats would have important ramifications for conservation recommendations. For example, if the fauna were homogeneously distributed, then a core conservation area could be established that might effectively protect the vast majority of the species. However, as the distribution of the species either among subregions or among macrohabitats becomes increasingly distinct and patchy, then a single core area, apart from the entire region, may not provide the desired level of protection. Chernoff et al. (1999) demonstrated how it is possible to use information on the relative heterogeneity of distributions among sub-regions or among macrohabitats to predict possible faunal changes in response to specific environmental threats and that such analyses be carried out within the framework of a rapid assessment program.
This paper will test two null hypotheses that are critical to the conservation of freshwater fishes of the Río Paraguay: that the fishes are randomly distributed among i) five subregions; and ii) eight macrohabitats. To test these hypotheses we add to the approaches of Chernoff et al. (1999) and use methods developed by Atmar and Patterson (1993). Because the two null hypotheses are rejected, we then estimate what changes in the ichthyofauna might take place given particular environmental threats.
Methods
Regions
The collecting expedition took place in Sept 4 to 18, 1997, when 110 stations were sampled in the Río Paraguay basin between Río Negro to the north and Río Aquidabán to the south (Figure 1). The survey included portions of Río Apa and Riacho La Paz, independent tributaries of Río Paraguay. The area was divided into five sub-regions (Figure 1) with the number of sampling stations as follows: i) Río Negro, 8 stations; ii) Upper Río Paraguay, upstream from Cerritos Pão de Açúcar at 21º26'S, 57º55'W to just above the mouth in Río Negro, 57 stations; iii) Lower Río Paraguay, downstream from Cerritos Pão de Açúcar to the mouth of the Río Aquidabán at 23º04'S, 57º32'W, 26 stations; iv) Río Apa, 15 stations; and v) Riacho La Paz, 4 stations. The exact localties with their descriptions are given in map 2 and appendix 12 in Chernoff et al. (2001).
The Río Paraguay in northern Paraguay is a large, meandering river flowing across relatively flat terrain. Water level fluctuations and flat terrain result in widespread flooding and frequent changes in the rivers course. Oxbow lakes collecand abandoned channels are common. Forests and palm stands that line the shores are flooded routinely for several months each year. Large masses of floating vegetation (e.g. Eichhornia sp.) that form floating meadows are common. Rock outcroppings near Cerritos Pão de Açúcar constrict the free flow of water downstream; this point was used to separate the upper from the lower sections of Río Paraguay. The Río Negro is a large black water tributary of the Río Paraguay. In the lower reaches where the collections were made, there are inundated forests and palm stands as well as floating meadows of Eichornia.
The Río Apa differs from Río Paraguay in that it is a smaller river through terra firme with cascades and waterfalls. The bottom is predominantly sand and the banks are high and well defined. Accumulations of tree trunks and branches are common. Habitat diversity in Río Apa is lower than in Río Paraguay. Water levels can change rapidly in response to rains; Río Apa acts as a storm water river. Riacho La Paz passes through terra firme and is a riffle-and-pool headwater stream; the bottom is sand and rock. Water levels change rapidly in response to rains. Habitat diversity is low.
Macrohabitats
Nine macrohabitats were collected in the upper Río Paraguay basin: Río Paraguay beach, Río Apa beach, backwater, flooded forest, floating vegetation, lagoon, main channel, clear water and rapids. Because only three species (Bryconamericus sp., Holoshesthes pequira and Imparfinis minutus) were collected in a single sample from a rapids, this macrohabitat was not analyzed further. Therefore, eight macrohabitats were included in the analyses.
Data and Statistics
The occurrence of any of the 173 species was recorded in species by subregion and species by macrohabitat matrices (appendices 12 and 13 in Chernoff et al., 2001; available at www. biodiversityscience.org/xp/CABS/research/ rap/aboutrap.xml) from which hypotheses were tested. The presence of a species was scored as 1 and its absence as 0 in the matrices with species on the rows and subregion or macrohabitat on the columns. Although all of the individuals captured at each field sampling station were counted, no calculations of abundance were made and the number of times that a species was collected in a subregion, in a macrohabitat or in a type of water was not used.
Given point source data obtained in rapid assessments only the presence of a species should be used as information. The absence of a species is ambiguous (Chernoff et al., 1999), signifying either that i) it was not collected in a subregion or macrohabitat; or ii) the species does not live in that subregion or macrohabitat. Given the nature of rapid assessments the two alternatives cannot be distinguished. The methods below use only the shared presence of a species as information.
The results of the methods that follow are potentially influenced by relative effort. Effort refers to the number of field stations (collections) that were made. This is important because Toledo- Piza et al. (2001; see fig. 6.1) demonstrated that the species accumulation curve was not asymptotic and the number of species continued to rise with the number of collections. To determine if there is an effect of sampling effort, the number of species found in each subregion or macrohabitat was regressed onto the number of collection stations. Because the independent variable, number of collection stations, was determined without error, type I least squares regressions (Sokal and Rohlf, 1995) were used. The regressions were constrained to pass through their origins (0 collections, 0 species) because they are nontrivial.
Chernoff et al. (1999) selected Simpsons Index of Similarity as the most consistent with data collected during rapid inventories or with point source data, such as for fossils, because only the joint presence contributes to the estimate of similarty. The following table is used to calculate the similarity between two lists or samples of species (or binary data)

where a: number of positive matches or species present in both samples, b: number of species present in sample 2 and absent from sample 1, c: the converse of b, d: number of negative matches or species absent from both localities. Simpsons Index of Similarity is,
S = a/n (1)
where n: number of species (a+b+c+d) in the samples, which is the same for both samples.
Ss = a/ns (2)
where ns: number of species present in the smaller of two lists. A robust estimate of similarity, S's, is calculated as follows:
1) create the ith subset of the larger sample by rarefying the number of species in the larger sample to equal the number of species in the smaller sample, ns,
2) calculate Ssi for the ith subset with equation 2,
3) iterate steps 1 and 2, 200 times,
4) calculate a mean similarity, S's, from the 200 similarities, Ssis, and enter into a table of similarities (e.g. Table I),
5) repeat steps 1-4 for all pairs subregions or macrohabitats. It is critical to determine if the mean similarities, S's, among the subregions or macrohabitats differ significantly from random lists of species. To do this, simulate across the range of the number of species, ns, found in the subregions or macrohabitats, as follows:
6) select ns that corresponds to the number found in the macrohabitat or subregion with the fewest species,
7) create two random lists of species of size ns, each drawn from the set of all species; referred to as the random pair,
8) calculate the Simpsons Similarity of the random pair,
9) iterate steps 7 and 8, 200 times,
10) calculate the mean random similarity, S*n, where n refers to the number of species present in a subregion or macrohabitat,
11) calculate the standard deviation, D*n, of Simpsons Similarity among the random pairs,
12) increment ns by 10 species and iterate steps 7-12 until ns exceeds the number of species in the richest subregion or macrohabitat.

These 200 random similarities approximate a normal distribution (Figure 2) Random similarity distributions were generated at intervals of 10 species in order to estimate S*n and its standard deviation for samples containing 20 to 120 species. This range of random list-sizes encompasses the actual number of species observed in subregions and in macrohabitats. The means and standard deviations are plotted against number of species found in a subregion or macrohabitat (Figure 3). As the number of species present in a sample increases the observed similarity due to random effects also increases but the variance decreases.


To complete the analysis the observed mean similarity, S's (step 1) is compared to mean random similarity, S*n (step 10), and its standard deviation, D*n (step 11). Using a 2-tailed parametric approach, the probability of obtaining S's at random is calculated from the number of standard deviations that S's is either above or below S*n. This probability is obtained by interpolation of the values presented in standard statistical tables (table A in Rohlf and Sokal, 1995). The criterion for rejection of a null hypothesis that S's= S*n was P=0.01. The significance of the probability values was adjusted with the sequential Bonferroni technique (Rice, 1989) because each sample is involved in multiple comparisons. The sequential Bonferroni procedure is conservative, making it harder to reject a null hypothesis.
If S's is found to be significantly different from S*n, then it is concluded that the observed similarity is not due to random effects. If the significant difference is on the lower tail of the distribution, then this indicates boundaries of faunal turnover, habitat partitioning, etc. If the significant difference is on the upper tail then it is concluded that the similarity is indicative of biological dependence or correlation, such as nested subsets, clinal variation, one population acting as the source population for another, etc.
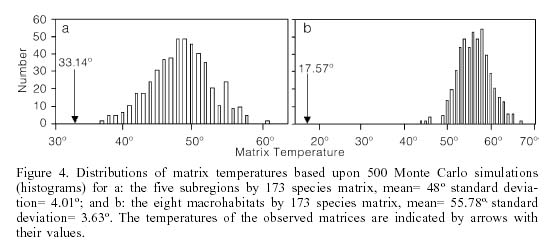
If similarities that are not random were discovered, then it was investigated whether the pattern of species presence is also non-random. The measure of matrix disorder as proposed by Atmar and Patterson (1993) calculates the matrix entropy as measured by temperature. Temperature measures the deviation from complete order (0º) to complete disorder (100º). To test whether the temperature could be obtained due to random effects, 500 Monte Carlo simulations of randomly determined matrices of the same geometry were calculated. Software to calculate matrix disorder is available from the authors at www.fieldmuseum.org
Branching diagrams are constructed as follows. For the subregional data, a Gabriel Network (Gabriel and Sokal, 1969) is used to represent the hydrological pattern of connectivity upon which the pattern of similarities are displayed. Dendrograms are constructed for the sub-regional and macrohabitat data sets using Camin-Sokal parsimony (CSp). CSp does not allow reversals, only independent acquisitions. An artificial outgroup was constructed of all zeros so that clustering summarizes similarities due to the shared presence of species, not the shared absence of species. PAUP* 4.0b was used to calculate CSp cluster analyses. A non-hierarchical, overlapping dendrogram (see Sneath and Sokal, 1973) was constructed to summarize the largest similarites for each macrohabitat.
A simulation procedure was used to evaluate the large number of unique species that were found in the Upper Río Paraguay subregion. Because this subregion had the largest number of species it should be expected to contain the largest number of unique species. This proposition became the null hypothesis and was tested as follows. A set of five, independent, random samples were drawn from the total list of all species such that one of the five contained the number of species observed in one of the subregions: 105, 93, 85, 35 and 41 species. This procedure was repeated 200 times to yield a distribution of the number of unique species expected at random. The number of unique species observed in the Upper Río Paraguay subregion was compared to the mean using standard deviations of the distribution based upon random processes.
Results
Effort
Confounding effects of differential effort were not found to exist in the data sets for subregions or macrohabitats. For subregions, the slope of the regression line was not significantly different from zero (F=5.26, df=1&3, P<0.11). The number of stations did not significantly affect the number of species captured in a subregion. For example, 57 stations were collected in the Upper Río Paraguay subregion from which 105 species were recorded. However, roughly half the effort in the Lower Río Paraguay subregion (26 stations) resulted in almost the same number of species (93). In the Apa subregion 85 species were collected from only 15 stations. The situation concerning the 10 macrohabitats is identical to that of subregions: the slope was not significantly different from zero (F=3.72, df=1&7, P>0.1).
Subregions
The fishes were found to be neither evenly nor randomly distributed among the five subregions. The Upper Río Paraguay subregion had the highest species richness; 60.7% of all species were collected there. The Lower Río Paraguay and Río Apa subregions, however, were not appreciably less diverse (Table I). These three regions account for 94.2% of all the species collected. Río Negro and Riacho La Paz accounted for only 10 species not found in the other subregions (Table I). Despite the degree of apparent overlap among subregions, a relatively small percentage of all the species collected were found in all subregions. Out of the 173 species collected, only nine species (5.2%) were captured in each of the subregions: Aphyocharax anisitsi, Characidium cf. fasciatum, Holoshesthes pequira, Moenkhausia dichroura, Odontostilbe paraguayensis, Psellogrammus kennedyi, Pyrrhulina australis, Steindachnerina brevipinna and Crenicichla lepidota.
The number of species found uniquely in the Upper and Lower Río Paraguay and Río Apa subregions comprise 19.4-28.2% of their species richnesses (Table I). A relatively high percentage would be expected from the Río Apa subregion because it passes through terra firme soils and has different habitats than are found in the Río Paraguay subregions. While some of the fishes collected only in Río Apa, such as Pimelodella laticeps and Thoracocharax stellatus are probably transients, others such as Imparfinis minutus, Jupiaba acanthogaster and Xenurobrycon macropus are not.
The interpretation of the high percentage of unique species in several of the subregions requires careful consideration (Table I). The set of random samples of the size of the Upper Río Paraguay subregion has a mean of 15.7 ±3.3 unique species. The observed value of 27 unique species is 3.4 standard deviations above the mean (P<0.0002), significantly more than expected by random processes. The fact that most of the species that were collected only in the Upper Río Paraguay are known to have much larger distributions (e.g. in the Pantanal of Brazil; Britski et al., 1999; Willink et al., 2000) may indicate that this region is not necessarily unique but rather influenced by the greater diversity found in the northern portions of the basin, including the Pantanal (Bonetto, 1986).
The numbers of unique species (Table I) found in Río Apa and in the Lower Río Paraguay are also greater than expected at random. The list of unique species for the Lower Río Paraguay contains many species that are clearly artifacts of point source data, such as Potamotrygon motoro, Serrasalmus elongatus, S. spilopleura, Ageneiosus brevifilis, Pimelodus blochii, P. maculatus, Pinirampus pirinampu and Sorubim lima. If we subtract minimally these 7 species from the list, then the number of unique species within the Lower Río Paraguay is not significantly different (P<0.22) from that expected at random. Though it will be concluded (see below) that there is an important non-random relationship between the Río Paraguay and Río Apa subregions, the differences due to the number of unique species reflect differences in macrohabitats that were sampled within each of these two subregions.
Seven of the 10 similarities coefficients (Table I) were significantly different from random similarities (P<.0001). Most of the significant similarities are above the mean random similarity, S* s, indicating that there is biological dependence among the subregions (Chernoff et al., 1999). The biological dependence may take the form of nested subsets or clinal turnover, etc. (see Leibold and Mikkelson, 2002), that may be due to relative area, hydrological connections or macrohabitats. For example, 29 of the 41 species collected in the Río Negro were found in the Upper Río Paraguay (S's >80%; Table I). Because the collections were taken from the lower reaches of Río Negro, in a relatively restricted area, this may comprise a nested subset of the larger Upper Río Paraguay subregion. In another example, more than 85% of the fishes collected in Riacho La Paz were also present in Río Apa (Table I). In this instance, however, the two rivers are not confluent but rather they are adjacent drainages on the east side of Río Paraguay. A nested relationship between the two would be due to similar macrohabitats found in these tributaries.
Entropy statistics (Atmar and Patterson, 1993) were used to test if the similarity matrix (Table I) was more structured (less disorder) than predicted at random. The subregional matrix had a temperature of 33.14º, a value significantly cooler (has more structure; P=1.07x10-4) than that predicted from 500 Monte Carlo simulations (Figure 4a). Thus, the occurrence of fish species in the subregions is significantly patterned despite the idiosyncratic occurrences of the unique species.
The Camin-Sokal parsimony analysis from the matrix of species distributions (appendix 13 in Chernoff et al., 2001) produces two clusters of subregions (Figure 5): i) Río Negro-Upper Río Paraguay- Lower Río Paraguay, and ii) Río Apa-Riacho La Paz. The groups result not only from high similarities but particularly from the shared presence of species that were not found elsewhere. For example, Riacho La Paz and Río Apa share 6 species uniquely (14.6% of the Riacho La Paz collection) of which only the first would seem to be artifactual: Astyanax fasciatus, Astyanax lineatus, Brachychalcinus retrospina, Bryconamericus sp. 1, Otocinclus mariae, and Piabarchus cf. analis.

The groups represent larger, riverine fauna and a terra firme tributary fauna, respectively (Figure 5). The similarities between the two clusters are largely not different from random expectations (Table I). The non-random artifacsimilarity between Riacho La Paz and the Lower Río Paraguay is most likely due to the fact that samples were not taken very far above the mouth of the former. Plotting the subregional similarities on the Gabriel network (Figure 6) disputes the hypothesis that faunal change is a function of distance. For example, the Lower and Upper Río Paraguay and Río Negro subregions have almost identical similarities to Río Apa despite the differences in distance (Figure 6, Table I).

Macrohabitat
The fishes were found to be neither evenly nor randomly distributed among the five subregions. The patterns discovered are of large faunal overlaps and of dependence. The beaches along Río Paraguay were the richest habitats with more than 1.6 times the number of species found in the next richest macrohabitat, flooded forests (Table II). Clear water and Río Apa beach macrohabitats join the flooded forests in the second tier of species richness.
The overall results indicate that species are widely dispersed among many of the macrohabitat types. Few species were found exclusively in two macrohabitats and just two pairs of macrohabitats shared 5 or more species exclusively (Table II). Furthermore, only four of the eight macrohabitats: Río Paraguay beaches, Río Apa beaches, main channel and clear waters, had more than 10 unique species.
The results indicate that there were also limitations on species distributions. There were no species that were found in more than six macrohabitats. Fifteen and 35 species were found in six and five macrohabitats, respectively. The number of fish species found in four or three macrohabitats continues to increase before dropping off at two.
Twenty of the 28 comparisons among macrohabitats were significantly different (P<.0001) from random similarity on the upper tail of the distribution (Table II). These results indicate that there are dependence relations among the macrohabitats. Of the eight similarities that fall within expectations of random similarity, six involve the main channel (Table II). Only the similarity of the main channel to the Río Paraguay beaches was significantly different from random. The main channel habitat extends into deeper water from beaches and shores. Many of the fish species found in the main channel were larger (e.g. the stingray, Potamotrygon motoro, the dorado, Salminus maxillosus, or the sorubí, Sorubim lima) and were not taken in other habitats. Given the adjacent spatial relationship of the beach and the main channel within Río Paraguay, it is not surprising that a relatively high percentage of the species collected in the main channel should also be collected along the beaches.

The observed temperature of the species by macrohabitat matrix, 17.6º, was more than 10.5 standard deviations below the mean of 500 Monte Carlo simulations (P=9.3x10-26, Figure 4b). The distribution of species among macrohabitats is significantly more orderly than would be expected at random. The idiosyncratic distribution of a number of species is not sufficient to disrupt an overall pattern of order.
The CSp analysis (Figure 7) emphasizes the dissimilarities of the main channel macrohabitat from the others. Among the remaining macrohabitats two clusters are present: i) those primarily associated with the mainstem of Río Paraguay; and ii) a cluster containing the Río Apa beach and clear water macrohabitats. The larger cluster is interesting because the macrohabitats cluster in relation to the flood cycle, from inland habitats to the shores of the Río Paraguay.

The group containing the Río Apa beach and clear water macrohabitats emphasizes the 5 fish species shared uniquely by them rather than the fact that their highest overall similarities are to the beaches of Río Paraguay. These species (e.g. Brachychalcinus retrospina, Corydoras aeneus, and Pimelodella mucosa) prefer smaller habitats with sandy bottoms, and less than turbid waters. All of the clear water habitats were either tributaries to Río Apa or in Riacho La Paz.
The non-hierarchical diagram displays the largest number of significant similarities among macrohabitats (Figure 8). This analysis places the Río Paraguay beach macrohabitat at the center of a concentric arrangement of habitats. In all cases the highest similarities are to the Río Paraguay beaches and then, secondarily or thirdly, to other macrohabitats. For example, backwaters join the flooded forests secondarily, which then joins equally but independently to floating vegetation and to lagoons. This indicates an apparent use of the environments by fish species at two levels. At the first level, Río Paraguay serves as a source or a sink for populations moving into or out of the other macrohabitats. At the second level, fishes are moving among several interior macrohabitats (e.g. from flooded forests into lagoons) that are interconnected during the flooding cycles.

The species accumulation curve for macrohabitats (Figure 9) was modeled with a logarithmic regression, yielding
F = 65.67×H0.216 + ε
where F: cumulative frequency of species, H: number of macrohabitats, and å: error term. The regression is highly significant (F=500.70, df=1,6 and P<.00001) and explains the variance in the data (R2=.988). Both the intercept and the exponent are significantly different from zero (P<.00001). These results will be used to recommend the number of macrohabitats that should be protected.

Discussion
Because of the imminent threats to ecosystems and their organisms across the globe (Myers, 1988, 1990; Sisk et al., 1994), a strategy of biological assessments across landscapes must target the best and most valuable habitats for immediate conservation and for detailed study. To provide maximal value to the process, inventories should provide not only surveys of the organisms and vital environmental correlates but also hypotheses about community structure or habitat partitioning within the region as well as resilience to environmental threats.
This task is particularly daunting in application to aquatic ecosystems within tropical South America because of their often-unappreciated dynamics and complexities (Goulding, 1980, 1981; Goulding et al., 1988; Machado- Allison, 1994; Barthem and Goulding, 1997). Given their importance and challenges for conservation (Naiman et al., 1995; Pringle, 1997; Pringle et al., 2000), critical information needs to be compiled from tropical aquatic systems. Human development strategies (e.g. Almeida and Campari, 1995) within the basins of the principal rivers of South America often completely ignore the impacts to and needs of aquatic ecosystems.
The methods used in this paper are designed to test the null hypotheses of randomness of distribution of the fishes (or any organisms) in relation to geography or macrohabitats. Nonhomogeneous, patterned distributions require different conservation strategies than does the situation where organisms are randomly or homogeneously distributed (Atmar and Patterson, 1993; Christensen, 1997). Our methods are appropriately conservative and robust, and have the advantage in that we can determine whether the similarity is greater than or less than random expectations. Though the use of point-source data is not necessarily conservative, rapid assessment programs must provide beginning insights into conservation plans to protect ecosystems and biodiversity to the maximum extent possible. We are persuaded by the argumentation of Shrader-Frechette and McCoy (1993) that the ethics of conservation require that we risk a type-I error (rejection of a true null hypothesis) rather than a type-II error (failure to reject a false null hypothesis). In the case presented here this means that it is better to err on the side of recognizing structure within a biological assemblage. The conservation plan will be more stringent than that required to conserve a homogeneously distributed community (Barrett and Barrett, 1997; Christensen, 1997).
We first discuss the results in relation to the two null hypotheses that the distributions of fishes in Río Paraguay are homogeneous with respect to i) subregions; and ii) macrohabitats. We then use that information in a predictive fashion relative to the current environmental threats to the region. We will attempt to provide lists of taxa as profiles for the assemblages that might survive under certain conditions.
Distributional Patterns and Null Hypotheses
For both subregions and macrohabitats, similarities among fish faunas were greater than expected at random (P<0.01) indicating interdependency on variables associated with geography or environment. We rejected (P<<0.001) null hypotheses of random pattern using entropy measures for the subregion or macrohabitat by species matrices. In the case of subregions, patterns of similarity and patterns of shared species, both exclusive and ubiquitous (Figures 5, 6) allowed us to discount smooth faunal turnover (clinal) as the pattern. Rather, the subregion effect comprises two groups, a Río Paraguay group and a Río Apa group, within which there are nested subsets. For macrohabitats, nested patterns are most consistent with i) a fauna being derived from (i.e., moving through) riverine habitats on seasonal cycles of flooding. and ii) a Río Apa beach-clear water group distinguishable from a group of macrohabitats connected to the Río Paraguay.
The results are consistent with published literature on the strong geographic and macrohabitat related effects on freshwater fish distributions (Balon and Stewart, 1983; Balon et al., 1986; Hawkes et al., 1986; Matthews, 1986; Ibarra and Stewart, 1989; Cox Fernandes, 1995; Chernoff et al., 1999). These and the current study refute statements by Lowe-McConnell (1987) and Goulding et al. (1988) that fish communities in the Neotropical lowlands are unstructured or random. Diversity of fish species in Río Paraguay is a non-random interaction of environmental quality and habitat diversity distributed through the river basin. Only 9 out of 173 fish species (5.2%) were found in all subregions; not a single species occurred in each of the eight major macrohabitats. In contrast, Chernoff et al. (1999) found 15.6% of the 313 species living in all six subregions of the Upper Río Orthon watershed, Bolivia.
The Río Paraguay has its headwaters to the north and northwest, and flows from the tropics to the temperate zone. While Gorman and Karr (1978) have postulated that species richness should increase in the lower reaches of rivers, this is not the case for Río Paraguay (Bonetto, 1986; Britski et al., 1999). Because the water temperature decreases downstream, the numbers of fish species drop off as one proceeds to the La Plata section of the river (Bonetto, 1986). The species richness is much higher in the Pantanal than in the section of Río Paraguay that we sampled; the Upper Río Paraguay subregion had slightly more species than other regions. Importantly, many of the species collected uniquely in the Upper Río Paraguay subregion are known from the Pantanal (Britski et al., 1999; Toledo- Piza, 2001).
The pattern of species associations among the macrohabitats was found to be congruent with that for the subregions. The two network analyses (Figures 7, 8) emphasize four important features that are more easily determined from the non-hierarchical cluster analysis. The first is that the Río Paraguay beaches serve as the primary population source or sink for all of the other macrohabitats. The Río Paraguay beaches are the most species rich and share the highest number of species with each of the other macrohabitats (Table II).
The second is that patterns of similarities and shared taxa demonstrate the role of floods in structuring faunal assemblages. Waters flood from the main river channel over beaches into the forests and into local depressions forming lagoons, which then may be stranded in the dry season. Twenty-nine of the 39 species found in lagoons were also found in the flooded forests; none were found exclusively in these two macrohabitats (Table II). It follows that the second largest observed similarity is between the backwater and flooded forest macrohabitats (Table II, Figure 8), based upon species that prefer vegetation and less current (e.g. Apistogramma spp., Cheirodon piaba, Hyphessobrycon eques, Odontostilbe paraguayensis, Otocinclus vestitus, Pyrrhulina australis). These species not only have important roles within the ecosystem (see Lowe-McConnell, 1987) but also have high value in the ornamental fish trade.
The third is that the fishes that inhabit the deeper environments of the main channel often leave deeper waters to inhabit the beach habitats during the evenings or early mornings. The fish species collected in the trawls and in the gill nets are only small portion of what is most certainly much larger fauna (Chernoff et al., 1999). The only significant similarity found between the main channel and any other macrohabitat was to the Río Paraguay beaches (Table III).

The fourth is that the relationship between the species of fishes found along Río Apa beaches and clearwater macrohabitats represents another grouping. These macrohabitats shared 37 species comprising the largest assemblage of ornamental and relatively smaller species (<120mm standard length), such as those in the Astyanax, Ancistrus, Apareiodon, Apistogramma, Bujurquina, Characidium, Cheirodon, Corydoras, Hemigrammus, Holoshesthes, Moenkhausia, Otocinclus, Odontostilbe and Xenurobrycon genera. The environments that provide critical habitat for many of these smaller species beyond the flood zone of Río Paraguay must become a key element of any conservation plan for the region.
Despite the complexity of some of the patterns that were discovered, we can derive a preliminary plan for the conservation of the ichthyofauna based upon the species accumulation analysis (Figure 9). The analysis indicates the number of macrohabitats in good health that are required to protect the majority of species of fishes. More than 90% of the biodiversity may be protected if a biologically significant area containing at least four macrohabitats (Río Paraguay and Río Apa beaches, and two flood zone macrohabitats) is set aside. What is not known as yet is how much area constitutes biologically significant, meaning of large enough area to maintain sustainable populations of the species.
Analysis of Threats
The major threats facing the Río Paraguay drainage are diverse, with short- and long-term consequences. They range from unregulated fisheries to habitat conversion to channelization and dredging (Chernoff et al., 2001). These activities threaten greatly the aquatic ecosystems of the Río Paraguay drainage. Large scale alterations to the Río Paraguay drainage will result not only in a major loss of biodiversity but also important commercial losses in fisheries.
In order to establish a baseline for estimation of the biological consequences of environmental threats, we provide a hypothesis about those fish species that might be the most resilient to environmental destruction. These lists are based upon breadth of fish species distributions across the five subregions and across the eight macrohabitats that we sampled within the Río Paraguay basin. The breadth of distribution may be taken as an estimator of environmental tolerance or survivability. We define common taxa to be those fish species that were found in at least three subregions and in five macrohabitats. There are 31 such species (Table III). The list of common species includes taxa from each of the major groups: tetras and their allies (characiforms), catfishes (siluriforms), electric fishes (gymnotiforms), and cichlids (Cichlidae). This assemblage is the one we consider most likely to exist in highly modified habitats but comprises only 17.9% of the species that we captured (appendix 13 in Toledo-Piza et al., 2001). The following discussion of threats is divided into subsections for clarity.
Hidrovia Paraguay-Paraná.
Brief descriptions of the Hidrovia Project can be found in Heath (1995), Ponce (1995), Hamilton (1999), Swarts (2000) and Pringle et al. (2000). The project intends to deepen and, perhaps, straighten the Río Paraguay, making a connection into the Amazon Basin in Bolivia. The immediate threats to the biotic systems of Río Paraguay come from channelization and river straightening activities. Secondary threats include pollution (bilge water and petroleum products) and interchange of Amazonian and Río Paraguay faunas.
Río Paraguay is a relatively shallow river above Asunción and meanders greatly. The depth of the river will need to be increased substantially in order to accommodate ocean going container ships (Heath, 1995; Ponce, 1995). The installation and maintenance of a commercial shipping channel will require constant dredging (as is done in the Río Orinoco, Venezuela). Rock outcroppings, such as near Cerritos Pão de Açúcar, may need to be eliminated. The overall effect of placing a large channel and straightening of the river will be to change the pattern of hydrography and sedimentation in the river (Hamilton, 1999; Ponce, 1995; Pringle et al., 2000). The deeper channel will actively pull water out of tributaries and from flooded lowlands. More water will exit the system more rapidly and will significantly decrease the degree and the time of inundation of the broad flood plains of the Chaco and the Pantanal (see Hamilton, 1999).
Reduction or prevention of flooding in tropical rivers is perhaps the greatest threat not only to biodiversity but also to productivity (Goulding, 1981; Petts, 1985; Lowe-McConnell, 1987; Goulding et al., 1988; Machado-Allison, 1994; Ponce, 1995; McCulley, 1996). Much of the dependence of the Río Paraguay fish species on annual flooding is evident in our data (Figure 8). Reduction of inland flooded macrohabitats would remove a critical resource for fishes, both for food gathering and reproduction, and for nursery areas. The number of species found in seasonally flooded habitats that result from inundations is almost as great as the number found on beaches (Table II). Of the captured species, 90 (52%) were found in four macrohabitats, including ornamental and commercially valuable food fishes. For example, all of the pacus and pacu-relatives are important components of those habitats, including Metynnis mola, Myleus tiete, Piaractus mitrei, and Mylossoma duriventre. Other food or potential food fishes included: Psectrogaster curviventris, Pygocentrus nattereri, Liposarcus anisitsi, Cochliodon sp., and Triportheus nematurus. There were numerous ornamentals that are common in the aquarium trade, including Gymnotus carapo, Eigenmannia trilineatus, Pyrrhulina australis, Otocinclus spp., Hyphessobrycon eques, Gymnocorymbus ternetzi, Bujurquina vittata, and Apistogramma spp.
Although the common species (Table III) are all found in the list of 90, they too are dependent upon maintenance of the flooding cycle to undergo their life cycles. Many if not most of the species in the list of 90, begin to yolk their eggs and spawn either on the rising floods or directly in the flooded zone (Goulding, 1981; Lowe-McConnell, 1987, Machado-Allison, 1994). Thus, reduction or elimination of the areas of inundation could result in almost 50% loss of the total ichthyofauna. More than half of the 90 species were not discovered in the Río Apa or clear water habitats.
Another consequence of increased drainage of the Río Paraguay due to Hidrovia or similar projects would be a vast reduction of Río Apa and Riacho La Paz during the dry season. The Río Apa-Riacho La Paz region contains minimally 30 fish species that are associated with the terra-firme, more headwater-like habitats and were not found elsewhere. These species are also potentially imperiled.
For both the Río Paraguay macrohabitats and the Río Apa- Riacho La Paz zone, the Pantanal of Mato Grosso do Sul would be a likely source from which recolonization of the fish fauna might be possible. However, the flood plains of the Pantanal, especially in the north, will shrink in size (Hamilton, 1999) and we predict will suffer heavy casualties. Using our data, and the heterogeneous patterns of distribution and the dependency upon the flooding cycle, we calculate that more than 60% of the fish species could be eliminated from the basin if the Hidrovia Project is implemented. The economic costs of this extinction must be explored.
Habitat Conversion in Río Apa and Riacho La Paz.
Much of the Río Apa sub-basin is used for agriculture and ranching. The area, however, is not stable and continued habitat conversion is underway. Because this section is more arid, with drier soils, river water is used for irrigation. Additionally, because the waters are clear and not turbid, there is damage to the ecosystem from sedimentation and erosion (Chernoff et al., 2001). The aquatic ecosystems of the Río Apa basin are very fragile.
Along Riacho La Paz there are approximately 30 fish species that were not found in the more lowland habitats. This represents approximately 17.3% of the fauna that is not easily replaced. The closest sources for recolonization are in the headwaters of the Pantanal or in Río Paraná. These are long distances and we doubt that these fishes are active long-distance colonizers (e.g. Microcharacidium). Continued habitat conversion could eliminate these species that are not found elsewhere in this portion of the Río Paraguay basin.
Conclusions and Recommendations
1. The freshwater fishes are non-randomly distributed with respect to subregions. There is a relatively sharp boundary between Río Apa-Riacho La Paz and Río Paraguay, with more than a 50% turnover in species of fishes. Therefore, it is critical to designate two areas within the Río Paraguay basin as core conservation areas: 1) Upper Río Paraguay-Río Negro and 2) Río Apa- Riacho La Paz. Together they represent more than 80% of the diversity within the region.
2. The Río Apa-Riacho La Paz zone is highly threatened because land conversion (agriculture, ranching, etc.) is actively changing the characteristic of the ecosystem. Extinction or reduction of the fauna in these streams will have serious consequences for the overall biodiversity because potential sources of organisms for recolonization are very far away (Pantanal headwaters and Río Paraná).
3. The freshwater fishes are non-randomly distributed with respect to macrohabitats within the Río Paraguay basin. Beach habitats serve as source populations for other habitats that extend into the forest. These habitats are interconnected during periods of inundation. Therefore, it is critical that the natural flood cycle be maintained and that broad areas for inundation be modified as little as possible. Interference or serious reduction of the flood cycle and inundated habitats could result in as much as a 60% loss of the species in the basin.
4. Sufficient area of four macrohabitats (Río Paraguay and Río Apa beaches, plus two flood zone habitats) are minimally necessary to protect the majority of the fishes. It is critical, however, to determine how much area is needed to support sustainable populations.
4. Sufficient area of four macrohabitats (Río Paraguay and Río Apa beaches, plus two flood zone habitats) are minimally necessary to protect the majority of the fishes. It is critical, however, to determine how much area is needed to support sustainable populations.
ACKNOWLEDGEMENTS
The fieldwork and AquaRAP program was funded through the generosity of the Rufford Foundation to Conservation International. The authors are grateful to Leeanne Alonso, Bruce Patterson, Matthew Leibold and Jensen R. Montambault for comments on the manuscript or advice, and to Mónica Toledo- Piza, Jaime Sarmiento, Darío Mandleburger and Mirta Medina for fieldwork. Equipment used was provided by thoughtful gifts from the Comer Science and Education Foundation, Jay Fahn, and Joan and Selma Goldstein. Lastly, the authors express their gratitude to John McCarter, Russel Mittermeier and Peter Seligman for their continuing support of the AquaRAP program.
REFERENCES
1. Almeida ALO, Campari JS (1995) Sustainable development of the Brazilian Amazon. Oxford University Press: New York, USA. 189 pp. [ Links ]
2. Atmar W, Patterson BD (1993) The measure of order and disorder in the distribution of species found in fragmented habitat. Oecologia 96: 373-382. [ Links ]
3. Balon EK, Crawford SS, Lelek A (1986) Fish communities of the upper Danube River (Germany, Austria) prior to the new Rhein- Main-Donau connection. Environmental Biology of Fishes 15: 243-271. [ Links ]
4. Balon EK, Stewart DJ (1983) Fish assemblages in a river with unusual gradient (Luongo, Africa-Zaire system), reflections on river zonation and description of another new species. Environmental Biology of Fishes 9: 225-252. [ Links ]
5. Barrett NE, Barrett JP (1997) Reserve design and the new conservation theory. In 5. Pickett STA, Ostfield RS, Schachak M, Likens GE (Eds.) The ecological basis of conservation: heterogeneity, ecosystems, and biodiversity. Chapman and Hall: New York, USA. pp 236-251. [ Links ]
6. Barthem R, Goulding M (1997) The catfish connection: ecology, migration, and conservation of Amazon predators. Columbia University Press: New York, USA. 144 pp. [ Links ]
7. Bonetto AA (1986) Fish of the Paraná system. In 7. Davies BR, Walker KF (Eds.) The ecology of river systems. W. Junk: Dordrecht, Netherlands. pp 573-588. [ Links ]
8. Britski HA, de Silimon KZS, Lopes BS (1999) Peixes do Pantanal: manual de identificação. EMBRAPA: Brasília, Brasil. 184 pp. [ Links ]
9. Chernoff B, Willink PW, Sarmiento J, Machado- Allison A, Menezes N, Ortega H ( 1999) Geographic and macrohabitat partitioning of fishes in Tahuamanu-Manuripi region, Upper Río Orthon basin, Bolivia: conservation recommendations. In Chernoff B, Willink PW (Eds.) A biological assessment of aquatic ecosystems of the Upper Río Orthon basin, Pando, Bolivia. Bulletin of Biological Assessment 15. Conservation International. Washington DC, USA. pp. 51-68. [ Links ]
10. Chernoff B, Willink PW, Montambault J (2001) A biological assesement of the aquatic ecosystem of the Río Paraguay basin, Alto Paraguay, Paraguay. Bulletin of Biological Assessment 19. Conservation International: Washington DC, USA. 156 pp. [ Links ]
11. Christensen NLJr (1997) Managing for heterogeneity and complexity on dynamic landscapes. In 11. Pickett STA, Ostfield RS, Schachak M, Likens GE (Eds.) The ecological basis of conservation: heterogeneity, ecosystems, and biodiversity. Chapman and Hall: New York, USA. pp 167-186. [ Links ]
12. Cox Fernandes C ( 1995) Diversity, distribution and community structure of electric fishes (Gymnotiformes) in the channels of the Amazon River system, Brasil. PhD Dissertation. Duke University: Durham, NC, USA. 394 pp. [ Links ]
13. Gabriel KR, Sokal RR (1969) A new statistical approach to geographic variation analysis. Systematic Zoology 18: 259-278. [ Links ]
14. Goulding M (1980) The fishes and the forests: explorations in Amazonian natural history. University of California Press: Los Angeles, CA, USA. 280 pp. [ Links ]
15. Goulding M (1981) Man and fisheries on an Amazon frontier. W.Junk: Boston, USA. 137 pp. [ Links ]
16. Goulding M, Carvalho ML, Ferreira EG (1988) Río Negro: rich life in poor water: Amazonian diversity and foodchain ecology as seen through fish communities. SPB Academic Publishing: The Hague, Netherlands. 200 pp. [ Links ]
17. Gorman OT, Karr JR (1978) Habitat structure and stream fish communities. Ecology 59: 507-515. [ Links ]
18. Hamilton SK (1999) Potential effects of a major navigation project (the Paraguay-Paraná Hidrovía) on inundation in the Pantanal floodplains: Regulated Rivers. Res.and Manag. 15: 289-299. [ Links ]
19. Hawkes CL, Miller DL, Layher WG (1986) Fish ecoregions of Kansas: stream fish assemblage patterns and associated environmental correlates. Environmental Biology of Fishes 17: 267-279. [ Links ]
20. Heath R (1995) Hells highway. New Scientist 146: 22-25. [ Links ]
21. Ibarra M, Stewart DJ (1989) Longitudinal zonation of sandy beach fishes in the Napo River basin, eastern Ecuador. Copeia 1989: 364- 381. [ Links ]
22. Leibold MA, Mikkelson GM (2002) Coherence, species turnover, and boundary clumping: elements of meta-community structure. Oikos 97: 237-250. [ Links ]
23. Lowe-McConnell RH (1987) Ecological studies in tropical fish communities. Cambridge University Press: New York, USA. 382 pp. [ Links ]
24. Machado-Allison A (1987) Los peces de los Llanos de Venezuela: un ensayo sobre su historia natural. Universidad Central de Venezuela. Consejo de Desarollo Cientifico y Humanistico: Caracas, Venezuela. 141 pp. [ Links ]
25. Machado-Allison A (1994) Factors affecting fish communities in the flooded plains of Venezuela. Acta Biol.Venez. 15: 59-75. [ Links ]
26. Matthews WJ (1986) Fish faunal breaks and stream order in the eastern and central United States. Environmental Biology of Fishes 17: 81-92. [ Links ]
27. McCulley P (1996) Silenced rivers. Zed Books: New Jersey, USA. 320 pp. [ Links ]
28. Myers N (1988) Threatened biotas: hotspots in tropical forests. The Environmentalist 8: 1-20. [ Links ]
29. Myers N (1990) The biodiversity challenge: expanded hot-spots analysis. The Environmentalist 10: 243-256. [ Links ]
30. Naiman RJ, Magnusin JJ, McKnight DM, Stanford JA (1995) The Freshwater imperative. Island Press: Washington DC, USA. 165 pp. [ Links ]
31. Patterson BD (1987) The principal of nested subsets and its implications for biological conservation. Conservation Biology 1: 323-334. [ Links ]
32. Petts GE (1985) Impounded rivers. Wiley: New York, USA. 344 pp. [ Links ]
33. Ponce V (1995) Hydrologic and environmental impact of the Parana-Paraguay Waterway on the Pantanal of Mato Grosso, Brazil. Special Report of the Environmental Defense Fund. Berkeley: CA, USA. 124 pp. [ Links ]
34. Pringle CM (1997) Expanding scientific research programs to address conservation challenges in freshwater ecosystems. In 34. Pickett STA, Ostfield RS, Schachak M, Likens GE (Eds.) The ecological basis of conservation: heterogeneity, ecosystems, and biodiversity. Chapman and Hall: New York, USA. pp 305-319. [ Links ]
35. Pringle CM, Freeman MC, Freeman BJ (2000) Regional effects of hydrologic alterations on riverine macrobiota in the New World: tropical - temperate comparisons. BioScience 50: 807-823. [ Links ]
36. Rice WR (1989) Analyzing Tables of statistical tests. Evolution 43: 223-225. [ Links ]
37. Rohlf FJ, Sokal RR ( 1995) Statistical Tables. 3rd ed. Freeman. New York, USA. 199 pp. [ Links ]
38. Shrader-Frechette KS, McCoy ED (1993) Methods in ecology. Cambridge University Press: New York, USA. 328 pp. [ Links ]
39. Sisk TD, Launer AE, Switky KR, Ehrlich and PR (1994) Identifying extinction threats. BioScience 44: 592-604. [ Links ]
40. Sneath PHA, Sokal RR (1973) Numerical Taxonomy. Freeman: San Francisco, USA. 573pp. [ Links ]
41. Sokal RR, Rohlf FJ (1995) Biometry. 3rd ed. Freeman: New York, USA. 887 pp. [ Links ]
42. Swarts FA (Ed.) (2000) The Pantanal: understanding and preserving the worlds largest wetland. Paragon: St. Paul, MN, USA. 301 pp. [ Links ]
43. Toledo-Piza M (2001) Geographic distribution of fish species collected during the AquaRAP expedition to the Departmento Alto Paraguay, Paraguay. In 43. Chernoff B, Willink PW (Eds.) A biological assessment of the aquatic ecosystems of the Río Paraguay basin, Departamento Alto Paraguay, Paraguay. Bulletin of Biological Assessment 19. Conservation International: Washington DC, USA. pp 148-156. [ Links ]
44. Toledo-Piza M, Chernoff B, Mandelburger D, Medina M, Sarmiento J, Willink PW (2001) Diversity and abundance of fishes in the Upper and Lower Río Paraguay basin and the Río Apa sub-basin. In 44. Chernoff B, Willink PW (Eds.) A biological assessment of the aquatic ecosystems of the Río Paraguay basin, Departamento Alto Paraguay, Paraguay. Bulletin of Biological Assessment 19. Conservation International: Washington DC, USA. pp 73-79. [ Links ]
45. Willink PW, Chernoff B, Alonso LE, Montambault JR, Lourival R (Eds.) (2000) A Biological Assessment of the Aquatic Ecosystems of the Pantanal, Mato Grosso do Sul, Brasil. Bulletin of Biological Assessment 18. Conservation International: Washington DC, USA. 306pp. [ Links ]
