











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.31 n.10 Caracas oct. 2006
IDENTIFYING PRIORITY AREAS FOR CONSERVATION IN MEXICAN TROPICAL DECIDUOUS FOREST BASED ON TREE SPECIES
Eva M. Cué-Bär, José Luis Villaseñor, Juan J. Morrone
and Guillermo Ibarra-Manríquez
Eva M. Cué-Bär. M.S. in Biological Science, University of Leipzig, Germany. Ph.D. student, Universidad Nacional Autónoma de México (UNAM). Address: Antigua Carretera a Pátzcuaro Nº 8701. Col. San José de la Huerta. 58190 Morelia, Michoacán, México. e-mail: evacue@oikos.unam.mx
José Luis Villaseñor. Ph.D. in Plant Systematics, The Claremont Graduate School, USA. Researcher, UNAM, Mexico. e-mail: vrios@ibiologia.unam.mx
Juan J. Morrone. Ph.D. in Natural Sciences, Universidad Nacional de La Plata, Argentina. Profesor, UNAM, Mexico. e-mail: jjm@hp.fciencias.unam.mx
Guillermo Ibarra-Manríquez. Ph.D. in Sciences, UNAM, Mexico. Researcher, UNAM, Mexico. e-mail: gibarra@oikos.unam.mx
SUMMARY
The main objective was to identify and rank areas for the conservation of exclusive, or nearly exclusive, tree species of the tropical deciduous forest in Mexico, a land cover type greatly endangered worldwide. A list of 425 tree species (67.5% endemic to Mexico), including 56 families and 185 genera registered at the state level, was compiled from an exhaustive revision of specialized floristic literature. The conservation status of these species was assessed by registering their presence in the tropical deciduous forest areas, located within the limits of Mexican natural protected areas. Parsimony analysis of endemicity led to identify 16 areas of endemism, supported by 54 synapomorphies and 73 autapomorphies (72.4% endemic to Mexico). Protected areas include a low to medium proportion both of widespread tree species (129 species, 30.3%) and species characterizing areas of endemism (56 species out of 127, 44.1%). In the latter group, only 7 species are included within a risk category. Analyses of complementarity (total richness of species and of species within areas of endemism) and of phylogenetic diversity (families and genera) are coincident in assigning high conservation priority for areas in the states of Chiapas, Guerrero, Jalisco, Michoacán, Oaxaca, and Yucatán. Based on the results, specific recommendations are provided for the development of strategies of tree conservation in the Mexican tropical deciduous forest.
IDENTIFICACIÓN DE ÁREAS PRIORITARIAS PARA LA CONSERVACIÓN DEL BOSQUE TROPICAL CADUCIFOLIO EN MÉXICO BASADA EN ESPECIES ARBÓREAS
RESUMEN
El objetivo principal del trabajo fue identificar y jerarquizar áreas prioritarias para la conservación de los árboles que se distribuyen de manera exclusiva o casi exclusivamente en el bosque tropical caducifolio en México, un tipo de vegetación seriamente amenazado a nivel mundial. La información se obtuvo a través de la consulta de literatura florística especializada, con la que se generó una lista de 425 especies arbóreas (67,5% endémicas de México), con 56 familias y 185 géneros registrados a nivel de estado. El grado de conservación de las especies se evaluó registrando su presencia en las Áreas Naturales Protegidas de México que incluyen dentro de sus límites porciones con bosque tropical caducifolio. Por análisis de parsimonia se determinaron 16 áreas de endemismo, sustentadas por 54 sinapomorfias y 73 autapomorfias (72,4% endémicas de México). Las áreas protegidas incluyen una baja a mediana proporción de las especies arbóreas de amplia distribución (129 especies, 30,3%) y de aquellas que caracterizan las áreas de endemismo (56 de 127 especies, 44,1%). En este último grupo, solo 7 especies están incluidas en una categoria de riesgo. El análisis de complementaridad (basado en la riqueza de especies y de aquellas localizadas en áreas de endemismo) y de diversidad filogenética (número de familias y géneros encontrados en cada estado) coinciden en asignar como áreas prioritarias para la conservación a los estados de Chiapas, Guerrero, Jalisco, Michoacán, Oaxaca y Yucatán. Con base en los resultados se proponen recomendaciones para el desarrollo de estrategias para asegurar la conservación de los árboles del bosque tropical caducifolio mexicano.
IDENTIFICAÇÃO DAS ÁREAS PRIORITÁRIAS PARA A CONSERVAÇÃO DO BOSQUE TROPICAL CADUCIFÓLIO NO MEXICO BASEADA EM ESPÉCIES ARBÓREAS
RESUMO
O objetivo principal do trabalho foi identificar e hierarquizar áreas prioritárias para a conservação das árvores que se distribuem de maneira exclusiva ou quase exclusivamente no bosque tropical caducifólio no México, um tipo de vegetação seriamente ameaçada a nível mundial. A informação se obteve através da consulta de literatura florística especializada, com a qual foi gerada uma lista de 425 espécies arbóreas (67,5% endêmicas do México), com 56 famílias e 185 gêneros registrados em nível de estado. O grau de conservação das espécies se avaliou registrando sua presença nas Áreas Naturais Protegidas do México que incluem dentro de seus limites porções com bosque tropical caducifólio. Por analise de parcimônia se determinaram 16 áreas de endemismo, sustentadas por 54 sinapomorfias e 73 autapomorfias (72,4% endêmicas do México). As áreas protegidas incluem uma baixa a média proporção das espécies arbóreas de ampla distribuição (129 espécies, 30,3%) e de aquelas que caracterizam as áreas de endemismo (56 de 127 espécies, 44,1%). Neste último grupo, somente 7 espécies estão incluídas em uma categoria de risco. A análise de complementaridade (baseada na riqueza de espécies e de aquelas localizadas em áreas de endemismo) e de diversidade filogenética (número de famílias e gêneros encontrados em cada estado) coincidem em designar como áreas prioritárias para a conservação aos estados de Chiapas, Guerrero, Jalisco, Michoacán, Oaxaca e Yucatán. Baseado nos resultados se propõe recomendações para o desenvolvimento de estratégias para assegurar a conservação das árvores do bosque tropical caducifólio mexicano.
KEY WORDS / Biogeography / Complementarity Analysis / Neotropics/ Parsimony Analysis of Endemicity / Phylogenetic Diversity /
Received: 02/15/2006. Modified: 09/01/2006. Accepted: 09/04/2006.
Endemism is related to geographical or ecological events resulting in the isolation of populations from one or several species (vicariance). Because of this, islands, mountains, and deserts are particularly rich in endemic taxa (Major, 1988; Brown and Lomolino, 1998). Explanations for the high number of endemic plant taxa in Mexico include that some areas behave like ecological islands, as well as events and environmental conditions in the geological past that lead to suggest that several regions acted as refuges in pre-Pleistocene or Pleistocene times (Wendt, 1993). Rzedowski (1991a) points to the tropical deciduous forest (TDF) as a clear example of an ecological island, because of its flora exclusivity originating through geographical isolation. In Mexico, this type of vegetation, encompassing tropical subdeciduous and thorn forests, includes nearly 6000 species of phanerogamous plants, equivalent to 20% of the total flora of the country, and nearly 40% of endemic species (Rzedowski, 1991a, b). Moreover, a significant richness and level of endemism has been attributed to vertebrate species in the Mexican TDF (Ceballos and García, 1995).
It has been hypothesized that the TDF has been the place of diversification of numerous plant lineages. At the family level, the Asteraceae, Bignoniaceae, Burseraceae, Cactaceae, Convolvulaceae, Euphorbiaceae, Fabaceae, Malvaceae, and Rubiaceae are relevant cases (Rzedowski, 1978, 1991a, b; Gentry, 1995). More precise data are available for the Euphorbiaceae, with 384 species registered in Mexico, of which 23% is present in the TDF and 36% of these species being exclusive to this land cover type (Martínez-Gordillo et al., 2002). At the genus level Bursera (Burseraceae) is a remarkable case, given that 92% of its 82 Mexican species are basically restricted to these forests (Rosalinda Medina personal communication), with 6 areas of high concentration of species having limited distribution (Rzedowski et al., 2005). Additional evidence for the floristic relevance of the TDF is also provided by the results of a census of the woody plant flora in 0.1ha areas in Chamela (Jalisco) and Caleta (Michoacán), located along the Pacific Ocean coast (Lott et al., 1987; Trejo and Dirzo, 2002); such regions are among the 10 most species-rich worldwide (Gentry, 1995; Phillips and Miller, 2002; Trejo and Dirzo, 2002).
Areas of endemism have been given several meanings in the biogeographic literature (e.g. Müller, 1973; Platnick, 1991; Morrone, 1994; Posadas and Miranda-Esquivel, 1999). In general, they may be defined as areas presenting congruent and non-random distribution patterns for two or more taxa, which may be or may not be ecological or phylogenetically related. The complexity of identifying areas of endemism increases in proportion to the number of taxa being analyzed. Morrone (1994) suggested the use of the parsimony analysis of endemicity method (PAE) to locate endemism areas based on their shared taxa (usually species), and finding the most parsimonious solution under the assumption that the resulting groups of species have a common history (Rosen, 1988; Morrone, 1994; Morrone and Crisci, 1995).
The PAE approach has been applied to several taxa and regions of the world (e.g. Linder, 2001; García-Barros et al., 2002; Rovito et al., 2004; Silva et al., 2004). In Mexico, several plant and animal groups have been analyzed at different scales (Morrone et al., 1999, 2002; Luna-Vega et al., 1999; Morrone and Escalante, 2002; Escalante et al., 2003; Espadas et al., 2003; Rojas et al., 2003). It has been suggested that parsimony analyses of endemicity may help choose areas for conservation (Posadas and Miranda-Esquivel, 1999; Cavieres et al., 2002; Méndez-Larios et al., 2005), by establishing which areas are unique regarding their composition, and giving priority to those areas displaying a high degree of endemicity.
In conservation biology, criteria and methods to select and to assign priority for conservation purposes to a particular region have been increasing in number and complexity. Among the most utilized options for selecting and ranking areas for conservation are iterative methods based on the principle of complementarity that allows the conservation of the largest number of species within a given region and in the minimum possible number of areas (Margules et al., 1988; Margules and Pressey 2000). This approach has been used for diverse groups of Mexican plant taxa (Villaseñor et al., 1998, 2003; Lira et al., 2002; Dávila et al., 2004).
The high floristic diversity and level of endemicity of the TDF, in addition to its ample distribution in Mexico, point out the need to localize and prioritize its areas of endemism. The importance of such aim is enhanced considering that the TDF is one of the most threatened tropical ecosystems, due mainly to extensive grazing and agriculture (Janzen, 1988; Murphy and Lugo, 1995; Trejo and Dirzo, 2000; Miles et al., 2006). Trees were selected in this study because they meet the requirements for being considered as an indicator group of biodiversity (Stork, 1994), and their relevant role in structure and several ecological functions in the Mexican TDF (Miranda and Hernández-X., 1963; Rzedowski, 1978; Lott et al., 1987; Trejo and Dirzo, 2000). Therefore, the objectives of this study are: 1) to localize the areas of endemism of tree species in the Mexican TDF and the species characterizing them, 2) to establish the priority for conservation of the states that include these endemic areas, based on their species richness and phylogenetic diversity, and 3) to asses the protection level enjoyed by the TDF trees and their risk category.
Materials and Methods
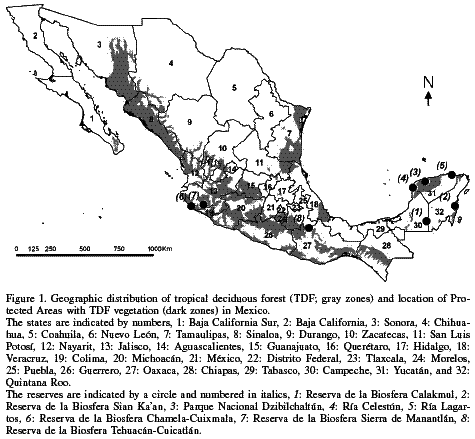
This forest type is distributed in Mexico along a nearly continuous strip along the Pacific slopes from Sonora (~28ºN) to Chiapas (ca. 14º30N), whereas in the Gulf of Mexico it has a discontinuous distribution from Tamaulipas to the Yucatán Peninsula. In addition, it is present in several central states and the southern tip of the Baja California Peninsula (Figure 1), from sea level to 2220masl (Rzedowski, 1978; Trejo and Dirzo, 2000). The total area covered by the TDF has been estimated to be ~8-17% of the total country area (Rzedowski, 1978, 1991a). The TDF is present in 10 of the 11 morphotectonic provinces established for Mexico by Ferrusquía-Villafranca (1993), being absent only in the Chihuahua-Coahuila Plateaus and Ranges province. It grows mainly in rocky, shallow soils or in soils with textures ranging from clay to sand, acid to slightly alkaline, rich in organic matter, and of clear to dark color (Rzedowski, 1978). Climates favoring TDF are dry to semi-dry, warm (Aw or Bs) and with mean annual temperature above 19oC, mean annual precipitation of 400-1350mm, and 6-9 months with precipitation <60mm (Miranda and Hernández-X., 1963; Rzedowski, 1978; Trejo and Dirzo, 2000). According to these authors, this forest is characterized by an upper arboreal stratum between 8-15m tall, and by a loss of foliage in over 75% of its tree components during 5-9 months per year.
Identification of high priority for conservation areas
A database was compiled with tree species that are nearly or totally exclusive to TDF habitats in Mexico, due to some species being also reported from the tropical subdeciduous forest, which is difficult to distinguish clearly from TDF (Rzedowski, 1978). The geographical distribution of these species was determined from specialized literature, such as floristic checklists and regional floras (e.g. McVaugh, 1974; Rzedowski et al. 2005), registering their presence or absence at the state level. Although geopolitical entities do not represent natural areas, they were used as units because this is the level used for delimitation and decrees of Natural Protected Areas in Mexico (Conabio, 1998; Villaseñor et al., 1998; Lira et al., 2002; Dávila et al., 2004). Although it would be desirable to assign conservation priorities at a finer scale (e.g. 0.5o or 1o squares) or by biogeographical natural unities, the lack of confident systematic information at such scales for the species included in the study impedes such an approach at present
A data matrix was created with the presence or absence of species (columns) in 25 states of Mexico registering TDF (rows), adding a hypothetical site coded as 0 for all species to root the cladogram (Morrone, 1994). The data matrix was analyzed using the NONA program (Goloboff, 1999), executed in WINCLADA (Nixon, 2002), with 1000 as the maximum trees kept at each round of analysis, 100 replications, and 10 starting trees per replication. Areas of endemism were delimited as those having at least two exclusive species. In this study, a synapomorphy is a species that is present in two or more endemic areas, and an autapomorphy is a species exclusive of one area. In order to know which species are under protection, we determined their presence in those Protected Areas that recorded species checklists (Bradburn and Darwin, 1982; Durán and Olmsted, 1987; Dávila et al., 1993; Cuevas et al., 1998; Durán et al., 1999a, b; Martínez et al., 2001; Lott, 2002). Finally, the Mexican Official Norm NOM-059-ECOL-2001 (SEMARNAT, 2002) was confronted with the species recorded in the areas of endemism in order to register their risk category.
A complementarity analysis (Vane-Wright et al. 1991) was undertaken with the purpose of establishing the relative priority of each state for the tree species conservation of the TDF. This analysis, based on total species richness, comprises the following steps: 1) the state with the greatest species richness is chosen first, and the taxa present in it are excluded in the following phases of the analysis; 2) step one is repeated with the remaining species (complement); 3) if two or more states have equal values of species richness, the state having the largest total number of species is chosen; 4) if a tie remains, the state located closer to any of the previously selected states is chosen; and 5) the procedure continues until all species have been selected. This algorithm was applied including all 25 states of Mexico with TDF, and the 13 states enclosing endemism areas resulting from PAE. The phylogenetic diversity was also obtained for this latter group of states, by counting the number of families and genera encompassed in the set of species characterizing the endemism areas. This information was utilized to establish conservation priorities (Faith, 1994).
Results
The TDF in Mexico includes 1365 species of trees distributed along the 25 states having this land cover type. Of this total, 425 species were recorded as exclusive or nearly exclusive to the TDF (118 are also found in tropical subdeciduous forests), and a total of 287 species (67.5%) are endemic to Mexico. The comparison of species richness among states shows noticeable differences in Guerrero, Jalisco, Oaxaca, Michoacán, and Chiapas (Table I). The number of species may be accounted for by the extension of TDF present in each state (r= 0.54, p<0.05) but, especially by the species density (r= 0.99, p<0.05). Despite the above, it is interesting to notice that states having relevant extensions of TDF, such as Sinaloa and Sonora, are surpassed in species richness by states having smaller areas, particularly Chiapas and Colima (Table I).

The PAE resulted in five equally parsimonious cladograms. The strict consensus cladogram has 1026 steps in length, a consistency index of 0.41 and a retention index of 0.53 (Figure 2). The cladogram encompasses a total of 127 species (54 synapomorphies and 73 autapomorphies) that characterize 16 areas of endemism; 92 (72.4%) of these species are endemic to Mexico (Table II). Mexico is divided in two main regions (Figure 2): an east-southeast region from San Luis Potosí to Quintana Roo, including endemic areas 1-8, basically integrated by the states along the coast of the Gulf of Mexico and the Yucatán Peninsula; and a western region including endemic areas 9-16, mainly in the states of the Pacific Ocean slope (Chihuahua and Sonora to Michoacán).


The eight Natural Protected Areas with TDF have records of only 129 species (30.3%) of the 425 species included in this work (Table III). Within the reserves there are from 72 (Reserva de la Biosfera Chamela-Cuixmalá, Jalisco) to 14 species (Reserva de la Biosfera Tehuacán-Cuicatlán, Oaxaca and Puebla). Once again, considering the 127 species identified as synapomorphies or autapomorphies, only 56 species (44.1%) were recorded from Natural Protected Areas (Table II). A total of 16 species (12.6%) are present in reserves located in the Yucatán Peninsula (Campeche, Quintana Roo, and Yucatán), their reserves displaying the highest values among Protected Areas shown in Table III. Of the 127 species characterizing endemic areas, only seven are listed in the risk categories as threatened species (Beaucarnea goldmanii, B. hiriartiae and Peltogyne mexicana) and subject to special protection (Stenocereus martinezii, Backebergia militaris, Bursera coyucensis, and B. bonetti).

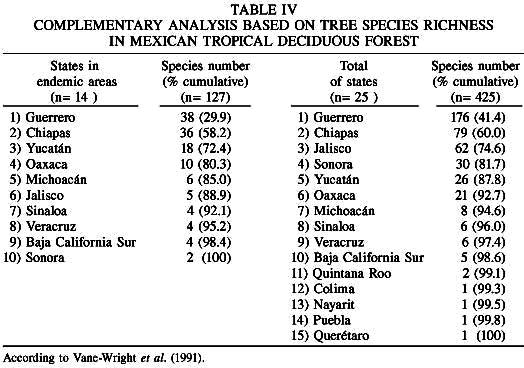
The complementarity analysis based on species richness and species located in areas of endemism, was highly coincident in pointing to the relevance of Chiapas, Guerrero, Jalisco, Oaxaca, and Yucatán within the six higher priorities (Table IV). It is noteworthy that the two higher priorities in both algorithms, Guerrero and Chiapas, would allow over 58% of the species included in the analysis. With regard to the phylogenetic analysis, a high similarity is observed in the states conservation priority based on the number of families or genera that contain, in particular the case of the first five choices which are identical (Table V).

Discussion
Some of the endemic areas identified in the present study had previously been identified by studies applying PAE to different taxa, in particular the endemism area of Baja California Sur (Morrone et al., 1999; Morrone and Escalante, 2002; Escalante et al., 2003; Rojas et al., 2003), area 5 (Veracruz, Chiapas, and Oaxaca; Luna-Vega et al., 1999; Escalante et al., 2003), and area 10 (Sinaloa; Morrone et al., 2002; Escalante et al., 2003). Endemic areas 2 and 3, identified within a distinctive biotic area of Mexico known as the Yucatán Peninsula province (Morrone et al., 2002; Morrone and Escalante, 2002; Escalante et al., 2003; Espadas et al., 2003), deserve a more detailed discussion. The biotic particularity of this region is out of any doubt; however, its southern limit has been the topic of a constant debate among biogeographers who have worked in such area (e.g. Estrada-Loera, 1991; Durán et al., 1998). In our study, this area merges with Chiapas, Oaxaca, and Veracruz (Figure 2), the states with TDF geographically closer to this endemic area (Figure 1). From a biogeographical perspective, the northern portion of Chiapas has been considered by some authors to be a part of this biogeographical province (e.g. Durán et al., 1998, but see also Carnevalli et al., 2001).
The results of the present study show that the Yucatán Peninsula does not relate directly with Chiapas (area of endemism 2), and probably the region should be divided into biogeographic districts (area of endemism 3) as proposed in previous biogeographic works (Estrada-Loera, 1991; Ibarra-Manríquez et al., 2002). Furthermore, given the relevance and complexity of the phytogeographical relationships postulated for the Yucatán Peninsula, in particular with Central America and the Antilles (Estrada-Loera, 1991; Chiappy-Jhones et al., 2001; Ibarra-Manríquez et al., 2002), it would be interesting to perform a PAE that encloses the regions with TDF in Central America and the Antilles. This would enable a clearer delimitation of the endemic areas for this vegetation type.
Likewise, endemic area 12, a part of the Balsas River Basin, was previously identified using PAE with columnar cacti (Dávila et al., 2002), land mammals (Morrone et al., 2002) and helminthic parasites of freshwater fishes (Aguilar-Aguilar et al., 2003), in addition to other biogeographical analyses (Rzedowski, 1978, 1991a; Arita and Santos, 1999; Rzedowski et al., 2005). Physiographically, the Balsas River Basin includes parts of the states of México, Morelos and Puebla, shown in our cladogram as a polytomy. Their segregation from endemic area 12 may be due to unequal floristic knowledge, and the need for a more in-depth exploration, as has been stated by several authors (e.g. Rzedowski, 1978, 1991a; Sosa and Dávila, 1994). Future botanical exploration in the region will aid to establish whether these states include within their territories the synapomorphies, which, in our study, characterize the Balsas River Basin.
Some authors have criticized PAE considering it to be an inadequate cladistic biogeographic method (Humphries and Parenti, 1999; Brooks and van Veller, 2003; Santos, 2005), because it does not consider the phylogenetic relations of the studied groups when discovering relationships between areas. Others have shown its usefulness for a preliminary biogeographic assessment (Escalante, 2002; Escalante et al., 2003). As applied herein, PAE is a useful tool for the identification of areas of endemism (Morrone, 1994; García-Barros, 2003). Once they have been identified, it is possible to postulate hypotheses on their historical relationships applying cladistic biogeographic methods (Morrone and Escalante, 2002; Silva et al., 2004; Méndez et al., 2005). For example, a hypothesis arising from the present work is that the Balsas River Basin could be divided in two regions: eastern (México, Morelos, and Puebla) and western (Guerrero, Jalisco, and Michoacán). A cladistic biogeographic analysis may allow to corroborate this hypothesis. PAE represents an alternative to the lack of cladistic studies (Posadas and Miranda-Esquivel, 1999), more so in regions high in species richness such as Mexico, one of the countries with the highest floristic richness in the whole world (Mittermeir, 1988; Groombridge, 1992; Villaseñor, 2003).
On the other hand, Natural Protected Areas of México protect 5400km2 of TDF, which represents 4% of the total land cover (Mas and Pérez-Vega, 2005), despite that CONABIO (1998) reports the surface area under protection to be only 1.1%. This percentage is low and insufficient, when compared to the 8-10% suggested for the conservation of a given land cover type at a regional level (Rodrigues et al., 2004). Besides, it is important to recognize that the Natural Protected Areas in Mexico differ widely in their legal status and their actual functioning in practice. Among some of the most acute problems of Mexico, is the extensive and intensive use of its natural resources in the absence of management plans. In addition very few reserves have biotic inventories. This deficiency is present in different categories of reserves having TDF, including Biosphere Reserves (e.g. La Sepultura, Chiapas, or Sierra de Abra Tanchipa, San Luis Potosí), making it extremely difficult to manage adequately their natural resources and its biotic components.
The data outlined in the present work provide evidence for the insufficiency of biological knowledge, and, also, a possible misrepresentation of the Natural Protected Areas in Mexico. Nearly 61.3% of the total species associated to this land cover type (261 species), and 63% of the species that characterize the endemic areas are unprotected. Despite the aforementioned fact, it is most encouraging to compare the coincident results of the analyses of richness for the species total, the species inside endemic areas, and the phylogenetic diversity that these areas and states contain, since Chiapas, Guerrero, Jalisco, Michoacán, Oaxaca, and Yucatán have a high priority for conservation. These results agree in part with those of Lira et al. (2002) and Villaseñor et al. (2003), who worked with Cucurbitaceae and the vascular plants of the tropical rain forest, respectively.
Although the conservation recommendations considered in the present study had to be conducted at the state level (see Methods section), it is important to emphasize that their relevance is rather high considering that 1) 425 species were analyzed, which restrict their distribution to the TDF (118 share with the tropical subdeciduous forests, a similar vegetation type); 2) the efforts and resources directed towards the conservation in the future should bee focused to specific areas in each state, taking into account the location and the TDF area they contain (Figure 2, Table I); 3) the conservation importance of the TDF is prioritized in each state at the level of the whole country, on the basis of the totality of restricted species, the species that define the endemism areas, and their supraspecific taxa richness (families and genera); and 4) several of the endemic areas determined for the TDF are coincident with the ones found with other flora and fauna groups utilizing PAE, or other types of biogeographic approaches.
Specific recommendations resulting from this study are 1) to increase the botanical research in each state, specially those that still possess important extensions of TDF and lack an adequate system of Protected Areas (Table I); 2) in the future, to define the distribution patterns of the 127 species that characterize the endemic areas, and to produce specific recommendations for their protection, in particular those that are endemic to México (Table II); 3) to increase the biological knowledge of this set of species in order to asses the degree of threat they face; and 4) to obtain regional inventories, and to carry out demographic and genetic studies of vascular plant species in the Mexican reserves with TDF, in order to asses their conservation representativeness at the country level, and the viability of their long-term conservation.
The TDF is considered worldwide to be a threatened vegetation type (Janzen, 1988; Olson and Dinerstein, 2002; Miles et al., 2006). The results presented herein clearly demonstrate that, until now, the efforts and resources directed towards the biological knowledge and the conservation of the TDF in Mexico have been insufficient. This problem needs to be dealt with as soon as possible, provided the severe threat faced by the TDF (e.g. Rzedowski, 1978; Trejo and Dirzo, 2000). The results of this study are expected to aid in a more representative selection of sites with high priority, and representation for conservation of the arboreal flora (and of their companions belonging to other life forms) of these forests in México.
ACKNOWLEDGEMENTS
The authors thank Rosalinda Medina Lemos for sharing information of the genus Bursera (Burseraceae) and Enrique Ortiz for the elaboration of the study area map. The senior author thanks the Secretaría de Relaciones Exteriores (2002-05) and the Dirección General de Estudios de Posgrado (2005-06) of the Universidad Nacional Autónoma de México (UNAM), for the scholarships for doctoral studies.
References
1. Aguilar-Aguilar R, Contreras-Medina R, Salgado-Maldonado G (2003) Parsimony Analysis of Endemicity (PAE) of Mexican hydrological basins base on helminth parasites of freshwater fishes. J. Biogeog. 30: 1861-1872. [ Links ]
2. Arita HT, Santos K (1999) Conservation biology of nectar feeding bats in México. J. Mammal. 80: 31-41. [ Links ]
3. Bradburn AS, Darwin SP (1982) An annotated checklist of plants. In Thien LB, Bradburn AS, Welden AL (Eds.) The woody vegetation of Dzibilchaltún. A Maya Archeological site in northwest Yucatán, México. Middle American Research Institute. Tulane University. New Orleans, USA. pp. 19-24. [ Links ]
4. Brooks DR, van Veller MGP (2003) Critique of parsimony analysis of endemicity as a method of historical biogeography. J. Biogeog. 30: 819-825. [ Links ]
5. Brown JH, Lomolino MV (1998) Biogeography. 2nd ed. Sinauer. Sunderland, MA, USA. 691 pp. [ Links ]
6. Carnevali FCG, Tapia-Muñoz JL, Jiménez-Machorro R, Sánchez-Saldaña L, Ibarra GL, Ramírez IM, Gómez MP (2001) Notes on the flora of the Yucatán Peninsula II: A synopsis of the orchid flora of the Mexican Yucatán Peninsula and a tentative checklist of the orchidaceae of The Yucatán Peninsula Biotic Province. Harv. Pap. Bot. 5: 383-466. [ Links ]
7. Cavieres LA, Arroyo MTK, Posadas P, Marticorena C, Matthei O, Rodríguez R, Squeo F, Arancio G (2002) Identification of priority areas for conservation in an arid: application of parsimony analysis of endemicity in the vascular flora of the Antofagasta region, northern Chile. Biodiv. Cons. 11: 1301-1311. [ Links ]
8. Ceballos G, García A (1995) Conserving Neotropical biodiversity: The role of dry forest in western México. Cons. Biol. 9: 1349-1353. [ Links ]
9. Chiappy-Jhones C, Rico-Gray C, Gama L, Giddings L (2001) Floristic affinities between the Yucatán peninsula and some karstic areas of Cuba. J. Biogeog. 28: 535-542. [ Links ]
10. Conabio (1998) La diversidad biológica de México. Estudio de país, 1998. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Mexico 349 pp. [ Links ]
11. Cuevas GR, Nuñez LNM, Guzmán, HL, Santana, MFJ (1998) El bosque tropical caducifolio en la Reserva de la Biosfera Sierra de Manantlán, Jalisco-Colima, México. Bol. Inst. Bot. Univ. Guadalajara 5: 445-491. [ Links ]
12. Dávila P, Arias S, Lira R, Villaseñor JL, Valiente A (2002) Phytogeography of the columnar cacti (tribe Pachycereeae) in México: A cladistic approach. In Fleming TH, Valiente A (Eds.) Columnar cacti and their mutualists. Evolution, Ecology and Conservation. University of Arizona Press. Tucson, AZ, USA. pp. 25-41. [ Links ]
13. Dávila P, Villaseñor JL, Medina, L Ramírez, A, Salinas, A, Sánchez-Ken, J, Tenorio P (1993) Listados Florísticos de México. Flora del Valle de Tehuacán-Cuicatlán. Instituto de Biología, UNAM. México. 195 pp. [ Links ]
14. Dávila P, Lira R, Valdés J (2004) Endemic species of grasses in México: A phytogeographic approach. Biodiv. Cons. 13: 1101-1121. [ Links ]
15. Durán R, Olmsted I (1987) Listado florístico de la Reserva de Sian Kaan. Amigos de Sian Kaan. Puerto Morelos, Quintana Roo, Mexico. 71 pp. [ Links ]
16. Durán GR, Trejo-Torres JC, Ibarra-Manríquez G (1998) Endemic phytotaxa of the Peninsula of Yucatán. Harv. Pap. Bot. 3: 263-314. [ Links ]
17. Durán R, Sima P, Juan-Quí M (1999a) Listado florístico de Ría Celestún. Centro de Investigación Científica de Yucatán. Mérida, Yucatán, Mexico. 52 pp. [ Links ]
18. Durán R, Simá P, Juan-Quí M, Méndez M, Tun F (1999b) Listado florístico de la Reserva de Ría Lagartos. Centro de Investigación Científica de Yucatán. Mérida, Yucatán, Mexico. 49 pp. [ Links ]
19. Escalante T, Espinosa D, Morrone JJ (2003) Using parsimony analysis of endemicity to analyze the distribution of Mexican land mammals. Southwest. Naturalist 48: 563-578. [ Links ]
20. Espadas C, Durán R, Argáez J (2003) Phytogeographic analysis of taxa endemic to the Yucatán Peninsula using geographic information systems, the domain heuristic method and parsimony analysis of endemicity. Div. Dist. 9: 313-330. [ Links ]
21. Estrada-Loera E (1991) Phytogeographic relationships of the Yucatán Peninsula. J. Biogeog. 18: 687-697. [ Links ]
22. Faith DE (1994) Phylogenetic diversity: a general framework for the prediction of feature diversity. In Forey PI, Humphries CJ, Vane-Wright RI (Eds.) Systematics and conservation evaluation. Clarendon Press. Oxford, RU. pp. 251-268. [ Links ]
23. Ferrusquía-Villafranca I (1993) Geology of México: a synopsis. In Ramamoorthy TP, Bye R, Lot A, Fa J (Eds.) Biological diversity of México: origins and distribution. Oxford University Press. New York, NY, USA. pp. 3-108. [ Links ]
24. García-Barros E (2003) Mariposas diurnas endémicas de la región Paleártica occidental: Patrones de distribución y su análisis mediante parsimonia (Lepidoptera, Papilionoidea). Graellsia 59: 233-258. [ Links ]
25. García-Barros E, Gurrea P, Luciáñez MJ, Cano JM, Munguira ML, Moreno JC, Sainz H, Sanz MJ, Simón JC (2002) Parsimony analysis of endemicity and its application to animal and plant geographical distributions in the Ibero-Balearic region (western Mediterranean). J. Biogeog. 29: 109-124. [ Links ]
26. Gentry AH (1995) Diversity and floristic composition of Neotropical dry forests. In Bullock HS, Mooney AH, Medina E (Eds.) Seasonally dry tropical forests. Cambridge University Press. Cambridge, UK. pp. 146-190. [ Links ]
27. Goloboff P (1999) NONA, version 2.0. San Miguel de Tucumán, Mexico. [ Links ]
28. Groombridge B (1992) Global biodiversity. Status of the Earths living resources. World Conservation Monitoring Centre. Chapman and Hall. London, UK. 585 pp. [ Links ]
29. Humphries C, Parenti L (1999) Cladistic biogeography, 2nd ed. Academic Press. London, UK. 199 pp. [ Links ]
30. Ibarra-Manríquez G, Villaseñor JL, Durán R, Meave J (2002) Biogeographical analysis of the tree flora of the Yucatán Peninsula. J. Biogeog. 29: 17-29. [ Links ]
31. Janzen D (1988) Tropical dry forest, the most endangered major tropical ecosystem. In Wilson EO (Ed) Biodiversity. Nacional Academy Press. Washington DC, USA. pp. 130-137 [ Links ]
32. Linder HP (2001) On areas of endemism, with an example from the African Restionaceae. Syst. Biol. 50: 892-912. [ Links ]
33. Lira R, Villaseñor JL, Ortiz E (2002) A proposal for conservation of the familiy Curcubitaceae in México. Biodiv. Cons. 11: 1699-1720. [ Links ]
34. Lott JE, Bullock SH, Solis-Magallanes JA (1987) Floristic diversity and structure of upland and Arroyo Forests of coastal Jalisco. Biotropica 19: 228-235. [ Links ]
35. Lott JE (2002) Lista anotada de las plantas vasculares de Chamela-Cuixmala. In Noguera FA, Vega JH, Garcia AN, Quesada M (Eds.) Historia Natural de Chamela. Instituto de Biología, UNAM. Mexico. pp. 99-136. [ Links ]
36. Luna-Vega I, Alcántara O, Espinosa D, Morrone JJ (1999) Historical relationships of the Mexican cloud forest: A preliminary vicariance model applying parsimony analysis of endemicity to vascular plant taxa. J. Biogeog. 26: 1299-1305. [ Links ]
37. Major J (1988) Endemism: a botanical perspective. In Myers AA, Giller PS (Eds.) Analytical biogeography: an integrated approach to the study of animal and plant distributions. Chapman and Hall. London, UK. pp. 117-146. [ Links ]
38. Margules CR, Pressey RL (2000) Systematic conservation planning. Nature 403: 243-253. [ Links ]
39. Margules CR, Nicholls AO, Pressey RL (1988) Selecting networks of reserves to maximise biological diversity. Biol. Cons. 43: 63-76. [ Links ]
40. Martínez E, Sousa M, Ramos CH (2001) Listados Florísticos de México. Región de Calakmul, Campeche. UNAM. Instituto de Biología, UNAM. Mexico. 55 pp. [ Links ]
41. Martínez-Gordillo M, Jiménez J, Cruz R, Juárez E, García R, Cervantes A, Mejía R (2002) Los géneros de la familia Euphorbiaceae en México. Anal. Inst. Biol., Serie Botánica 73: 155-281. [ Links ]
42. Mas JF, Pérez-Vega A (2005) La representatividad del Sistema Nacional de Áreas Protegidas (SINAP). INE-SEMARNAT, México. Gaceta Ecológica 74: 5-14. [ Links ]
43. McVaugh R (1974) Fagaceae. Cont. Univ. Michigan Herb. 12: 1-93. [ Links ]
44. Méndez-Larios I, Villaseñor JL, Lira R, Morrone JJ, Dávila P, Ortiz E (2005) Toward the identification of a core zone in the Tehuacán-Cuicatlán Biosphere Reserve, México, based on parsimony analysis of endemicity of flowering plant species. Interciencia 30: 267-274. [ Links ]
45. Miles L, Newton AC, DeFries RS, Ravilious C, May I, Blyth S, Kapos V, Gordon JE (2006) A global overview of the conservation status of tropical dry forests. J. Biogeog. 33: 491-505. [ Links ]
46. Miranda F, Hernández-X E (1963) Los tipos de vegetación de México y su clasificación. Bol. Soc. Bot. Mex. 28: 29-57. [ Links ]
47. Mittermeir RA (1988) Primate diversity and the tropical forest: case studies from Brazil and Madagascar and the importance of the megadiversity countries. In Wilson EO (Ed) Biodiversity. National Academy Press. Washington, DC, USA. pp. 145-154. [ Links ]
48. Morrone JJ (1994) On the identification of areas of endemism. Syst. Biol. 43: 438-441. [ Links ]
49. Morrone JJ, Crisci JV (1995) Historical biogeography: introduction to methods. Annu. Rev. Ecol. Syst. 26: 373-401. [ Links ]
50. Morrone JJ, Escalante T (2002) Parsimony analysis of endemicity (PAE) of Mexican terrestrial mammals at different area units: When size matters. J. Biogeog. 29: 1095-1104. [ Links ]
51. Morrone JJ, Espinosa D, Aguilar C, Llorente J (1999) Preliminary classification of the mexican biogeographic provinces: A parsimony analysis of endemicity based on plant, insect and bird taxa. Southwest. Naturalist 44: 507-514. [ Links ]
52. Morrone JJ, Espinosa D, Llorente J (2002) Mexican biogeographic provinces: Preliminary scheme, general characterizations, and synonymies. Acta Zool. Mex. 85: 83-108. [ Links ]
53. Müller P (1973) Introducción a la zoogeografía. Blume. Barcelona, Spain. 232 pp. [ Links ]
54. Murphy PG, Lugo AE (1995) Dry forests of Central America and the Caribbean. In Bullock SH, Mooney HA (Eds.) Seasonally dry tropical forests. Cambridge University Press. Cambridge, UK. pp. 9-34. [ Links ]
55. Nixon KC (2002) Winclada ver. 1.00.08. Ithaca, New York, USA. www.cladistics .com/about_winc.htm. [ Links ]
56. Olson DM, Dinerstein E (2002) The Global 200: priority ecoregions for global conservation. Ann. Miss. Bot. Gard. 89: 199-224. [ Links ]
57. Phillips O, Miller JS (2002) Global patterns of plant diversity: Alwyn H. Gentrys forest transect data set. Monogr. Syst.c Bot. Miss. Bot. Gard. 89: 1-319. [ Links ]
58. Platnick N (1991) On the areas of endemism. Aust. Syst. Bot. 4: 11-12. [ Links ]
59. Posadas P, Miranda-Esquivel DR (1999) El PAE (Parsimony Analysis of Endemicity) como una herramienta en la evaluación de la biodiversidad. Rev. Chil. Hist. Nat. 72: 539-546. [ Links ]
60. Rodrigues ASL, Andelman SJ, Bakarr MI, Boitani L, Brooks TM, Cowling RM, Fishpool LDC, da Fonseca GAB, Gaston KJ, Hoffmann M, Long J, Marquet PA, Pilgrim JD, Pressey RL, Schipper J, Sechrest W, Stuart SN, Underhill LG, Waller RW, Watts MEJ, Yan X (2004) Effectiveness of the global protected area network in representing species diversity. Nature 428: 640-643. [ Links ]
61. Rojas O, Alcántara O, Navarro A (2003) Regionalization of the avifauna of the Baja California Peninsula, México: A parsimony analysis of endemicity and distributional modelling approach. J. Biogeog. 30: 449-461. [ Links ]
62. Rosen BR (1988) From fossils to Earth history: Applied historical biogeography. In Myers AA, Giller PS (Eds.) Analytical biogeography: An integrated approach to the study of animal and plant distributions. Chapman and Hall. London, UK. pp. 437-481. [ Links ]
63. Rovito SM, Arroyo M, Pliscoff P (2004) Distributional modeling and parsimony analysis of endemicity of Senecio in the Mediterranean-type climate area of Central Chile. Glob. Ecol. Biogeog. 13: 85-92. [ Links ]
64. Rzedowski J (1978) Vegetación de México. Limusa. México. 432 pp. [ Links ]
65. Rzedowski J (1990) Vegetación Potencial. IV.8.2. Atlas Nacional de México. Vol II. Escala 1:4000000. Instituto de Geografía. UNAM. México. [ Links ]
66. Rzedowski J (1991a) Diversidad y orígenes de la flora fanerogámica de México. Acta Bot. Mex. 14: 3-21. [ Links ]
67. Rzedowski J (1991b) El endemismo en la flora fanerogámica Mexicana: Una apreciación analítica preliminar. Acta Bot. Mex. 15: 47-64. [ Links ]
68. Rzedowski J, Medina L, Calderón de Rzedowski G (2005) Inventario del conocimiento taxonómico, así como de la diversidad y del endemismo regionales de las especies mexicanas de Bursera (Burseraceae). Acta Bot. Mex. 70: 85-111. [ Links ]
69. Santos CM (2005) Parsimony analysis of endemicity: Time for en epitaph? J. Biogeog. 32: 1281-1286. [ Links ]
70. SEMARNAT (2002) Norma Oficial Mexicana NOM-059-ECOL-2001. Protección ambiental-Especies nativas de México de flora y fauna silvestre-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-Lista de especies en riesgo. Secretaría de Medio Ambiente y Recursos Naturales. Diario Oficial de la Federación. 6/3/2002. pp. 2-56. [ Links ]
71. Silva JM, Cardosos M, Castelletti CHM (2004) Areas of endemism for passerine birds in the Atlantic forest, South America. Glob. Ecol. Biogeog. 13: 85-92. [ Links ]
72. Sosa V, Dávila P (1994) Una evaluación del conocimiento florístico de México. Ann. Miss. Bot. Gard. 81: 749-757. [ Links ]
73. Stork NE (1994) Inventories of biodiversity: More than a question of numbers systematics and conservation evaluation. In Forey PL, Humphries CJ, Vane-Wright RI (Eds.) The Systematics Association. Clarendon Press. Oxford, UK. pp. 81-97. [ Links ]
74. Trejo I, Dirzo R (2000) Deforestation of seasonally dry tropical forest: A national and local analysis in México. Biol. Cons. 94: 133-142. [ Links ]
75. Trejo I, Dirzo R (2002) Floristic diversity of Mexican seasonally dry tropical forest. Biodiv. Cons. 11: 2063-2084. [ Links ]
76. Vane-Wright RI, Humphries CJ, Williams PH (1991) What to protect? Systematics and the agony of choice. Biol. Cons. 55: 235-254. [ Links ]
77. Villaseñor JL (2003) Diversidad y distribución de las Magnoliophyta de México. Interciencia 28: 160-167. [ Links ]
78. Villaseñor JL, Ibarra-Manríquez G, Ocaña D (1998) Strategies for the conservation of Asteraceae in México. Cons. Biol. 12: 1066-1075. [ Links ]
79. Villaseñor JL, Meave J, Ortiz-Bermúdez E, Ibarra-Manríquez G (2003) Biogeografía y conservación de los bosques tropicales húmedos de México. En Morrone JJ, Llorente J (Eds.) Una perspectiva Latinoamericana de la biogeografía. UNAM. México. pp. 209-216. [ Links ]
80. Wendt T (1993) Composition, floristic affinities, and origins of the canopy tree flora of the Mexican Atlantic slope rain forest. In Ramamoorthy TP, Bye R, Lot A, Fa J (Eds.) Biological diversity of México: origins and distribution. Oxford University Press, Nueva York. pp. 595-680. [ Links ]
