











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.34 n.2 Caracas feb. 2009
Caracterización del material forestal de reproducción de cinco procedencias de nothofagus alessandrii espinosa, una especie en peligro de extinción.
Rómulo Santelices Moya, Rafael María Navarro Cerrillo y Fernando Drake Aranda
Rómulo Santelices Moya. Magíster en Ciencias Forestales, Universidad de Chile. Profesor, Universidad Católica del Maule (UCM), Chile. Dirección: Departamento de Ciencias Forestales, Facultad de Ciencias Agrarias y Forestales, UCM, Casilla 617, Talca, Chile. e-mail: rsanteli@ucm.cl.
Rafael María Navarro Cerrillo. Doctor Ingeniero de Montes, Universidad Politécnica de Madrid, España. Profesor, Universidad de Córdoba, España.
Fernando Drake Aranda. Doctor en Ciencia Forestal e Ingeniería de Recursos Naturales, Universidad de Córdoba, España. Profesor, Universidad de Concepción, Chile.
RESUMEN
Nothofagus alessandrii es una especie endémica de la zona mesomórfica de Chile que se encuentra en peligro de extinción y de la cual existe aún información insuficiente respecto de las diferencias entre sus poblaciones. Por ello, en este trabajo se analizan y caracterizan parámetros cualitativos y cuantitativos del material germinativo y el comportamiento en vivero de cinco procedencias representativas del área de distribución natural de la especie. Se colectaron semillas de estas procedencias en febrero 2000 y se determinó su tamaño, forma, peso y capacidad germinativa. En septiembre del mismo año se cultivaron plantas en vivero y se evaluó su crecimiento diametral y en altura. Los resultados indican que el tamaño de las semillas es, en general, homogéneo y solo se observan diferencias estadísticas en la longitud de las dímeras y ancho de las trímeras. El peso de 1000 semillas y la capacidad germinativa varió significativamente entre las procedencias. El desarrollo de las plantas en vivero fue homogéneo y no hubo diferencias en el diámetro de cuello de la raíz ni en la altura que alcanzaron las plantas después de una temporada de cultivo. N. alessandrii tiene un comportamiento diferente al de otras especies del mismo género, sin que se observe una variación de tipo clinal. Sin embargo, la procedencia más austral tiende a diferenciarse de las otras y, probablemente, corresponda a un ecotipo diferente.
Characterization of forest reproductive material from five provenances of nothofagus alessandrii espinosa, an endangered species.
SUMMARY
Nothofagus alessandrii is an endemic and endangered species from the mesomorphic zone of Chile. Currently, differences across populations are not well known. To this end, quantitative and qualitative parameters pertaining to germinative material and nursery performance of seedlings from five provenances representing the natural distribution of the species were analyzed and characterized. Seeds were collected in February 2000 and size, form, weight, and germination capacity were determined. Seedlings were then cultivated in nursery in September of the same year, and basal diameter and height growth were quantified. Results indicate that seed size is generally homogeneous across sites, and statistical differences were limited to length of dimerous seeds and width of trimerous seeds. Seed weight and germination capacity varied significantly among the provenances. After one cultivation season, the development of the nursery plants was homogenous across provenances, with no differences in basal diameter or height. N. alessandrii differs, in relation to the variation among the studied provenances, from that of other species of the same genus, with the exception of a pronounced clinal type. However, the southern-most provenance tended, in general, to differ from the other ones and likely represents a different ecotype.
Caracterização do material florestal de reprodução de cinco procedências de nothofagus alessandrii espinosa, uma espécie em perigo de extinção.
RESUMO
Nothofagus alessandrii é uma espécie endêmica da zona mesomórfica do Chile que se encontra em perigo de extinção e da qual existe ainda informação insuficiente em relação às diferenças entre suas populações. Por isto, neste trabalho se analisam e caracterizam parâmetros qualitativos e quantitativos do material germinativo e o comportamento em viveiro de cinco procedências representativas da área de distribuição natural da espécie. Recolheram-se sementes destas procedências em fevereiro de 2000 e se determinou seu tamanho, forma, peso e capacidade germinativa. Em setembro do mesmo ano se cultivaram plantas em viveiro e se avaliou seu crescimento diametral e em altura. Os resultados indicam que o tamanho das sementes é, em geral, homogêneo e somente se observam diferenças estatísticas no comprimento das dímeras e largura das trímeras. O peso de 1.000 sementes e a capacidade germinativa variou significativamente em relação às procedências. O desenvolvimento das plantas em viveiro foi homogêneo e não houve diferenças no diâmetro do colo da raíz nem na altura que alcançaram as plantas depois de uma temporada de cultivo. N. alessandrii tem um comportamento diferente ao de outras espécies do mesmo gênero, sem que se observe uma variação do tipo clinal. No entanto, a procedência mais austral tende a diferenciar-se das outras e, provavelmente, corresponda a um ecótipo diferente.
PALABRAS CLAVE/ Cultivo en Vivero / Nothofagus alessandrii / Procedencias / Ruíl / Semillas /
Recibido: 29/11/2007. Modificado: 28/11/2008. Aceptado: 10/12/2008.
Introducción
Nothofagus alessandrii Espinosa, conocido corrientemente como ruíl, es una especie endémica de la zona central de Chile, declarada oficialmente por el Estado chileno como en peligro de extinción (Benoit, 1989). Tiene un área de distribución natural muy restringida, limitada a la Cordillera de la Costa de la Región del Maule, en una faja que no supera los 100km de extensión latitudinal, variando entre los 100 y los 450msnm (San Martín et al., 1991). Forma parte de la asociación conocida como Bosque Maulino Costero y tiende a formar bosques puros en exposiciones de umbría. Es un árbol con una excelente forma que puede llegar a medir hasta 30m de altura y 1m de diámetro (Muñoz, 1973; Rodríguez et al., 1983; Serra et al., 1986; Rodríguez y Quezada, 2003). Su madera es de reconocida calidad, especialmente por su resistencia a la pudrición (Aravena y Molina, 1976; Donoso, 1983; Fierro y Pancel, 1998). A comienzos del siglo XX su madera fue utilizada en la construcción de embarcaciones conocidas como faluchos maulinos, y en la actualidad aún es posible encontrar en localidades cercanas a su distribución natural estacas de cerco de más de 80 años conservadas en buen estado (Aravena y Molina, 1976; Donoso, 1983). Las propiedades mecánicas son comparables, e incluso algunas superiores, a las de Nothofagus pumilio (Poep. et Endel) Krasser y Nothofagus nervosa (Phil.) Dim. et Mil., las dos especies nativas de Chile más comercializadas en el mercado nacional e internacional (Santelices y Contreras, 2004).
A comienzos del siglo pasado, los bosques fueron objeto de tala y quema (Donoso y Landaeta, 1983). Las poblaciones remanentes se encuentran muy fragmentadas y están rodeadas por una matriz de plantaciones de pino radiata (Pinus radiata D. Don), una especie agresiva con capacidad de invadir con éxito los bosques de ruíl (Bustamante y Grez, 1995; Bustamante y Castor, 1998). Se han contabilizado 185 fragmentos que, en promedio, tienen una superficie de 1,9ha y solo el 5% de ellos supera las 12ha (Bustamante y Grez, 1995). Debido a la situación de esta especie, considerada como el árbol más amenazado de Chile, debería tener una alta prioridad de conservación (UICN, 2001; Hechenleitner et al., 2005), más aún cuando siguen presentes los procesos antropogénicos que la han llevado a su actual estado de degradación. En 12 años, la fragmentación ha aumentado en más de un 2,2%, siendo las actividades del ser humano la principal causa de ello (Olivares et al., 2005).
La conservación de los recursos genéticos depende de la magnitud y del patrón de variación intraespecífica. Por lo general, la determinación de patrones de variación genética en poblaciones de plantas es muy variable e involucra complejas interacciones entre atributos tales como formas de vida, arquitectura floral, formas de reproducción, y factores ambientales y ecológicos que pueden incidir en la polinización, tamaño y aislamiento de la población (Mamo et al., 2006). Estos y otros factores podrían influir en el polimorfismo entre las poblaciones. Existen evidencias de que podrían presentarse diferentes poblaciones de N. alessandrii y que existiría una baja variabilidad genética al interior de los fragmentos, lo que explicaría, por ejemplo, la alta tasa de semillas no viables como consecuencia de un proceso de endogamia (Pineda, 1998). Particularmente, la morfología de las semillas de N. alessandrii determina que el patrón de dispersión sea muy pobre y quede fundamentalmente reducido al medio terrestre aéreo-seco y que se produzca, principalmente, por gravedad (Hill y Jordan, 1993) y en pequeña proporción por el viento. La anemocoría no es eficiente a causa del reducido tamaño de las alas y el obstáculo de árboles vecinos de la matriz (San Martín et al., 2006).
La selección de la procedencia y de la fuente de semilla es de gran importancia en cualquier programa de forestación (Jara, 1995) y es mucho más relevante en una especie amenazada como N. alessandrii. En Chile, el conocimiento que se tiene sobre la variabilidad dentro y entre las poblaciones de la mayoría de las especies nativas es aún escaso, y se puede encontrar en Donoso et al. (2004). Para N. alessandrii es casi nulo, reduciéndose a un trabajo de fin de carrera (Acevedo y Urra, 2002). La mayor información existente, que incluye a la especie en estudio, está restringida a la distribución de tipos forestales, en función de su composición y ubicación geográfica (Donoso, 1993; San Martín et al., 2006). Por consiguiente, estudios que aborden la variabilidad entre procedencias debe ser uno de los requisitos previos para implementar cualquier programa de conservación. Algunos autores señalan la importancia de realizar pruebas que puedan diferenciar la variabilidad asociada a semillas en función de sus orígenes (Mamo et al., 2006) y si bien es cierto que este enfoque está orientado al mejoramiento genético, adquiere una importancia mayor en un programa de conservación. Generar información que permita identificar las características particulares de la semilla en función de su procedencia, debería facilitar los trabajos de conservación y de restauración ecológica. En el caso de N. alessandrii, solo se tiene conocimiento de la distribución espacial de los fragmentos que aún quedan (San Martín y Sepúlveda, 2002; Olivares et al., 2005) y de algunas características de las semillas de sitios puntuales (Olivares et al., 2005; San Martín et al., 2006) sin que se haya realizado un análisis comparativo en forma global. Por ello, estudiar la variabilidad entre las diferentes procedencias es condición necesaria para la gestión y conservación de los fragmentos existentes de esta especie.
El objetivo de este estudio fue analizar y caracterizar parámetros cualitativos y cuantitativos del material forestal de reproducción de cinco procedencias de Nothofagus alessandrii Espinosa y su comportamiento en vivero.
Área de Estudio
Las cúpulas de N. alessandrii fueron cosechadas de cinco procedencias representativas de su distribución natural. La ubicación geográfica y condiciones climáticas de los sitios se presentan en la Tabla I y en la Figura 1. Todos los rodales estudiados se clasifican en el tipo forestal Roble-Hualo, subtipo Bosquetes de ruíl (Donoso, 1993) y se encuentran rodeados de plantaciones de pino radiata.
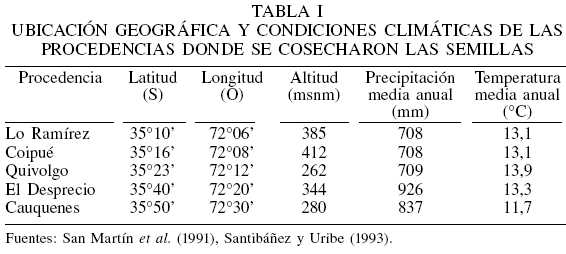
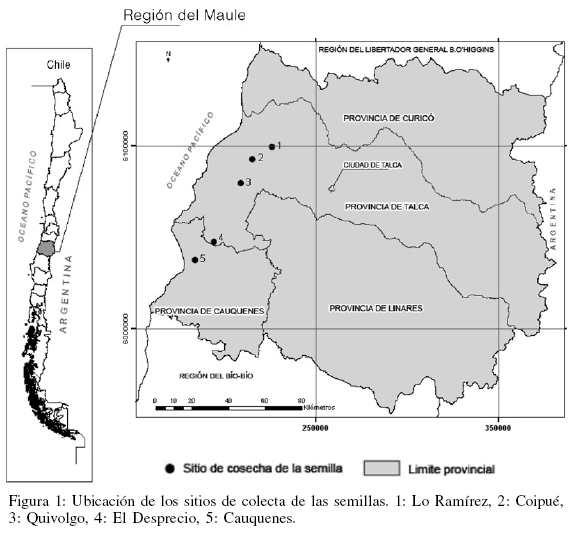
Del área estudiada, los sitios de Lo Ramírez y Coipué representan las distribuciones más septentrionales, mientras que El Desprecio y Cauquenes son las más australes. La geomorfología del área de distribución natural de N. alessandrii integra la columna vertebral de la Cordillera de la Costa, que en la región del Maule presenta un elemento adicional característico, como son las secciones transversales perpendiculares a la dirección de su eje principal. Estas secciones, al canalizar los cuerpos fluviales, representan cuencas que difieren en magnitud, siendo las de mayor importancia las drenadoras de los ríos Mataquito y Maule. Estas cuencas definen dos zonas interfluviales donde se localiza el bosque de N. alessandrii. La primera, entre los ríos Mataquito y Maule y la segunda, entre el río Maule y el Curanilahue (San Martín et al., 2006). Landaeta (1981), siguiendo a Köppen, clasifica el clima para el área de distribución de N. alessandrii como templado cálido con estaciones secas prolongadas e influencia costera (Csb1n) y templado cálido con estaciones secas prolongadas (Csb1). De acuerdo a Hajek y di Castri (1975) la temperatura media es 13,7ºC con máxima en enero de 24,8ºC y mínima en julio de 5,9ºC. La precipitación promedio anual es de 830,7mm, observándose la máxima en junio con 186,4mm. La humedad relativa media anual es 78%, registrándose los menores valores entre noviembre y enero (69%) y la máxima desde abril a julio (86,3%). La amplitud térmica inferior a 20ºC muestra una acentuada termorregulación de origen marino que es reforzada por una fría y húmeda brisa marina del noroeste en invierno, y del suroeste en verano. Esta atenuación térmica es potenciada por neblinas otoñales e invernales que penetran al interior del continente por quebradas y junto con aumentar la humedad contribuyen a disminuir la temperatura ambiental (San Martín et al., 2006).
Los suelos donde se distribuye naturalmente N. alessandri tienen un origen metamórfico con sobreposición de los tipos pardo no cálcicos (dominan en el área interfluvial de los ríos Mataquito y Maule) y los del tipo lateritas pardo rojizos (dominan desde Constitución al sur). Esta disposición imposibilita una definición clara, y son descritos como transicionales entre lateritas pardo rojizos y pardos no cálcicos (Roberts y Díaz, 1959-1960). A este grupo, y en algunos sitios, se interponen otros de origen granítico. Una característica común que tienen es, de acuerdo a San Martín et al. (2006), su baja fertilidad, que se ve potenciada por un bajo contenido de fósforo. Como consecuencia de su antigüedad e influencia climática, son suelos muy intemperizados, delgados, con muy baja retención de agua y susceptibles a la erosión de manto y pluvial. Otro elemento importante es la disposición en altura y en laderas, lo que junto a su sequedad restringe la capacidad de uso al ámbito forestal. De acuerdo a Landaeta (1981) en las capas superiores la textura es franco arcillo arenosa con coloración pardo rojizo oscuro y, por el contrario, a niveles más profundos esta textura cambia a arcillo limosa y de color pardo rojizo a rojo. En general, son suelos con texturas superficiales moderadamente finas con buen drenaje.
Materiales y Métodos
Cosecha y procesamiento de semillas
La cosecha de los frutos se realizó durante la primera quincena de febrero de 2000, de árboles fenotípicamente superiores en cuanto a diámetro, altura, rectitud, forma, desarrollo de la copa y estado sanitario. La cantidad de árboles muestreados por procedencia estuvo sujeta a la fructificación producida en esa temporada (N. alessandrii presenta ciclos de producción de semillas, o semillación, poco conocidos) y varió desde tres (Quivolgo) hasta doce individuos (Lo Ramírez). Se procuró que los árboles estuvieran lo más distanciados entre sí para asegurar el máximo de variación genética. Las cúpulas se cosecharon directamente de las ramas y fueron transportadas al vivero de la Universidad Católica del Maule el mismo día de la colecta. Posteriormente las semillas fueron limpiadas y, con el objeto de mantener su viabilidad, fueron almacenadas en bolsas de papel en un refrigerador convencional a 4°C (Hartmann y Kester, 1998) durante cinco meses.
Características morfo-fisiológicas de las semillas
Con el propósito de evaluar la variabilidad de las características de las semillas se cuantificó su forma y tamaño, así como el peso de 1000 granos, y se determinó el grado de pureza de cada lote. Por cada procedencia, se seleccionaron aleatoriamente cinco muestras de 15 semillas cada una, se separaron las trímeras de las dímeras y, en forma individual, se les midió con un micrómetro digital (Mitutoyo®) longitud, ancho y espesor, dependiendo de su morfología. Para el peso de las semillas, se pesaron en una balanza digital (Ohaus®) diez muestras de 100 semillas cada una (ISTA, 2006), para así estimar el peso de 1000 semillas.
Con las semillas cosechadas se realizó un ensayo de germinación bajo condiciones de invernadero, para lo cual se contó con camas calientes que permitieron mantener el substrato a una temperatura constante de 20°C y riego automático por aspersión (Mistamatic®). Solo se consideraron semillas viables, para lo cual después de una prueba de flotación por 24h se eliminaron las que flotaron (Donoso, 1975). Posterior a la imbibición de agua, se seleccionaron al azar cinco muestras de 50 semillas viables cada una y se sembraron en bandejas de poliestireno expandido expuestas a la luz. El proceso de germinación fue monitoreado diariamente durante 100 días y se consideró exitoso cuando la radícula alcanzó 2mm de longitud. Posterior a este tiempo, se calculó la capacidad germinativa (CG) como el porcentaje de semillas germinadas respecto del total sembradas y se graficó la curva de germinación.
Atributos morfológicos de la planta
Con la finalidad de evaluar el desempeño en vivero de las cinco procedencias seleccionadas y obtener una producción más eficiente, se consideró sembrar las semillas pregerminadas (Landis et al., 1999). Una vez iniciada la germinación, las semillas se sembraron en contenedores estriados de 140ml de capacidad (Termomatrices®) y como substrato se utilizó una mezcla de corteza compostada de pino y perlita 8 (7:3 v/v). El desarrollo de las plantas en vivero de las diferentes procedencias fue evaluado por medio de un ensayo con un diseño experimental en bloques completamente aleatorizados de efectos fijos. Se efectuaron cinco repeticiones por cada tratamiento y se utilizaron 25 plantas por unidad experimental, para un total de 125 plantas por tratamiento.
En el cultivo de las plantas se realizaron las prácticas culturales habituales en vivero. El riego se efectuó mediante microaspersores y un programador de riego, manteniendo el substrato a capacidad de campo. Las plantas se mantuvieron libres de malezas y agentes patógenos; se efectuó un control de malezas manual y para curar la aparición de hongos posterior a la emergencia, en dos oportunidades se aplicó con frecuencia quincenal una mezcla de los fungicidas Strepto plus® (30g·l-1) y Dithane M-45® (15g·l-1) en 30 litros de agua. El substrato fue mezclado con los fertilizantes salitre potásico (2,5kg·m-3 con 16% N y 15% K) y superfosfato triple (2,5kg·m-3 con 20,1% P). En los tres últimos meses las plantas fueron asperjadas dos veces por semana con el abono foliar Bayfolan 250 SL® (30ml en 15 litros de agua). Durante el cultivo, las plantas estuvieron protegidas de la insolación directa con una malla de sombra de 80%, colocada a 1m de los contenedores. Finalizada una temporada de crecimiento en vivero (ocho meses), se evaluó el desarrollo de las plantas midiendo los parámetros morfológicos altura y diámetro de cuello de la raíz.
Análisis de los datos y diseño experimental
Los análisis de varianza (ANOVA) y las comparaciones de medias, se realizaron usando el procedimiento GLM (General Linear Model) del programa estadístico SPSS para Windows v. 15. Con el propósito de normalizar aquellas variables expresadas en porcentaje, antes de efectuar los análisis de varianza se les aplicó una transformación en valores angulares (y'= arcsen p; Ostle, 1992). Los valores medios que presentaron diferencias significativas fueron comparados con la prueba de Tukey al nivel del 5%.
El análisis de varianza para los tratamientos relacionados con el tamaño y peso de la semilla se realizó con un diseño al azar simple de efectos fijos. Se efectuaron cinco réplicas para el tamaño de la semilla y 15 semillas por unidad experimental. En el caso del peso de 1000 semillas se consideraron 10 repeticiones de 100 semillas cada una.
Resultados
Características morfo-fisiológicas de las semillas
El tamaño de las semillas fue en general homogéneo y solo se observaron diferencias significativas en la longitud de las dímeras y ancho de las trímeras (Tabla II). La longitud media de las semillas dímeras fue de 5,5-6,4mm, observándose un mayor tamaño en las de la procedencia Cauquenes. No se registraron diferencias estadísticas en el ancho de las semillas dímeras, pero sí en las trímeras, coincidiendo en este último caso también con la procedencia Cauquenes. El espesor de las semillas dímeras resultó ser muy homogéneo con promedio de 1,24mm. La longitud de las trímeras fue de 5,8-6,1mm. En general, las semillas de la procedencia Cauquenes tienen un mayor tamaño y las de Quivolgo son las más pequeñas.
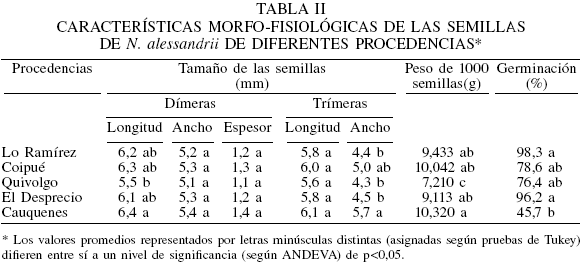
El peso de 1000 semillas varió significativamente entre las procedencias, encontrándose valores medios de 7,21-10,32g (Tabla II). El menor valor correspondió a las semillas provenientes de Quivolgo y los mayores para aquellas de la procedencia de Cauquenes.
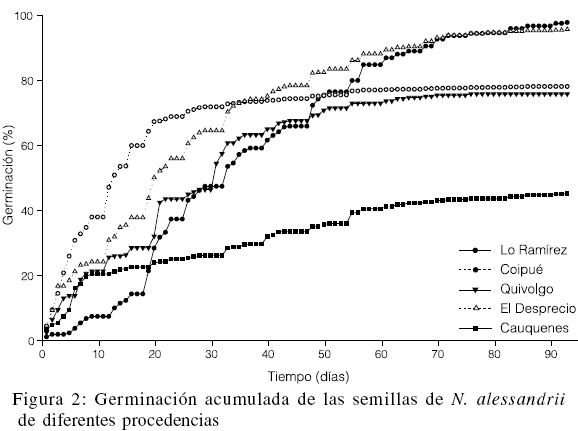
Se observaron diferencias significativas en la capacidad germinativa de las semillas de N. alessandrii (Tabla II). Además, la germinación acumulada en el tiempo varió según la procedencia (Figura 2). La mayor tasa de germinación se observó en las semillas de la procedencia más septentrional (Lo Ramírez), mientras que la más baja correspondió a las semillas de la procedencia más meridional (Cauquenes).
Características de las plantas cultivadas en vivero
El desarrollo de las plantas en vivero fue homogéneo y no se observaron diferencias significativas en su crecimiento en diámetro y altura (Tabla II). Tras ocho meses de cultivo, se registró un promedio de 11,5cm de crecimiento en altura y de 2,5mm en diámetro. Al relacionar estas variables se observa un coeficiente de estabilidad (o pandeo) que va desde 4,2 para la procedencia Cauquenes, hasta 5,1 para las plantas procedentes de Quivolgo.
Discusión
Características morfo-fisiológicas de las semillas
Las características de las semillas pueden ser diferentes como consecuencia de la variación geográfica, pudiendo establecerse marcadas diferencias entre poblaciones (Donahue y Lopez, 1996; Seltmann et al., 2007). La distribución natural de N. alessandrii está restringida a condiciones edáficas y climáticas relativamente homogéneas (San Martín et al., 1991; Santibáñez y Uribe, 1993), lo que debería reflejarse en poblaciones sin grandes variaciones. En este estudio las características de las semillas de N. alessandrii de las cinco procedencias estudiadas no presentan mucha variación, salvo en la longitud de las semillas dímeras y ancho de las trímeras; en cambio, sí se observan diferencias en su peso. El rango observado concuerda con lo señalado por Garrido y Landaeta (1983), quienes encontraron que más de un 84% de las semillas tuvo un diámetro de 4,5-5,5mm. Sin embargo, estos autores no señalan la fuente de semilla, ni discriminan entre dímeras y trímeras.
Se ha observado que en un amplio rango de distribución y diferentes condiciones de sitio, hay variaciones en el calibre de las semillas, existiendo una correlación positiva entre la longitud de ellas y el nivel de precipitación registrado en el sitio de distribución de la población (Mamo et al., 2006). Este tipo de asociación no fue posible observarlo en las procedencias estudiadas, así como tampoco una posible correlación entre el tamaño de las semillas y la latitud.
Al analizar el peso de 1000 semillas, se observan diferencias entre las procedencias, registrándose en las de Cauquenes la mayor masa. Sin embargo, al igual que al evaluar el tamaño de las semillas, no se aprecia una correlación entre el peso de éstas y la variación latitudinal de la procedencia. El rango registrado en este estudio concuerda con lo señalado por varios autores en relación con el número de semillas por kg (Donoso, 1975; Landaeta, 1981; Garrido y Landaeta, 1983; Cabello, 1987; Serra et al., 1986; Mebus, 1993; Espina y Núñez, 1996; Acuña, 2001). Al transformar la información entregada por estos autores, el intervalo observado va desde 10,32 a 7,21g por 1000 semillas. Al comparar una misma procedencia en diferentes temporadas, los resultados varían en el tiempo. Así, por ejemplo, con la información entregada por Landaeta (1981) para la procedencia Lo Ramírez, se registra un peso de 10,3g, valor mayor al resportado en este estudio y a los 8,3g observados por el primer autor (R.S.M.). Estas diferencias deberían estar relacionadas con características genéticas propias de la especie y/o a la modelación ambiental a la que está sujeto el material de reproducción. Los procesos de fructificación y semillación podrían estar influenciados por variaciones climáticas locales. En este sentido, San Martín et al. (2006) señalan que las semillas de árboles de N. alessandrii ubicados en el fondo de quebradas tienen semillas más grandes y pesadas que aquellos en posiciones más altas e iluminadas. En el proceso de recolección de semillas para este estudio, fue posible advertir que la producción se concentró en aquellas partes de la copa de los árboles expuestas a mayor luminosidad, encontrándose árboles con nula producción. En consecuencia, es recomendable estudiar la influencia de factores ambientales como la luminosidad en los procesos de semillación de N. alessandrii.
Aunque no se conocen con exactitud los factores que afectan la ciclicidad del proceso de semillación de N. alessandrii, la producción de semillas varía en el tiempo y ello debiera afectar a su tamaño y a su peso. Otras especies de Nothofagus muestran una producción variable de semillas entre procedencias y entre años (Donoso et al., 2006a; Monks y Kelly, 2006). Marchelli y Gallo (1999), estudiaron la producción de semillas de N. nervosa (Phil.) Dim. et Mil. durante cuatro años, cubriendo toda el área de distribución de esta especie en Argentina, que es pequeña y comparable con este estudio; los autores encontraron diferencias importantes en el peso de las semillas en función de su origen y los años de colecta. En aquellos años de mayor producción las semillas fueron más grandes y pesadas. Además, observaron una variación latitudinal de tipo clinal, encontrando semillas más pesadas en la zona norte de distribución. Este tipo de variación también se ha observado para otros Nothofagus en Chile (Donoso, 1987; Donoso et al., 2006a; González et al., 2006).
Considerando que las características de las semillas observadas en las cinco procedencias de N. alessandrii son en general relativamente homogéneas, y que las condiciones ambientales no varían considerablemente en toda el área de su distribución natural (lo que no favorecería la diferenciación genecológica de sus poblaciones) es probable que la mayoría de las procedencias estudiadas no se diferencien en poblaciones genéticamente diferentes. Sin embargo, se debe tener en cuenta que las semillas de la procedencia más meridional (Cauquenes) tienden a ser diferentes, observándose en ellas un mayor tamaño y peso. Además, esta área geográfica presenta el nivel de precipitación más alto, lo que se contrapone con los estudios realizados con otros Nothofagus (Donoso, 1987; Donoso et al., 2006a, b, c) donde so observó que el tamaño de la semilla disminuía a medida que aumentaba la precipitación. En consecuencia, sería interesante realizar estudios complementarios en este aspecto para N. alessandrii, siendo uno de ellos la caracterización genética entre y dentro de las poblaciones.
Por otra parte, es necesario estudiar la producción de semillas en el tiempo y establecer la duración y el impacto de los ciclos en las características del material forestal de reproducción. En este contexto, el problema que genera la ciclicidad de la semillación, podría solucionarse evaluando la variación de la capacidad germinativa de las semillas con diferentes tratamientos de almacenamiento.
En el presente estudio se observó que la tasa de germinación para la mayoría de las procedencias es en general homogénea y relativamente alta, entre 76,4 y 98,3%, salvo para Cauquenes, que solo supera el 45%. El desarrollo de la germinación fue dispar entre las procedencias y la velocidad de germinación no siempre estuvo asociada a la capacidad germinativa (Figura 2). Así, por ejemplo, la pendiente de la curva de germinación de las semillas de Lo Ramírez es una de las más suaves y con este origen geográfico se obtuvo la mayor tasa de germinación. En cambio, para las semillas de Cauquenes, durante los diez primeros días se observa una alta velocidad de germinación, para luego decrecer y cambiar muy paulatinamente. La mayor rapidez inicial en el proceso de germinación se observa con las semillas de Coipué que, en un lapso de 22 días llegó a registrar una tasa acumulada de germinación de 70%, que varió muy poco hasta el final del ensayo.
No se detectó correlación entre la variación latitudinal y la tasa de germinación de las semillas. Tampoco fue posible asociar la cantidad de semillas producidas con la capacidad germinativa observada de una determinada procedencia. Como se mencionó, en Quivolgo solo fue posible cosechar semillas de tres árboles y, aún así, tuvo una mayor tasa de germinación que Cauquenes. La marcada diferencia registrada en la capacidad germinativa de la procedencia más meridional sugiere estudiar en mayor profundidad este aspecto.
Las tasas de germinación observadas en las procedencias Lo Ramírez y El Desprecio, mayores al 96%, indican que se estaría cerca del máximo de la especie y asegurarían una futura producción de plantas. Esto último, aunque en menor grado, también se visualiza para las de Coipué y Quivolgo. Sin embargo, debido a la ciclicidad en la semillación, estos parámetros no necesariamente se mantendrían al cosechar semilla en diferentes años.
Los valores observados en las tasas de germinación son, a excepción de la procedencia Cauquenes, superiores a los señalados en estudios anteriores. En condiciones experimentales y ex situ, Donoso (1975) con estratificación a 4ºC durante 30-45 días logró una capacidad germinativa de un 47%. Posteriormente Donoso y Cabello (1978), con estratificación a 5ºC durante 20 días y posterior alternancia de temperaturas de 20 y 30ºC, entre 16 y 8h para 30 días, aumentaron la tasa de germinación a 50,7%. Es posible que el mayor porcentaje de germinación alcanzado en este estudio se deba a que se trabajó bajo condiciones de temperatura y humedad controladas en invernadero.
El tamaño de las semillas y en especial su masa, es un aspecto importante en la estrategia reproductiva de muchas especies (Mamo et al., 2006). Sin embargo, en las muestras observadas no se aprecia una correlación entre la masa de las semillas y su capacidad germinativa, lo que sugiere que este aspecto no jugaría un rol importante para predecir la germinación de las semillas de N. alessandrii. Esto último concuerda con lo observado por Close y Wilson (2002) en semillas de Eucalyptus regnans F. Muell y E. delegatensis R.T. Batter.
Al igual que en el análisis de las características de las semillas, las cuatro procedencias más septentrionales presentaron tasas de germinación relativamente homogéneas. En el mismo contexto, la capacidad germinativa de las semillas colectadas en Cauquenes presentó diferencias significativas respecto a las demás y es probable que éstas se deban a factores genéticos. Esto refuerza la necesidad de ahondar los estudios en esta dirección.
Al comparar el peso de las semillas de N. alessandrii con su capacidad germinativa, se observa que no hay correlación entre estas variables. En otros estudios se ha reportado que el tamaño de las semillas de N. dombeyi (Mirb.) Oerst., N. obliqua (Mirb.) Oerst., y N. nervosa disminuye gradualmente de norte a sur y, junto con ello, también lo hace la capacidad germinativa de las semillas, aunque en algunos casos la correlación no es tan clara, como es el caso de N. nervosa (Donoso, 1987; Gallo et al., 2004; Donoso et al., 2006a, b). Esto sugiere la ocurrencia de una variación clinal de tipo genecológico para las especies mencionadas, la cual, por el contrario, en el caso de N. alessandrii no es posible proponer a raíz de los resultados presentados en este estudio.
Plantas cultivadas en vivero
El desarrollo de las plantas en vivero fue similar entre las diversas procedencias. Después de ocho meses de cultivo, se registró un promedio de 11,5cm en altura y 2,5mm en diámetro. Al relacionar estas variables se observa un coeficiente de estabilidad (o pandeo) que va desde 4,2 para la procedencia Cauquenes hasta 5,1 para las plantas procedentes de Quivolgo. Las plantas, a pesar de no tener un gran tamaño, sí presentan gran estabilidad, lo que supuestamente facilitaría su posterior establecimiento en campo.
Los valores de crecimiento registrados se podrían considerar bajos al compararlos con los señalados por otros autores. Por ejemplo, San Martín et al. (2006) reportan crecimientos para plantas cultivadas durante una temporada en la ciudad de Valdivia, en contenedores de 130ml, de alturas superiores a los 35cm y diámetros de cuello de 4-6mm. Por su parte, Fierro y Pancel (1998) indican la posibilidad de producir en una temporada plantas de más de 50cm de altura. Esto sugiere estudiar en mayor profundidad los factores que afectan el desarrollo de las plantas en vivero. En el caso de N. alessandrii, los antecedentes relacionados con la autoecología de la especie (San Martín et al., 2006), permiten suponer que el efecto de la luminosidad, época de siembra y fertilización, o la interacción entre estos factores, podría incidir fuertemente en el desarrollo de las plantas sometidas a cultivo.
Conclusiones
El comportamiento que tiene N. alessandrii, en cuanto a la diferenciación entre las procedencias estudiadas, es diferente al de otras especies del mismo género, sin que se observe una variación de tipo clinal. De las cinco procedencias consideradas en este trabajo, en la más meridional (Cauquenes) se observan, en general, características diferentes en sus semillas y proceso de germinación. En efecto, las semillas tienden a ser más pesadas y de mayor tamaño, pero con una capacidad germinativa significativamente inferior al de las otras razas geográficas. Sin embargo, el crecimiento en vivero fue similar para todas las procedencias. La variabilidad observada sugiere que la procedencia Cauquenes puede ser un ecotipo diferente, siendo necesario profundizar los estudios en este aspecto, particularmente para manejar los fragmentos de N. alessandrii con fines de conservación y restauración.
REFERENCIAS
1. Acevedo S, Urra C (2002) Caracterización de Procedencias en la Etapa de Viverización de Nothofagus alessandrii Espinosa (ruíl) y Nothofagus glauca (Phil.) Krasser (hualo). Tesis. Universidad Católica del Maule. Talca, Chile. 76 pp. [ Links ]
2. Acuña M (2001) Formulación de un Protocolo de Trabajo para el Análisis de Semillas de Especies Leñosas Nativas. Memoria. Facultad de Ciencias Forestales, Universidad de Chile, Santiago, Chile. 87 pp. [ Links ]
3. Aravena P, Molina V (1976) Los Robles-Nothofagus de la Séptima Región de Chile. UC Maule 3: 13-18. [ Links ]
4. Benoit I (Ed.) (1989) Libro Rojo de la Flora Terrestre de Chile (Primera parte). Corporación Nacional Forestal (CONAF). Santiago, Chile. 157 pp. [ Links ]
5. Bustamante R, Castor C (1998) The decline of an endangered ecosystem: the ruil (Nothofagus alessandrii) forest in Central Chile. Biodiv. Cons. 7: 1607-1626. [ Links ]
6. Bustamante R, Grez A (1995) Consecuencias ecológicas de la fragmentación de los bosques nativos. Ambiente y Desarrollo 11: 58-63. [ Links ]
7. Cabello A (1987) Proyecto de Protección y Recuperación de Especies Arbóreas y Arbustivas Amenazadas de Extinción. Partes I y II. Documentos Técnicos 21 y 22. Revista Chile Forestal. Santiago, Chile. 16 pp. [ Links ]
8. Close DC, Wilson SJ (2002) Provenance effects on pre-germination treatments for Eucalyptus regnans and E. delegatensis seed. For. Ecol. Manag. 170: 299-305. [ Links ]
9. Donahue J, López J (1996) Geographic variation in leaf, cone and seed morphology of Pinus greggii in native forests. For. Ecol. Manag. 82: 145-157. [ Links ]
10. Donoso C (1975) Aspectos de la Fenología y Germinación de las Especies de Nothofagus de la Zona Mesomórfica. Boletín Técnico Nº 34, Facultad de Ciencias Forestales, Universidad de Chile, Santiago, Chile. 32 pp. [ Links ]
11. Donoso C (1983) Árboles Nativos de Chile: Guía de Reconocimiento. Marisa Cuneo Ediciones. Valdivia, Chile. 116 pp. [ Links ]
12. Donoso C (1987) Variación natural de Nothofagus en Chile. Bosque 8: 85-97. [ Links ]
13. Donoso C (1993) Bosques Templados de Chile y Argentina. Variación, Estructura y Dinámica. Editorial Universitaria. Santiago, Chile. 484 pp. [ Links ]
14. Donoso C, Cabello A (1978) Antecedentes fenológicos y de germinación de especies leñosas chilenas. Cienc. Forest. 1: 31-40. [ Links ]
15. Donoso C, Landaeta E (1983) Ruil (Nothofagus alessandrii), a threatened Chilean tree species. Env. Cons. 10: 159-162. [ Links ]
16. Donoso C, Premoli A, Gallo L, Ipinza R (Eds.) (2004) Variación Intraespecífica en Especies Arbóreas de los Bosques Templados de Chile y Argentina. Editorial Universitaria. Santiago, Chile. 420 pp. [ Links ]
17. Donoso P, Donoso C, Marchelli P, Gallo L, Escobar B (2006a) Nothofagus nervosa (Phil.) Dim. et Mil. Otros nombres científicos usados: Nothofagus alpina, Nothofagus procera. Raulí. En Donoso C (Ed.) Las Especies Arbóreas de los Bosques Templados de Chile y Argentina, Autoecología. Marisa Cuneo. Valdivia, Chile. pp. 448-461. [ Links ]
18. Donoso P, Donoso C, Gallo L, Navarro C, Escobar B (2006b) Nothofagus dombeyi (Mirb.) Oerst. Coihue, Coigüe. Familia Fagaceae. En Donoso C (Ed.) Las Especies Arbóreas de los Bosques Templados de Chile y Argentina, Autoecología. Marisa Cuneo. Valdivia, Chile. pp. 423-432. [ Links ]
19. Donoso P, Donoso C, Gallo L, Azpilicueta M, Baldini A, Escobar B (2006c) Nothofagus obliqua (Mirb.) Oerst. Roble, Pellín, Hualle. En Donoso C (Ed.) Las Especies Arbóreas de los Bosques Templados de Chile y Argentina, Autoecología. Marisa Cuneo. Valdivia, Chile. pp. 471-485. [ Links ]
20. Espina J, Núñez L (1996) Características germinativas del ruíl (Nothofagus alessandrii Espinosa) para la zona de Curepto. Taller de Habilitación Profesional, Facultad de Ciencias Agrarias y Forestales. Universidad Católica del Maule. Talca, Chile. 42 pp. [ Links ]
21. Fierro S, Pancel L (1998) Experiencia Silvicultural del Bosque Nativo de Chile, recopilación de 57 Especies Arbóreas y Evaluación de Prácticas Silviculturales: Proyecto Manejo Sustentable del Bosque Nativo. Corporación Nacional Forestal (CONAF) y Sociedad Alemana de Cooperación Técnica (GTZ). Santiago, Chile. 420 pp. [ Links ]
22. Gallo L, Donoso C, Marchelli P, Donoso P (2004) Variación en Nothofagus nervosa (Phil.) Dim. et Mil (N. alpina, N. procera). Donoso C, Premoli A, Gallo L, Ipinza R (Eds.) (2004) Variación Intraespecífica en Especies Arbóreas de los Bosques Templados de Chile y Argentina. Editorial Universitaria. Santiago, Chile. pp. 115-144. [ Links ]
23. Garrido F, Landaeta E (1983) Algunos antecedentes sobre el ruíl (Nothofagus alessandrii Espinosa). Cienc. Forest. 3: 3-20. [ Links ]
24. González M, Donoso C, Ovalle P, Martínez-Pastur G (2006) Nothofagus pumilio (Poep. et Endl) Krasser. Lenga, roble blanco, leñar, roble de Tierra del Fuego. Familia: Fagaceae. En Donoso C (Ed.) Las Especies Arbóreas de los Bosques Templados de Chile y Argentina, Autoecología. Marisa Cuneo. Valdivia, Chile. pp. 486-500. [ Links ]
25. Hajek E, di Castri F (1975) Bioclimatografía de Chile. Universidad Católica de Chile. Santiago, Chile. 107 pp. [ Links ]
26. Hartmann H, Kester D (1998) Propagación de Plantas, Principios y Prácticas. Continental. Ciudad de México, México. 760 pp. [ Links ]
27. Hechenleitner P, Gardner M, Thomas P, Echeverría C, Escobar B, Brownless P, Martínez C (2005) Plantas Amenazadas del Centro-sur de Chile. Distribución, Conservación y Propagación. 1ª ed. Universidad Austral de Chile y Real Jardín Botánico de Edimburgo. Valdivia, Chile. 188 pp. [ Links ]
28. Hill R, Jordan G (1993) The evolutionary history of Nothofagus (Nothofagaceae). Australian Systemat. Bot. 6: 111-126. [ Links ]
29. ISTA (2006) International Rules for Seed Testing. Edition 2006. International Seed Testing Association. Bassersdorf, Suiza. [ Links ]
30. Jara L (1995) Mejoramiento Forestal y Conservación de Recursos Genéticos Forestales. Tomo I. Manual técnico Nº 14. CATIE. Turrialba, Costa Rica. 174 pp. [ Links ]
31. Landaeta E (1981) Estudio de las Semillas y Plantas de Vivero para Cuatro Procedencias de Nothofagus alessandrii Espinosa. Tesis. Universidad de Chile. Santiago, Chile. 141 pp. [ Links ]
32. Landis TD, Tinus RW, Barnett JP (1999) The Container Tree Nursery Manual. Vol. 6. Seedling propagation. Agricultural Handbook 674. USDA Forest Service. Washington, DC, EEUU. 166 pp. [ Links ]
33. Mamo N, Mihretu M, Fekadu M, Tigabu M, Teketay D (2006) Variation in seed and germination characteristics among Juniperus procera populations in Ethiopia. For. Ecol. Manag. 225: 320-327. [ Links ]
34. Marchelli P, Gallo LA (1999) Annual and geographic variation in seed traits of Argentinean populations of southern beech Nothofagus nervosa (Phil.) Dim. et Mil. For. Ecol. Manag. 121: 239-250. [ Links ]
35. Mebus I (1993) Enraizamiento en Estacas de Nothofagus Spp. de la Zona Mesomórfica de Chile Amenazadas de Extinción. Tesis. Universidad Austral de Chile. Valdivia, Chile. 68 pp. [ Links ]
36. Monks A, Kelly D (2006) Testing the resource-matching hypothesis in the mast seeding tree Nothofagus truncata (Fagaceae). Austral Ecol. 31: 366-375. [ Links ]
37. Muñoz C (1973) Chile: Plantas en Extinción. Editorial Universitaria. Santiago, Chile. 248 pp. [ Links ]
38. Olivares P, San Martín J, Santelices R (2005) Ruíl (Nothofagus alessandrii): Estado del conocimiento y desafíos para su conservación. Comisión Nacional del Medioambiente (CONAMA). Talca, Chile. 55 pp. [ Links ]
39. Ostle B (1992) Estadística Aplicada. 12ª reimp. Limusa. Ciudad de México, México. 629 pp. [ Links ]
40. Pineda G (1998) Determinación de los Patrones de Variabilidad Genética en Poblaciones de Raulí (Nothofagus alpina (Poepp. et Endl.) y Ruíl (Nothofagus alessandrii Espinosa), por Medio de Electroforesis Horizontal en Geles de Almidón Espinosa), por Medio de Electroforesis Horizontal en Geles de Almidón. Tesis. Universidad de Chile. Santiago, Chile. 79 pp. [ Links ]
41. Roberts R, Díaz C (1959-1960) Los grandes grupos de suelos de Chile. Agric. Técn. 19, 20: 7-36. [ Links ]
42. Rodríguez R, Matthei O, Quezada M (1983) Flora Arbórea de Chile. Universidad de Concepción. Concepción, Chile. 408 pp. [ Links ]
43. Rodríguez R, Quezada M (2003) Fagaceae. Flora de Chile 2: 64-76. [ Links ]
44. San Martín J, Mourgues V, Villa A, Carreño C (1991) Catastro Actualizado De La Distribución y Estado de Conservación de los Bosques de Ruíl en la VII Región. Corporación Nacional Forestal (CONAF). Talca, Chile. 30 pp. [ Links ]
45. San Martín J, Sepúlveda C (2002) Diagnóstico del Estado Actual de los Fragmentos de Nothofagus alessandrii, Ruíl, Fagaceae (= Nothofagaceae), de la Región del Maule, Chile Central. CONAMA. Talca, Chile. 45 pp. [ Links ]
46. San Martín J, Santelices R, Henríquez R (2006) Nothofagus alessandrii Espinosa, Ruíl. Familia: Fagaceae. En Donoso C (Ed.) Las Especies Arbóreas de los Bosques Templados de Chile y Argentina, Autoecología. Marisa Cuneo. Valdivia, Chile. pp. 390-400. [ Links ]
47. Santelices R, Contreras J (2004) Antecedentes dasométricos y de la madera de Nothofagus alessandrii Espinosa. En 2º Cong. Chileno de Ciencias Forestales. Valdivia, Chile. p. 12. [ Links ]
48. Santibáñez F, Uribe J (1993) Atlas Agroclimático de Chile, Regiones VI, VII, VIII y IX. Universidad de Chile. Santiago, Chile. 99 pp. [ Links ]
49. Seltmann P, Leyer I, Renison D, Hensen I (2007) Variation of seed mass and its effects on germination in Polylepis australis: implications for seed collection. New Forests 33: 171-181. [ Links ]
50. Serra MT, Gajardo R, Cabello A (1986) Programa de Protección y Recuperación de la Flora Nativa de Chile: Ficha Técnica de Especies Amenazadas: Nothofagus alessandrii Espinosa "ruíl" (Fagaceae), Especie en Peligro. Corporación Nacional Forestal (CONAF). Santiago, Chile. 32 pp. [ Links ]
51. UICN (2001) Categorías y Criterios de la Lista Roja de la UICN. Ver. 3.1. Comisión de Supervivencia de Especies. Unión Internacional para la Conservación de la Naturaleza y los Recursos Naturales. Gand, Suiza y Cambridge, RU. 33 pp. [ Links ]
