











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.34 n.10 Caracas oct. 2009
Relaciones entre anatomía foliar, intercambio de gases y crecimiento en juveniles de cuatro especies forestales
Osmary Araque, Ramón E. Jaimez, Carmen Azócar, Wilmer Espinoza y Wilmer Tezara
Osmary Araque. Ingeniero Forestal y M.Sc. en Manejo de Bosques, Universidad de Los Andes (ULA), Venezuela. Proyecto Fortalecimiento de la Ruta del Chocolate. Instituto de Investigaciones Agropecuarias. Dirección: Instituto de Investigaciones Agropecuarias. ULA. Apartado 77, La Hechicera, Mérida, 5101-A. Venezuela. e-mail: oaraque@cantv.net.
Ramón E. Jaimez. Licenciado en Biología, M.Sc. en Agroforestería y Doctor en Ecología Tropical, ULA, Venezuela. Profesor ULA, Venezuela. e-mail: rjaimez@ula.ve.
Carmen Azócar. Licenciada en Biología, y estudiante del Postgrado en Ecología Tropical, ULA, Venezuela. e-mail: carmenaz@ula.ve.
Wilmer Espinoza. Técnico Agropecuario, ULA, Venezuela. Técnico, Instituto de Investigaciones Agropecuarias, Mérida, Venezuela. e-mail: wilmerespinozab@hotmail.com.
Wilmer Tezara. Licenciado en Biología y Doctor en Ciencias mención Botánica, Universidad Central de Venezuela (UCV). Profesor, UCV, Venezuela. e-mail: wilmer.tezara@ciens.ucv.ve.
RESUMEN
Este estudio tuvo por objetivo relacionar la estructura anatómica foliar con las características ecofisiológicas de Cordia thaisiana (Boraginaceae), Cedrela odorata y Swietenia macrophylla (Meliaceae) y Tabebuia rosea (Bignoniaceae) durante la fase de establecimiento (<2 años), cultivadas en una región al Sur del Lago de Maracaibo, Venezuela. Se realizaron cortes transversales de las láminas foliares, tinciones, mediciones de intercambio gaseoso (CO2 y H2O) y mediciones de crecimiento. Todas las especies presentaron hojas hipoestomáticas, con epidermis uniestratificada en ambas caras, mesófilo bifacial, parénquima esponjoso medianamente laxo y estomas anomocíticos. Se detectaron diferencias significativas para las variables número de estomas/mm2, espesores de lámina foliar, parénquima en empalizada, parénquima esponjoso y epidermis. La mayor tasa de asimilación (A) de CO2 ocurrió en C. odorata y la menor en C. thaisiana. Esta última especie presentó los mejores rendimientos en crecimiento diametral y longitudinal, y, en consecuencia, es la especie más apta para integrar plantaciones agroforestales en el sitio estudiado.
Relations between foliar anatomy, gas exchange and growth in juveniles of four forest species
SUMMARY
The aim of the study was to compare anatomical structure of the leaves and ecophysiological traits of the species Cordia thaisiana (Boraginaceae), Cedrela odorata, Swietenia macrophylla (Meliaceae) and Tabebuia rosea (Bignoniaceae) cultivated in the South of the Maracaibo Lake, Venezuela. Transverse leaf sections were prepared and stained, and gas exchange (CO2 and H2O) and growth were measured. All the species had hypostomatics leaves with monostratified epidermis at both surfaces, bifacial mesophyll moderately lax spongy parenchyma, and anomocytic stomata. Significant differences were found for the number of stomata/mm2 and leaf thickness, as well as palisade, spongy parenchyma and epidermis thicknesses. The gas exchange parameters showed that C. odorata has the highest photosynthetic rate (A) and C. thaisiana the lowest rate. The latter species presented better yields, with higher diametrical and length growth, being thus recommended for agroforestry plantations in the studied site.
Relações entre anatomia foliar, intercâmbio de gases e crescimento em juvenis de quatro espécies florestais
RESUMO
Este estudo teve por objetivo relacionar a estrutura anatômica foliar com as características ecofisiológicas de Cordia thaisiana (Boraginaceae), Cedrela odorata e Swietenia macrophylla (Meliaceae) e Tabebuia rosea (Bignoniaceae) durante a fase de estabelecimento (<2 anos), cultivadas em uma região ao Sul do Lago de Maracaibo, Venezuela. Realizaram-se cortes transversais das lâminas foliares, tinção, medições de intercâmbio gasoso (CO2 e H2O) e medições de crescimento. Todas as espécies apresentaram folhas hipoestomáticas, com epiderme uniestratificada em ambas caras, mesófilo bifacial, parênquima esponjoso medianamente laxo e estomatos anomocíticos. Detectaram-se diferenças significativas para as variáveis número de estomas/mm2, espessores de lâmina foliar, parênquima em empalizada, parênquima esponjoso e epiderme. A maior taxa de assimilação (A) de CO2 ocorreu em C. odorata e a menor em C. thaisiana. Esta última espécie apresentou os melhores rendimentos em crescimento diametral e longitudinal, e, em consequência, é a espécie mais apta para integrar plantações agroflorestais na área estudada.
PALABRAS CLAVE / Anatomía Foliar / Cedrela odorata / Cordia thaisiana / Intercambio Gaseoso / Swietenia macrophylla / Tabebuia rosea /
Recibido: 11/11/2008. Modificado: 16/09/2009. Aceptado: 17/09/2009.
Introducción
La fase de establecimiento de árboles a campo abierto, especialmente durante los dos primeros años, es un período crítico porque durante ese lapso ejecutan estrategias de adaptación a las condiciones edafoclimáticas del sitio. En este primer estadio ocurren cambios especialmente en las características foliares, los cuales influyen en las tasas de crecimiento (Cornelissen et al., 1996). Las características ambientales (radiación, humedad relativa, temperatura y disponibilidad de agua) donde se desarrolla una planta actúan directamente sobre la estructura anatómica foliar (Fanh, 1990; Lindorf, 1980, 1992; Castro et al., 1982; Ragonese, 1990; Roth, 1990; Flores-Vinda, 1999; Jáuregui y Cardozo, 2000; Ely et al., 2005). Se han definido diferentes tipos de hojas: xeromorfa, mesomorfa e higromorfa, determinándose características propias para cada tipología (Lindorf, 1994; 1998)
Son pocos los estudios de anatomía foliar en especies con madera de alto valor comercial, y menos aun su relación con parámetros ecofisiológicos. Los cambios en la anatomía foliar pueden causar variaciones en el intercambio de gases (CO2, H2O), concentraciones de nitrógeno y eficiencia en el uso de agua (EUA) (Dornhoff y Shibles, 1976; Garnier y Laurent, 1994; Romero-Aranda et al., 1997; Niinemets, 1999; Castro-Díez et al., 2000; Mediavilla et al., 2001; Rengifo et al., 2002; Colombo et al., 2007), lo cual podría explicar las estrategias adaptativas que una especie pueda tener ante determinadas condiciones ambientales.
En regiones tropicales situadas por debajo de los 500msnm, las altas radiaciones, periodos de sequía, textura y condiciones nutricionales de los suelos, además de algunos factores bióticos, influyen en el desarrollo y crecimiento de los árboles, especialmente durante la fase de establecimiento. Bajo estas condiciones surge la interrogante de si las características anatómicas foliares de las especies forestales pueden ser una ventaja adaptativa y una respuesta, con respecto a la fotosíntesis, que influyen en el crecimiento de especies forestales.
Tomando en cuenta la importancia económica de las especies C. odorata, C. thaisiana, S. macrophylla y T. rosea, el objetivo del presente trabajo fue relacionar sus características anatómicas foliares con parámetros ecofisiológicos y de crecimiento, a fin de conocer posibles estrategias de adaptación en la fase de establecimiento en condiciones de campo, y dar recomendaciones de manejo en el sitio estudiado.
Materiales y Métodos
Material vegetal y diseño experimental
El estudio se realizó en el marco del ensayo Proyecto Sistemas Agroforestales con énfasis en cacao, conducido en la Finca La Judibana de la Universidad de Los Andes, localizada en el Municipio Alberto Adriani, El Vigía, estado Mérida, Venezuela (8°3726"N y 71°4222"O) a una altura de 64-73msnm. La temperatura, humedad relativa y precipitación promedio anual en la zona son 26,8°C, 80% y 1834mm, respectivamente (Urbano et al., 2002). Los suelos poseen textura entre franco, franco-arenoso y arenoso; pH ligera a moderadamente ácido; contenido de materia orgánica de bajo a medio y contenidos de N y P de bajos a muy bajos. Plántulas de Cordia thaisiana (Boraginaceae), Cedrela odorata y Swietenia macrophylla (Meliaceae) y Tabebuia rosea (Bignoniaceae) provenientes de viveros de la estación El Irel (estado Barinas) fueron sembradas entre enero y marzo 2007 en un diseño agroforestal de bloques con parcelas al azar con tres repeticiones. Cada parcela consta de nueve árboles separados por 12m entre hileras y 9m entre filas. En base a las condiciones nutricionales del suelo se realizaron dos fertilizaciones con NPK, la primera a finales de mayo 2007 con 15 gr/planta de la formula 15-15-15 y la segunda en noviembre aplicando 85 gr/planta de 18-18-18.
Anatomía foliar
El material de estudio correspondió a muestras de hojas expuestas, maduras y sanas de tres individuos de diez meses de edad, con características fenotípicas (tamaño, vigor y estado fitosanitario) semejantes. De las especies C. odorata, S. macrophylla y T. rosea, que poseen hojas compuestas, se colectaron los foliolos centrales. Las muestras de fueron colocadas en FAA (formaldehído-ácido acético-alcohol etílico). En la preparación del material para el estudio microscópico se siguieron procedimientos estándar sugeridos por Johansen (1940). Los cortes se realizaron en un micrótomo de rotación (American Optical 820, Spencer, EEUU) a un grosor de 20µm. Para el estudio de epidermis y estomas, las muestras fueron obtenidas remojando porciones de lamina foliar en hipoclorito de sodio al 4% por 48-72h y luego se separó las capas epidérmicas del mesófilo, las cuales fueron sometidas a deshidratación, tinción y montaje. Para cada carácter evaluado cuantitativamente se realizaron veinte mediciones. Se tomaron fotomicrografías de las epidermis, corte transversal y vena media para cada especie con una cámara Pentax SFXn acoplada a un microscopio binocular (Kyowa, Medilux 12, Japón).
Intercambio gaseoso
La tasa de asimilación de CO2 (A), transpiración (E) y conductancia estomática foliar (gs) fueron medidas en época de lluvia entre las 11:00 y 13:00, con una densidad de flujo de fotones (DFF) entre 1400-1600μmol·m-2·s-1 en cinco días diferentes entre agosto 2007 y abril 2008, para lo cual se empleó un sistema portátil de intercambio gaseoso (LCA-4, ADC, Hoddesdon, Herts, RU). Se escogieron cuatro árboles de 10 meses de edad por especie, ubicados en parcelas distintas, y en cada uno de ellos se realizaron dos mediciones. El área foliar (AF) se midió con un escáner (CID, CL-202, EEUU) y se determinó el peso seco de cada hoja o foliolo, según el caso, para luego estimar el área foliar específica (AFE). La medición del índice de área foliar (IAF) se llevó a cabo con un analizador de dosel (LAI-2000, LI-COR, Nebraska, EEUU) a los 18 meses, tomando nueve árboles por especie. La A estandarizada se calculó, mediante la ecuación A estandarizada= A´IAF; con el fin de estimar la tasa de asimilación de CO2 potencial alcanzada por un individuo, considerando que todas las hojas tienen igual A instantánea.
Parámetros de crecimiento
Las variables analizadas fueron el diámetro basal a 5cm del suelo y la altura total a los 7, 12 y 18 meses después del trasplante, para lo cual se seleccionaron los árboles centrales de cada parcela de los bloques 2 y 3, totalizando 8 individuos por especie.
Análisis estadístico
Se realizaron análisis de varianza (a=0,05) y pruebas de diferencia de medias (Tukey) empleando el programa estadístico SPSS 13.0 para Windows, sobre los datos correspondientes a espesores de lámina foliar, parénquima en empalizada, parénquima esponjoso, epidermis adaxial y abaxial, número de estomas/mm2, intercambio gaseoso, variables de crecimiento e IAF. Para cada variable se reportan promedios ±error estándar.
Resultados
Las especies estudiadas presentaron características tanto de tipología mesomórficas: epidermis uniestratificada en ambas caras con paredes delgadas, ausencia de hipodermis, estomas solo en la cara abaxial (hipoestomáticas), mesófilo bifacial, parénquima en empalizada alargado, tricomas en ambas caras; como de tipología xeromórfica: haces vasculares rodeados por vainas extendidas, alta densidad estomática (>300est/mm2), cutícula gruesa y pelos peltados en T. rosea, mientras que en C. odorata y S. macrophylla hay presencia de células esclerenquemáticas.
Se encontraron diferencias significativas (a=0,05) entre especies (Tabla I) en la densidad estomática, espesor de lámina foliar, espesor de parénquima en empalizada, espesor de parénquima esponjoso y espesor de epidermis. La mayor densidad estomática se observó en S. macrophylla, mientras que C. thaisiana presentó el mayor espesor en la lámina foliar, parénquima en empalizada y en ambas epidermis.
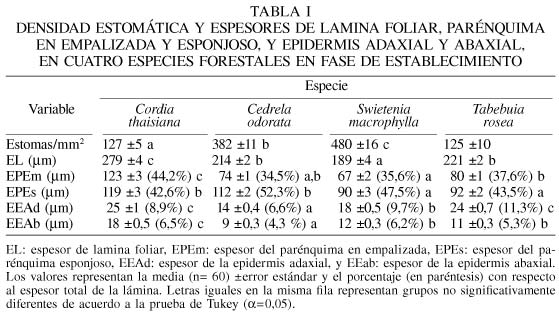
En cuanto a las proporciones de los tejidos epidérmicos y del parénquima, con respecto al total de la lamina foliar se observó que C. thaisiana presenta el mayor porcentaje de parénquima en empalizada, exhibiendo entre 2-5 corridas de células, mientras que el resto de las especies presentan 1-2 corridas (Figura 1). C. odorata presentó el parénquima esponjoso de mayor espesor (Tabla I).

No se encontraron diferencias significativas entre especies para los valores de A, E y gs (Tabla II). T. rosea tuvo el mayor AFE y S. macrophylla el menor. Con relación al IAF, las especies se dividen en dos grupos: C. thaisiana y C. odorata con valores >2, mientras que el grupo de S. macrophylla y T. rosea presentaron valores ~1,6. Las especies mostraron diferencias significativas en las variables de crecimiento. Los mayores valores de diámetro y altura a los 18 meses de edad se registraron en C. thaisiana y C. odorata.

Existe una relación lineal positiva entre los IAF y las dos variables de crecimiento (Figura 2), indicando que las variaciones en diámetro y altura de los árboles pudieran ser atribuidas en 97 y 99%, respectivamente, a los cambios registrados en IAF. De igual forma, se observó una alta relación lineal entre diámetro y altura y la A estandarizada (Figura 3).


Discusión
La conformación del mesófilo bifacial y la combinación de características meso y xeromórficas presente en las cuatro especies es típica de plantas adaptadas a altas condiciones de luz y alta disponibilidad de agua (Smith et al., 1997), que son las condiciones predominantes durante casi todo el año en la zona de estudio. Sin embargo, es posible, debido al alto porcentaje de arena del suelo (>40% hasta los 40cm de profundidad) encontrar condiciones de déficit hídrico muy rápidamente en periodos de poca precipitación.
El parénquima esponjoso medianamente laxo existente en todas las especies puede explicarse como un mecanismo de adaptación a cambios diarios de temperatura foliar que pueden oscilar entre 25 y 38°C (Ragonese, 1990; Ely et al., 2005). La presencia de indumento en ambas epidermis es común en tres de las cuatro especies estudiadas. C. odorata y S. macrophylla muestran pelos simples y pluricelulares, y T. rosea pelos peltados. Estos últimos pueden cumplir una función de absorción y depósito de agua más que de secreción (Ely et al., 2005). Solo dos especies exhiben cutícula en la cara adaxial, la de C. odorata es delgada y papilosa, mientras que la de T. rosea es gruesa y estriada, y puede cumplir la función de reflejar el exceso de luz sobre la lámina foliar (Ely et al., 2005).
Las vainas que rodean los haces vasculares presentes en todas las especies y que en algunos casos se orientan hacia ambas caras de la lámina foliar podrían actuar como una estructura de protección de los haces en situaciones de alta luminosidad. Junto con la presencia de células esclerenquimáticas, también existentes en todas las especies, puede considerarse como un mecanismo para el almacenamiento de agua, ayudando a incrementar la conducción en el mesófilo; además la abundancia de este tejido cumple funciones de soporte en la hoja cuando hay pérdida excesiva de agua (Esau, 1976; Jáuregui y Cardozo, 2000; Ely et al., 2005).
La presencia de cistolitos (carbonato de Ca) en C. thaisiana, combinado con algún terpenoide que según Chen et al. (1983) se extrae de la hoja de esta especie y se usa como repelente de insectos, específicamente de hormigas, puede ofrecer una resistencia al ataque de insectos, ya que en la zona estudiada esta es la única especie que no ha sido atacada por hormigas ni otro tipo de insectos. Esto puede incrementar las posibilidades de éxito en el establecimiento de esta especie en las condiciones bajo estudio. C. odorata amerita tratamientos biológicos y químicos para el control del insecto Hypsiphylla grandella (barrenador de las meliáceas), cuyo ataque limita el crecimiento (Jardim et al, 2004; Lozada y Graterol, 2003) e incrementa los costos de mantenimiento.
Las tasas de A no mostraron diferencias significativas entre especies; sin embargo, la mayor correspondió a C. odorata (11,7) y la menor a C. thaisiana (9μmol·m-2·s-1). Los valores de A reportados por López et al. (2001) son similares para T. rosea, mientras que las tasas obtenidas para C. odorata y C. thaisiana (13,8 y 14μmol·m-2·s-1, respectivamente) son superiores a las encontrados en el presente estudio. Los menores valores pueden estar asociados a las temperaturas registradas durante los días de medición (30-33ºC), ya que López et al. (2001) determinaron que un aumento de la temperatura (>32ºC) trae consigo disminución en la A. De igual forma, al comparar con los resultados obtenidos en Europa por Mediavilla et al. (2001) en especies maderables tanto deciduas como siempre verdes (Crataegus monogyna, Pyrus bourgaeana, Quercus faginea, Q. pyrenaica, Q. rotundifolia, Q. suber) las tasas son relativamente superiores (11-16μmol·m-2·s-1) a las registradas en el presente trabajo, excepto para C. odorata.
En condiciones tropicales las especies siempreverdes como C. thaisiana muestran menores A y AFE, diferencia que ya se presenta en el primer año de establecimiento y que puede estar relacionada con una mayor resistencia física en periodos no favorables para la supervivencia, tales como momentos de alto déficit hídrico o ataque de insectos (Castro-Diez et al., 2000). Sin embargo, esta relación no se mantiene para el resto de las especies. No se observó relación entre la densidad estomática y las variables de intercambio gaseoso, lo cual resulta contrario a lo reportado en otras especies cultivadas (Aguirre et al., 2002) y en arbustos y gramíneas (Rengifo et al., 2002). En cuanto a la relación entre la estructura anatómica foliar y variables ecofisiologicas, Romero-Aranda et al. (1997) en cítricas y Mediavilla et al. (2001) en árboles de zonas templadas encuentran que a mayor espesor en la lámina o menor área foliar especifica (AFE) hay una tendencia a menores tasas de A. Pachepsky y Acock (1998) desarrollaron un modelo teórico que relaciona anatomía foliar con intercambio de gases, y reportan simulaciones donde la tasa de A disminuye a medida que el grosor de la hoja aumenta. Las especies C. thaisiana, C. odorata y T. rosea muestran esta tendencia, mientras que en S. macrophylla se presentan tanto las tasas de A como los espesores de hoja más bajos. Aparentemente no hay una relación entre las tasas de transpiración y la estructura anatómica, lo cual coincide con el planteamiento de Pachepsky y Acock (1998).
Las altas regresiones positivas encontradas ente el diámetro y la altura con el IAF al año y medio de establecimiento indican que el IAF pudiera usarse como indicador del crecimiento en los primeros estadios. Resulta contradictorio que C. thaisiana, a pesar de tener los menores valores de A, presenta las mayores tasa de crecimiento, tanto en altura como en diámetro. Sin embargo, al calcular la tasa de A estandarizada total por individuo, se observa una relación directa (R= 0,89) con las variables de crecimiento. Esto sugiere una mayor superficie de captación de luz utilizable para procesos fotosintéticos y una mayor cantidad de carbohidratos destinada al crecimiento, al menos durante la fase de establecimiento. No obstante, para poder corroborar este planteamiento, es necesario realizar evaluaciones adicionales de estos parámetros, en la medida que los árboles crecen.
Conclusiones
Solamente se encontró una relación inversa entre el espesor de la lámina foliar y las tasas de asimilación de CO2. El crecimiento que presentan las especies puede explicarse en gran parte al relacionar el IAF con la A estandarizada; por lo tanto, en el periodo evaluado, C. thaisiana y C. odorata parecen ser las especies más exitosas y las que pudieran ser recomendadas para la zona en función del crecimiento obtenido.
Agradecimientos
Los autores agradecen a Williams J. León H., Jorge Parra M., Luis E. Gámez A., Luis Cedeño y Wuilson Ramírez, Facultad de Ciencias Forestales y Ambientales, Universidad de Los Andes, Venezuela, y al personal de la finca La Judibana por su apoyo logístico y acertadas observaciones durante el desarrollo de la investigación. Este estudio fue financiado por el MCT, Sub-proyecto 5 del Proyecto en red de la "Ruta del Chocolate" N° 200500898.
Referencias
1. Aguirre J, Acosta J, Ruiz L, Kohashi J, Trejo C (2002) Diferencias morfológicas de la epidermis de la hoja de frijol común y su relación con tolerancia a la sequia. Agric. Téc. Méx. 28: 53-64. [ Links ]
2. Castro-Diez P, Puyravaud J, Cornelissen J (2000) Leaf structure and anatomy as related to leaf mass per area variation in seedlings of a wide range of woody plant species and types. Oecología 124: 476-486. [ Links ]
3. Castro R, Lindorf H, Morillo G (1982) Comparación Anatómica entre diferentes especies del género Matelea (Asclepiadaceae) los cuales crecen en ambiente de bosque húmedo y bosque seco. Mem. Soc. Cien. Nat. La Salle 117: 9-31. [ Links ]
4. Chen T, Ales D, Baenziger N, Wiemer D (1983) Ant-repellent triterpenoids from Cordia alliodora. J. Org. Chem. 48: 3525-3531. [ Links ]
5. Colombo R, Marin O, Irazabal S, Tezara W (2007) Relaciones Hídricas, Fotosíntesis y Anatomía Foliar de dos especies del Género Calotropis. Interciencia 32: 791-796. [ Links ]
6. Cornelissen JHP, Castro-Diez P, Hunt R (1996) Seedling growth, allocation and leaf attributes in a wide range of woody plant species and types. J. Ecol. 84: 755-765. [ Links ]
7. Dornhoff G, Shibles R (1976) Leaf morphology and anatomy in relations to CO2 exchange rate of soybean leaves. Crop Sci. 16: 377-381. [ Links ]
8. Ely F, Torres F, Gaviria J (2005) Relación entre la morfoanatomía foliar de tres especies de Miconia (Melastomataceae) con su hábitat y distribución altitudinal en el Parque Nacional Sierra Nevada de Mérida, Venezuela. Acta Bot. Venez. 29: 275-300. [ Links ]
9. Esau K (1976) Anatomía Vegetal. Omega. Barcelona, España. 779 pp. [ Links ]
10. Fahn A (1990) Plant Anatomy. 4a ed. Pergamon. Totonto, Canadá. 558 pp. [ Links ]
11. Flores-Vinda E (1999) La Planta, Estructura y Función. Vol. II. Tecnológica de Costa Rica. San José, Costa Rica. 834 pp. [ Links ]
12. Garnier E, Laurent G (1994) Leaf anatomy, specific mass and water content in congeneric annual and perennial grass species. New Physiol. 128: 725-736. [ Links ]
13. Jardim F, Ribeiro J, Rosa L (2004) Avaliação preliminar de sistema agroflorestal no Projeto Agua Verde, Albrás, Barcarena, Pará-I. Rev. Ciên. Agr. 41: 25-46. [ Links ]
14. Jauregui D, Cardozo A (2000) Anatomía foliar de dos especies de Chrysobalanaceae presentes en el Parque Nacional Henry Pittier. Acta Bot. Venez. 23: 9-18. [ Links ]
15. Johansen, D (1940) Plant Microtechnique. McGraw Hill. Nueva Cork, EEUU. 523 pp. [ Links ]
16. Lindorf H (1998) Correlaciones eco-anatómicas entre la madera y la hoja. Mem. Inst. Biol. Exp. 1: 209-212. [ Links ]
17. Lindorf H (1994) Eco-anatomical wood features of species from very dry tropical forest. IAWA Journal 15: 361-376. [ Links ]
18. Lindorf H (1992) Anatomía Foliar de especies de un bosque húmedo en el Territorio Federal de Amazonas, Venezuela. Mem. Soc. Cien. Nat. La Salle 52: 65-92. [ Links ]
19. Lindorf H (1980) Estructura foliar de quince monocotiledóneas de sombra del bosque nublado Rancho Grande I: Bifaciales. Araceae, Maranthaceae y Musaceae. Mem. Soc. Cien. Nat. La Salle 113: 19-71. [ Links ]
20. López J, Riaño N, López Y (2001) Características fotosintéticas de cinco especies forestales. Cenicafé 52: 191-169. [ Links ]
21. Lozada J, Graterol D (2003) Practicas agroforestales en el Municipio Rosario de Perijá, estado Zulia, Venezuela. Rev. Fores. Latinoam. 33: 21-36. [ Links ]
22. Mediavilla S, Escudero A, Heilmeier H (2001) Internal leaf anatomy and photosynthetic resource-use efficiency: interspecific and intraspecific comparison. Tree Physiol. 21: 251-259. [ Links ]
23. Niinemets U (1999) Components of leaf dry mass per area-thickness and density-alter leaf photosynthetic capacity in reverse directions in woody plants. Research review. New Physiol. 144: 35-47. [ Links ]
24. Pachepsky LB, Acock B (1998) Effect of leaf anatomy on hypostomatous leaf gas exchange: a theoretical study with the 2Dleaf model. Biotronics 27: 1-14. [ Links ]
25. Ragonese A (1990) Caracteres xeromorfos foliares de Nassauvia lagascae (Compositae). Darwiniana 30: 1-10. [ Links ]
26. Roth I (1990) Leaf structure of a Venezuela cloud forest in relation to the microclimate. Encyclopedia of Plant Anatomy XIV, 1. Berlín, Alemania. [ Links ]
27. Rengifo E, Urich R, Herrrera A (2002) Water relations and leaf anatomy of the tropical species Jatropa gossypifolia and Alternanthera crucis, grown under an elevated CO2 concentration. Photosyntetica 40: 397-403. [ Links ]
28. Romero-Aranda R, Bondada B, Syvertsen J, Grosser J (1997) Characteristics and Net Gas Exchange of Diploid and Autotetrapoid Citrus. Ann. Bot. 79: 153-160. [ Links ]
29. Smith WK, Vogelmann TC, DeLucia EH, Bell DT, Shepherd A (1997) Leaf form and photosynthesis. BioScience 47: 785-793. [ Links ]
30. Urbano D, Dávila C, Saavedra S, Rivas N, Morantes W (2002) Impacto económico sobre producción de leche en un sistema de pastoreo con asociaciones gramíneas-leguminosas arbóreas en el Sur del Lago de Maracaibo, Venezuela. Arch. Latinoam. Prod. Anim. 10: 102-110. [ Links ]
