











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista del Instituto Nacional de Higiene Rafael Rangel
versión impresa ISSN 0798-0477
INHRR v.39 n.2 Caracas dic. 2008
El tiempo, los genes y la historia (Parte III).
Time, genes and history (Part III).
Carlos Aponte*
* Coordinador de Investigación, Gerencia de Docencia e Investigación.
El azar parece conjugarse con el tiempo para jugar un papel importante en la construcción de lo viviente. Así, la evolución y la complejidad biológica pa re cen ser el producto de mutaciones aleatorias variaciones, como lo llamó Darwin) que se producen en la estructura genética de los organismos. Es decir, la aparente direccionalidad de la evolución biológica se encontraría comprometida si asumimos que la espectacular aparición de la vida y su devenir sobre el planeta Tierra es el resultado de un accidente, del azar, de la contingencia.
El concepto de accidente en el origen de la vida y la no-direccionalidad de la evolución biológica, no exclu yen la existencia de una historia de las especies existentes y extintas. La asunción de que todas las formas vivientes sobre la Tierra son descendientes de una o unas pocas formas originales, y que los descendientes tienden a divergir, unos a otros, continuamente de esas formas originales, convirtiendo a estas últimas en los de nominados ancestros comunes, dota a las especies de una historia. Las características variantes presentes en los organismos divergentes tienden a ser heredables. Esta dinámica, molecularmente hablando, está inscrita en un sistema de copiado altamente eficiente (si excluimos a los virus ARN) de la doble cadena de Ácido Desoxirribonucleico (ADN). Así, la posición y orden de los pares de bases nitrogenadas (Citosina-Guanina y Adenina-Timina) a lo largo del interior hidrofóbico de la doble hélice define la secuencia de letras, y por ende de genes, que actuarán como templado durante la replicación del ADN.
Los genes son las unidades físicas y funcionales fundamentales de la herencia. Son segmentos de ácido nucleico que definen una secuencia específica y un número dado de unidades químicas nitrogenadas, la cual codifica, regula y/o modula la expresión genética del individuo. Así, los genes eucarióticos usualmente consisten de varios segmentos denominados segmentos codificantes de proteínas (Exones), interrumpidos por secuencias intervinientes, llamadas intrones. Adicionalmente, existen unidades funcionales tales como promotores, sitios de "splicing", enhancers (secuen cias amplificadoras) y silenciadores. También coexisten, con estas secuencias, largas cadenas de ADN sin aparente funcionalidad biológica ("Junk" DNA).
La reproducción biológica, la variación y la herencia constituyen la tríada esencial básica que nos permite comprender, bajo la presión de la selección natu ral, el origen y evolución de las especies. Ese hilo cristaloidal aperiódico (ADN) es el verdadero hilo de Ariadna que nos permite seguir, comparar, analizar y comprender las múltiples facetas del devenir de una especie. Simplemente observando su estructura (Fig. 1), el ADN ya nos sugiere la base física para su duplicación, variación (mutación) y herencia. Así tenemos que: 1) La ruptura de los puentes de hidrógeno y apertura de la doble hélice del ADN explica la duplicación y repartición de la misma información genética de una célula original a las dos células hijas descendientes; 2) El da ño físico o errores introducidos en la secuencia de letras (A, T, G, C) durante el proceso de copiado del templado, previo a la división celular, explica la variación o mutación; y 3) La información genética pasa de generación en generación con un muy alto grado de fidelidad (si bien las tasas de mutabilidad difieren de una especie a otra, éstas tienden a mantenerse en un promedio de 10-8 sustituciones/nucleótido/generación), lo que explica el fenómeno de la heredabilidad.

Estos tres preceptos básicos se conjugan para que, a partir de ello, podamos reconstruir la historia de las especies. Esa historia que conecta el origen con el devenir de los organismos. Cuando se hizo posible secuenciar genomas enteros y fragmentos parciales de ADN de especies diversas fue viable realizar: a) un análisis comparativo de secuencias (análisis por homología de se cuencia); 2) un análisis filogenético de secuencias de ADN; y 3) una inferencia de sus relaciones evolutivas. Ya que la homología de secuencia, que en primera instancia es un análisis basado en la correspondencia existente entre dos genomas dados, conlleva a una diag nosis de la divergencia evolutiva. Si, para este análisis, partimos de una secuencia ancestral común, la divergencia observada entre las secuencias descendientes será el resultado de eventos de especiación o duplicación génica producidos en dichas secuencias. Es importante aclarar a este punto la existencia de dos conceptos básicos de genes, los cuales son de una importancia filogenética remarcable, y que son el resultado de los análisis por homología: 1) Los denominados genes ortólogos y cuyo devenir evolutivo refleja aquel de la especie (p.ej. los genes que codifican para las a y b- hemoglobinas de raton y humanos); y 2) los genes paralogos, los cuales son el producto de duplicacion genica, y corren en paralelo a la historia de la especie (p.ej. a y b hemoglobina de raton). Nosotros podemos medir el tiempo de divergencia de dos especies dadas si medimos el numero de sustituciones de nucleotidos/ADN/posicion o aminoacidos/proteina/ posicion en un set de n genes o proteinas ortologas dadas (Fig. 2). Bajo esta premisa, p.ej. eubacterias y ar chaebacterias divergieron hace unos 3,5 billones de anos.
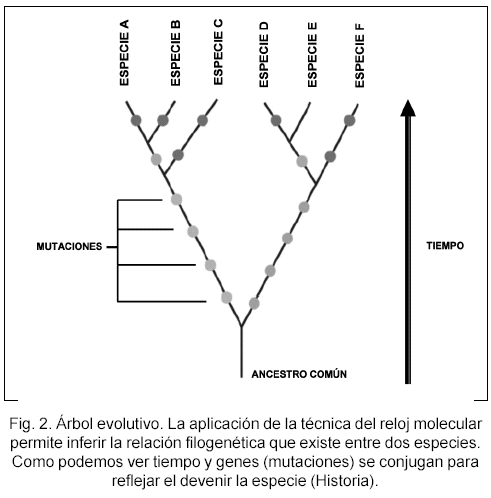
Ahora bien, los cambios acaecidos en las secuencias de ADN perteneciente a una especie dada, tienden a suceder a una tasa mas o menos predictible en el tiempo, siendo la base del denominado Reloj Molecular. Esto nos permite "caminar" por las rutas del producto del analisis filogenetico: el arbol filogenetico, el cual responde, en parte, al como evolucionan las ramas descendientes que parten de un ancestro comun dado. Asi, a partir de un grupo de especies relacionadas se puede inferir, al utilizar estas metodologias de analisis, el arbol evolutivo o filogenia de las especies seleccionadas. Simplemente, podemos medir la longitud de tiempo que le tomo a dos o mas especies divergir de su ancestro comun (Fig. 3). Por ende, el concepto de Reloj Molecular no solo nos permite explorar los acontecimientos evolutivos en el tiempo sino que tambien nos permite arrojar luz sobre los mecanismos y procesos de la evolución. La utilidad del reloj molecular queda aún más clara cuando podemos inferir, a partir de la longitud de las ramas descendientes y el ritmo de mutación, p. ej., que el último ancestro común a la actual pandemia del virus de la inmunodeficiencia humana tomó lugar en 1930 y su posible salto interespecie desde la especie Pan troglodytes troglodytes (chimpancé). La transmisión a humanos muy probablemente se produjo a través del contacto con sangre de monos infectados que habían resultado cazados y despiezados para su consumo. La expansión del grupo M VIH-1 toma lugar en la poblaciones humanas hace alrededor de unos 70 años.

BIBLIOGRAFÍA RECOMENDADA
1. Bromham, L. & Penny, D. (2003). "The Moderm Molecular Clock". NATURE REVIEWS | GENETICS. 4: 216- 224. [ Links ]
2. Mindell, D (2009). Evolution in the everyday world. Scient. Am. January 68-75. [ Links ]
