











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista Científica
versión impresa ISSN 0798-2259
Rev. Cient. (Maracaibo) v.16 n.3 Maracaibo mayo 2006
Concentraciones Sanguíneas de Calcio, Fósforo y Magnesio en Mautas Mestizas (Taurus-Indicus) Suplementadas Estratégicamente durante el periodo Peripuberal
Silvio Miranda López1, Decio González1, Nidia Rojas1 y Gloria Villalobos2
1Departamento de Producción e Industria Animal, Facultad de Ciencias Veterinarias, Universidad del Zulia. Maracaibo, Venezuela. smiranda2002@yahoo.es.
2Egresado de la Facultad de Ciencias Veterinarias, Universidad del Zulia. Maracaibo, Venezuela.
RESUMEN
Con el objetivo de examinar las concentraciones sanguíneas de calcio, fósforo y magnesio durante el período peripuberal en mautas bajo pastoreo y suplemententadas con dos estrategias alimenticias (Alimento balanceado comercial y un suplemento con Gliricidia sepium), se realizó un experimento con 24 animales mestizos doble propósito, distribuidos en forma aleatorizada en 3 grupos de 8 animales cada uno, los cuales constituyeron los tratamientos a saber: T1: consumo de gramíneas a pastoreo (CGP); T2: CGP + el consumo de alimento balanceado comercial y T3: CGP + el consumo de un suplemento alimenticio que contenía harina de hojas de Gliricidia sepium (GS). Se recolectaron muestras de suero sanguíneo, 2 semanas antes y 2 semanas después de la pubertad para determinar la concentración de calcio, fósforo y magnesio. Los datos fueron analizados a través de un análisis de varianza del sistema de análisis estadístico (SAS) empleando el procedimiento GLM. Las concentraciones sanguíneas de P y Mg observados en las semanas 2 (6,27 y 2,53 mg/dL, respectivamente), 3 (6,41 y 2,55 mg/dL, respectivamente), 4 (6,31 y 2,60 mg/dL, respectivamente) y 5 (6,39 y 2,49 mg/dL, respectivamente) superan (P < 0,01) a los mostrados en las semanas 1 (5,07 y 1,30 mg/dL, respectivamente). Particularmente el Ca sérico durante las semanas 3 (11,60 mg/dL), 4 (11,43 mg/dL) y 5 (11,72 mg/dL) fue superior (P < 0,01) que las semanas 1 (9,68 mg/dL) y 2 (10,80 mg/dL). Las mautas de T2 y T3, presentaron concentraciones sanguíneas de Ca, P y Mg, superiores (P < 0,01) a las observadas en T1. En atención a los resultados de la presente investigación, los niveles sanguíneos de Ca, P y Mg, se incrementaron una semana antes de la pubertad, siendo esto niveles mantenidos hasta la última semana del periodo peripuberal. El alimento balanceado comercial y el suplemento alimenticio con GS, fueron efectivos en aumentar las concentraciones sanguíneas de Ca, P y Mg.
Palabras clave: Mautas, pubertad, minerales, calcio, fósforo, magnesio.
Blood Levels of Calcium, Phosphorus, and Magnesium in Crossbred Heifers (Taurus-Indicus) Strategically Fed During Peripuberty Period
ABSTRACT
With the objective of examining serum concentrations of calcium, phosphorus and magnesium during the peripuberal period in grazing heifers feeding two supplemental strategies (balanced commercial food and Gliricidia sepium), An experiment was carried out with 24 dual purpose crossbred heifers, distributed in 3 randomized groups (8 animals each): T1: gramineous grazing (CGP); T2: CGP + commercial balanced food and T3: CGP + nutritional supplement that contained flour of leaves of Gliricidia sepium (GS). Serum samples were taken, 2 weeks before and 2 weeks after the puberty to determine the concentration of calcium, phosphorus and magnesium. The data was analyzed by an analysis of variance of the statistical analysis system (SAS) using the procedure GLM. The serum concentrations of P and Mg observed in the weeks 2 (6.27 and 2.53 mg/dL, respectively), 3 (6.41 and 2.55 mg/dL, respectively), 4 (6.31 and 2.60 mg/dL, respectively) and 5 (6.39 and 2.49 mg/dL, respectively) were higher (P < 0.01) to those shown in the weeks 1 (5.07 and 1.30 mg/dL, respectively). Particularly the serum Ca during the weeks 3 (11.60 mg/dL), 4 (11.43 mg/dL) and 5 (11.72 mg/dL) it was superior (P < 0.01) that the weeks 1 (9.68 mg/dL) and 2 (10.80 mg/dL). Heifers of T2 and T3, showed higher serum concentrations of Ca, P and Mg presented (P < 0.01) compared to those observed in T1. In attention to the results of the present investigation, the serum levels of Ca, P and Mg, were increased one week before the puberty, being this levels maintained until the last week of the peripuberal period. The commercial balanced food and the nutritious supplement with GS, were effective in increasing the serum concentrations of Ca, P and Mg.
Key words: Heifers, puberty, minerals, calcium, magnesium, phosphorus
Recibido: 06 / 05 / 2005. Aceptado: 05 / 03 / 2006.
INTRODUCCIÓN
La pubertad en la hembra bovina es comúnmente considerada como el período de tiempo en que inicia la función gonadal. Esta se manifiesta por la secreción de cantidades gradualmente crecientes de gonadotropinas, sobre todo la hormona folículo estimulante (FSH), y por un incremento concomitante de la capacidad de los ovarios para responder a las gonadotropinas [12]. Es así como, el inicio de la pubertad es producto de una serie de eventos complejos que son desarrollados a nivel del eje endocrino reproductivo (hipotálamo, hipófisis y ovario), lo cual permite que a determinada edad, se inicie la actividad reproductiva en la hembra bovina [29]. Smith y Akinbamijo [32], describen los cuatros factores principales que afectan el inicio de la pubertad del ganado bovino en áreas tropicales; mérito genético, medio ambiente, prácticas de manejo animal y la nutrición.
Evidencias literarias y experiencias prácticas sugieren que los factores nutricionales son considerados como los más cruciales, en términos de su incidencia directa en el fenómeno de la reproducción y por modular los efectos potenciales de los otros factores antes citados. De forma específica, la nutrición influye directamente en los eventos metabólicos que facilitan la transformación del animal joven hacia un estado adulto. Referidos eventos están acompañados con cambios en el comportamiento y apariencia corporal, los cuales han sido considerados como verdaderos indicadores externos de la pubertad. No obstante, la limitación en el suministro de nutrientes esenciales a los animales, se refleja en una alteración generalizada de las funciones corporales, cuyos efectos a edad temprana involucran retardo en la ganancia de peso e inicio de la actividad del eje hipotálamo-hipófisis-ovario [11]. Entre estos nutrientes destacan las proteínas y minerales, cuyos concentraciones en los pastos tropicales no satisfacen las cantidades requeridas por el ganado vacuno que las consume [20, 27].
Una estrategia para corregir estas deficiencias nutricionales es a través del suministro de alimentos balanceados concentrados de manufacturación comercial o suplementos alimenticios elaborados en las propias explotaciones agropecuarias. La principal limitante en el uso de alimentos balanceados comerciales es su alto costo y disponibilidad en el mercado. En referencia a los suplementos alimenticios, resultan económicos según las materias primas utilizadas. Entre las alternativas utilizadas en el trópico, para la formulación de estos suplementos destacan las hojas de leguminosas arbóreas tales como; Gliricidia sepium y Leucaena leucocephala, las cuales representan una fuente de proteína vegetal de bajo costo para la nutrición de rumiantes [Reverón y col. 1986 citado por González y col. [9, 10]. En otro aspecto, las deficiencias de minerales en el suelo y la mayoría de los forrajes de las praderas tropicales, han sido consideradas causantes de las bajas tasas de crecimiento y de los problemas reproductivos observados en los rebaños bovinos a pastoreo [3].
En el caso específico de la reproducción, la función óptima de los tejidos reproductivos de la hembra bovina, puede ser limitada por deficiencias específicas de calcio, fósforo y magnesio durante periodos críticos incluyendo la pubertad, el parto y la lactancia [14]. La participación del calcio en la pubertad es fundamentada por la participación en la biosíntesis de esteroides a nivel de ovarios, siendo los mecanismos involucrados dependientes de calcio más que del AMP cíclico (AMPc) [25]. El calcio también puede tener un importante papel en la esteroidogénesis por influenciar la utilización del colesterol en la mitocondria o por estimular la conversión de pregnolona a progesterona [38]. En este mismo orden de ideas, los factores liberatorios gonadotrópicos que estimulan las descargas de la hormona leutinizante (LH) desde las células de la adenohipófisis, involucran mecanismos dependientes del calcio. A este nivel no hay dependencia de AMPc, por lo tanto una ausencia de calcio inhibe la liberación de LH y en consecuencia es evidente un marcado atraso en la pubertad [23].
En referencia al fósforo, participa en el 95% de las reacciones de transferencia y utilización de energía influyendo directamente sobre la ganancia de peso corporal, la cual esta relacionada con el inicio de la pubertad [2, 5]. Asimismo el ovario es exigente en fósforo y en consecuencia, estados de deficiencias retrazan el inicio de la actividad ovárica y dependiendo de lo prolongado del periodo de deficiencia pudiera comprometerse de forma permanente la infuncionalidad de estos órganos reproductivos [1, 3]. La relación del magnesio y la pubertad, está circunscrita a la capacidad de este mineral en bioactivar gran parte de las enzimas que intervienen en el normal metabolismo de lípidos y carbohidratos, lo cual influye directamente sobre el balance energético del animal [19]. En líneas generales la disponibilidad de calcio, fósforo y magnesio para la pubertad es de gran importancia, ya que son requeridos en la homeorresis de los órganos reproductivos y el resto de los tejidos corporales [1, 4, 35]. Los niveles sanguíneos de calcio, fósforo y magnesio durante la pubertad en el ganado doble propósito no han sido completamente definidos [34] y en consecuencia, la posibilidad de poder inferir sobre el metabolismo y los requerimientos nutricionales de los tres minerales antes citados durante el periodo peripuberal resulta incierta [8]. Razón por la cual, el objetivo del presente ensayo fue examinar las concentraciones sanguíneas de calcio, fósforo y magnesio durante el periodo peripuberal en mautas mestizas doble propósito, bajo condiciones de pastoreo y suplementadas con dos estrategias alimenticias a saber; un alimento balanceado comercial y un suplemento alimenticio que contenía Gliricidia sepium.
MATERIALES Y MÉTODOS
Distribución y manejo del componente animal
En el experimento se utilizaron 24 mautas mestizas doble propósito (Holstein, Pardo Suizo y Zebú) pertenecientes a un rebaño comercial finca La Duquesa, localizada en el sector Río Grande, municipio Panamericano estado Táchira-Venezuela. Agroecológicamente la zona es clasificada como un bosque húmedo tropical [Ewel y col., 1968 citado por González y col. [9]; la temperatura anual oscila entre 24 y 28°C y las precipitaciones anuales entre 1800 y 2200 mm. El peso corporal y la edad promedio de las mautas al inicio del ensayo fue de 278,95 ± 32,9 kg y 22,7 ± 2,3 meses respectivamente. Los animales fueron distribuidos al azar y de forma homogénea (8 mautas por grupo) en tres grupos según el régimen alimenticio suministrado, lo cual permitió definir los tres tratamientos evaluados: T1; representado por el consumo exclusivo de gramíneas a pastoreo (CGP) principalmente pasto Tanner (Brachiaria arrecta) + 50g/animal/día de un suplemento mineral completo (50gSMC): T2; CGP + 50gSMC + 1,5 kg/animal/día de un alimento balanceado comercial (Procria, V35), según la etiqueta de fabricación fue elaborado a base de harina de germen y subproductos de maíz, harina de trigo, afrecho de arroz y trigo, levadura de cerveza, melaza, nepe de cebada, bagacillo de caña, urea, oligominerales y vitaminas: T3; CGP + 50gSMC + 2 kg/animal/día de un suplemento alimenticio elaborado en la finca, cuya composición porcentual es la siguiente: 50% de harina de maíz amarrillo molido, 30% de harina de hoja de mata ratón (Gliricidia sepium) y 20% de melaza. En la TABLA I, se indica el análisis bromatológico de pasto Tanner, alimento balanceado comercial, suplemento alimenticio y mezcla mineral comercial [9]. Las cantidades diarias suministrada de alimento balanceado comercial y el suplemento alimenticio fueron ajustados según la National Research Council [NRC, 1985 citado por González y col. [9], a objeto de permitir una ganancia de peso corporal de 500 g/día/animal. Tanto el pasto como los suplementos alimenticios fueron consumidos por un periodo no menor de tres meses antes de la aparición de la pubertad [9].

Estimación del periodo peripuberal
Para la cuantificación del periodo peripuberal, se utilizó la data aportada por González y col. [9], quienes determinaron y cuantificaron semanalmente la concentración de progesterona en el suero sanguíneo de las mautas antes citadas. La aparición de la pubertad fue establecida cuando la concentración de progesterona excedió 1 ng/mL durante dos semanas consecutivas, lo cual indica que ha ocurrido la ovulación [22]. Una vez estimada la fecha de aparición de la pubertad, se procedió a delimitar el periodo peripuberal en dos semanas antes y dos semanas después de la pubertad.
Parámetros evaluados
Los parámetros evaluados en el trabajo de investigación fueron los siguientes: Concentraciones de calcio, fósforo y magnesio en suero sanguíneo:
Se recolectaron semanalmente muestras sanguíneas por punción de la vena yugular en tubos de vidrios sin anticoagulante (Vacutainer®). Las muestras fueron centrifugadas (a 3500 rpm por un periodo de 20 minutos) una vez finalizada la faena, el suero sanguíneo extraído fue desproteinizado con ácido tricloracético al 10% (p/v) y almacenado a 5°C, hasta su posterior análisis siguiendo el método descrito por Fick y col. [6]. El calcio y magnesio fueron analizados por el método de espectrofotometría de absorción atómica de acuerdo a la técnica recomendada [26], y para la determinación del fósforo se utilizó el método colorimétrico descrito por Fiske y Subbarow [7].
Análisis estadístico
Los datos obtenidos fueron analizados a través de un diseño completamente aleatorizado y medido por un análisis de varianza de los mínimos cuadrados del paquete estadístico S.A.S [33]. Como variables dependientes se evaluaron las concertaciones de calcio, fósforo y magnesio, en el suero sanguíneo de las mautas sometidas a los diferentes tratamientos durante el periodo peripuberal; se consideró como variable independiente el efecto los suplementos alimenticios. El modelo aditivo lineal que explica el comportamiento de las variables respuestas concentración sanguínea de calcio, fósforo y magnesio fue:
Yijk = µ +Ti + Dj + TDij + Eijk.
donde:
Yijk : Concentración sanguínea de calcio, fósforo y magnesio.
µ : Media general de todas las observaciones.
Ti : Efecto del i-ésimo tratamiento (i= 1, 2 y 3) donde:
1= Pastoreo sin suplementación.
2= Pastoreo más alimento balanceado comercial.
3= Pastoreo más suplemento alimenticio.
Dj : Efecto de la j-esima (j = 1, 2, 3, 4 y 5) semana.
Tdij : Interacción del i-esimo tratamiento y la j-esima semana.
Eijk : Efecto de los factores no controlados en el experimento sobre las unidades
experimentales en el k-ésimo período, j-ésimo unidad experimental y que recibió
el i-ésimo tratamiento.
Para comparar la media de las variables respuestas, donde fueron detectadas diferencias significativas entre tratamientos, se utilizó la prueba de t por medio de la aplicación LSMEANS de procedimiento GLM del S.A.S [33].
RESULTADOS Y DISCUSIÓN
Concentraciones sanguíneas de calcio, fósforo y magnesio durante el periodo peripuberal
En la TABLA II, se observa el promedio de las concentraciones sanguíneas de calcio, fósforo y magnesio en el rebaño experimental durante las cinco semanas del periodo peripuberal.
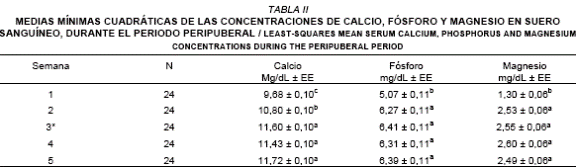
a, b, c: Valores con letras distintas dentro de la misma columna, presentan diferencias estadísticas (P < 0,01). EE: Error Estándar. * Durante esta
semana los animales fueron púberes.
Los niveles de calcio sanguíneo en las semanas; 1, 2, 3, 4 y 5 fueron: 9,68; 10,80; 11,60; 11,43 y 11,72 mg/dL respectivamente. Según el análisis estadístico (P < 0,01) los valores observados en las semanas 3, 4 y 5, superan a los mostrados en las semanas 1 y 2. Esta última semana, presentó concentraciones de calcio sanguíneo superiores estadísticamente (P < 0,01) a la semana 1. Los valores normales de calcio en suero sanguíneo varían de 9 a 13 mg de Ca/dL para bovinos jóvenes [36, 37]. En consideración a este rango y al nivel crítico sugerido por McDowell y col. [18] de 8 mg de Ca/dL, podemos indicar que todas las mautas pertenecientes al rebaño experimental presentaron valores normales durante el período peripuberal. El incremento del calcio sanguíneo observado en el presente ensayo al inicio de la pubertad, pudiera estar relacionado con la participación de varios factores metabólicos a considerar; el calcio favorece la secreción de factores liberatorios gonadotrópicos con la consecuente liberación de las hormonas peptídicas FSH y LH [23], e interviene en la síntesis de progesterona [38].
Las concentraciones sanguíneas de fósforo en las semanas 1, 2, 3, 4 y 5 fueron; 5,07; 6,27; 6,41; 6,31 y 6,39 mg/dL respectivamente. Según el análisis, diferencias significativas (P < 0,01) fueron detectadas a favor de las semanas 2, 3, 4 y 5 vs.1, siendo esta última semana la que refirió los más bajos valores de fósforo sanguíneo. Sin embargo, en atención al nivel crítico de deficiencia para el fósforo sugerido por McDowell y col. [18] de 4,5 mg P/dL, todas las mautas durante el período peripuberal no presentaron deficiencias de fósforo. Los resultados observados en el presente trabajo, permiten indicar que una semana antes de la pubertad de las mautas, los niveles sanguíneos de fósforo se incrementaron para luego mantenerse durante el resto del período peripuberal. Este aumento de fósforo sanguíneo tiene gran significancia para el acondicionamiento metabólico que es requerido por el animal al inicio de la pubertad. Desde el punto de vista hormonal, gonadotropinas hipofisiarias relacionadas con la pubertad [22] demandan para su síntesis (conformación de la cadena peptídica) gran cantidad de energía química, constituida principalmente por adenosín trifosfato [14]. El adenosín trifosfato (ATP), está constituido por adenina (base nitrogenada), ribosa y una unidad de trifosfato (rica en fósforo), permitiendo este último un alto potencial de transferencia de grupos [17], necesarios para la conformación de los enlaces peptídico de las hormonas antes citadas [15, 25].
Los valores sanguíneos de magnesio en las semanas 1, 2, 3, 4 y 5 presentaron las siguientes concentraciones; 1,30; 2,53; 2,55; 2,60 y 2,49 mg/dL respectivamente. Se observaron diferencias estadísticas a favor de las semanas 2, 3, 4 y 5 vs.1, donde esta última semana refirió la más baja concentración sanguínea de magnesio. No se evidenciaron diferencias significativas entre las semanas 2, 3, 4 y 5. Las concentraciones sanguíneas normales de magnesio varían desde 1,7 a 3,3 mg/dL [16], estableciendo como nivel crítico de deficiencia niveles inferiores a los 1,0 mg/dL [18]. Basándose en este criterio y fundamentado en que los niveles representan un buen indicador del contenido corporal [14], no se detectaron deficiencia de magnesio en el rebaño experimental. Sin embargo, es importante destacar que en la semana 1, la concentración sanguínea de magnesio fue inferior a los valores normales, pero superior al nivel crítico de deficiencia sugerido por McDowell y col. [18]. Los valores sanguíneos de magnesio en la segunda semana, fueron el doble de lo observado en la primera semana, este evento pudiera estar relacionado con la capacidad del magnesio, en bioactivar de forma específica la adenilciclasa o la fosfolipasa C. La adenilciclasa es formada tras la conjugación de la LH o FSH con su receptor a nivel ovárico, lo cual origina un segundo mensajero el AMP cíclico o la fosfolipasa C [8].
Concentraciones sanguíneas de calcio, fósforo y magnesio afectadas por el tipo de dieta, durante el periodo peripuberal
El efecto de los tratamientos en estudio sobre las concentraciones sanguíneas de calcio, fósforo y magnesio durante el periodo peripuberal es presentado en la TABLA III.

Los valores promedios de calcio sanguíneo en los tratamientos 1, 2 y 3 fueron; 10,13; 11,26; 11;64 mg/dL respectivamente. Según el análisis estadístico (P < 0,01) las concentraciones observadas en los tratamientos 2 y 3, superan las mostradas en el tratamiento 1. Estas diferencias observadas en el presente trabajo, a favor de las mautas de los tratamientos 2 y 3, pueden ser atribuidas al consumo estratégico del alimento balanceado comercial y del suplemento alimenticio con Gliricidia sepium, alimentos cuyo contenido de calcio es ilustrado en la TABLA I, suministraron adicionalmente a lo aportado por la dieta basal (suplemento mineral completo + forraje) 7,2 y 10,4 g de Ca/animal/d en los tratamientos 2 y 3. Este efecto aditivo del calcio dietético sobre los valores sanguíneos del mismo, concuerdan con reportes anteriores en bovinos jóvenes [5, 21].
Los promedios de las concentraciones de fósforo sanguíneo para los tratamientos 1, 2 y 3 fueron; 5,08; 6,22 y 6,40 mg/dL respectivamente. El tratamiento 1 refirió estadísticamente (P < 0,01), los más bajos valores de fósforo sanguíneo en comparación con los tratamientos 2 y 3. Estos dos últimos tratamientos no mostraron diferencias significativas. En el presente ensayo, las concentraciones sanguíneas de fósforo en las mautas de los tratamientos 2 y 3, fueron incrementadas por un aumento sustancial del fósforo dietético (7,8 y 5,0 g de fósforo, para los tratamientos 2 y 3, respectivamente), tras el consumo del alimento balanceado y suplemento alimenticio con Gliricidia sepium. Al respecto, Espinoza y col. [5], y Horst [13] establecen en rumiantes, que la absorción del fósforo está relacionada con la cantidad de este mineral presente en la dieta y también se relaciona directamente con las concentraciones del fósforo sanguíneo.
Los niveles de magnesio en el suero sanguíneo fueron altos (P < 0,01) en las mautas pertenecientes a los tratamientos 2 y 3 con 2,24 y 2,42 mg de mg.dL respectivamente vs. 1,10 mg de mg.dL del tratamiento 1. Las concentraciones observadas en el tratamiento 1, son inferiores a los valores sanguíneos normales citados por Littledike y Goff, [16], pero superiores al nivel crítico de deficiencia [18]. Los tratamientos 2 y 3, no presentaron diferencias significativas. El magnesio presente en el alimento balanceado comercial y suplemento alimenticio con Gliricidia sepium, proporciona 4,5 y 4,0 g/animal/día para los tratamientos 2 y 3 respectivamente. Este magnesio dietético adicional a lo consumido por las mautas del tratamiento 1, presumiblemente incrementó las concentraciones sanguíneas de magnesio en los tratamientos antes citados, lo cual concuerda con las observaciones de Neathery y col. [24], quienes indican que los valores de magnesio en suero sanguíneo son afectados por la cantidad de magnesio consumido, absorbido o movilizado desde el tejido óseo.
Concentraciones sanguíneas de calcio afectadas por el tipo de dieta y la semana, durante el periodo peripuberal
Las concentraciones de calcio en suero sanguíneo en cada una de las cinco semanas que constituyó el periodo peripuberal, son presentadas en la TABLA IV.

En la misma se observa que durante las cinco semanas del período peripuberal las mautas que consumieron los suplementos de los tratamiento 2 y 3 (alimento comercial y suplemento alimenticio con Gliricidia sepium respectivamente), presentaron estadísticamente las mayores (P < 0,01) concentraciones sanguíneas de calcio vs. el tratamiento 1, cuyas mautas no consumieron un suplemento alimenticio adicional a su dieta basal (pastoreo). No se detectaron diferencias significativas entre los tratamientos 2 y 3. En este mismo orden de ideas, al analizar el efecto de la semana sobre las concentraciones sanguíneas de calcio dentro de los tratamientos evaluados, se evidenció diferencias significativas (P < 0,01), específicamente en la semana 1 del tratamiento 1, el cual reveló los más bajos niveles de calcio sanguíneo (8,63 mg de Ca/dL) para mencionado tratamiento. No obstante, las mautas del tratamiento 2 en las semanas 1 y 2 (10,32 y 10,98 mg de Ca/dL respectivamente) mostraron niveles inferiores (P < 0,01), al resto de las semanas dentro del mismo tratamiento. Similarmente la semana 1 del tratamiento 3, refirió los valores más bajos (P < 0,01) de calcio sanguíneo (10,13 mg de Ca/dL) que el resto de las semanas de periodo peripuberal. Sin embargo, en las semanas 3 y 4 las concentraciones sanguíneas de calcio (12,42 y 12,39 mg Ca/dL respectivamente) fueron significativamente superiores (P < 0,01) a los observados en las semanas 2 y 5 (11,40 y 11,92 mg de Ca/dL respectivamente). Escasa es la literatura que revela los niveles sanguíneos de calcio durante el periodo peripuberal, no obstante existen reportes en otras fases reproductiva del ganado bovino que facilitan el entendimiento de las pautas metabólicas del calcio en el fenómeno de la reproducción. En este sentido, Small y col. [30] observaron un ligero aumento en la concentración de calcio sanguíneo al día 21 del ciclo estrual, en aquellos animales que posteriormente concebían. Littledike y Goff [16], indican una elevación del calcio sanguíneo 2 a 3 días antes del parto, para luego descender bruscamente en niveles inferiores a los considerados normales [31].
Concentraciones sanguíneas de fósforo afectadas por el tipo de dieta y la semana, durante el periodo peripuberal
La TABLA V, muestra los niveles sanguíneos de fósforo en las mautas sometidas a los tres tratamientos evaluados, durante las cinco semanas del periodo peripuberal.

En la primera semana del periodo peripuberal, las mautas del tratamiento 2 presentaron los valores más altos (P < 0,01) de fósforo sanguíneo (5,40 mg de P/dL) vs. los tratamientos 1 y 3 (4,78 y 5,04 mg de P/dL, respectivamente). En las semanas 2 y 3, se detectaron diferencias significativas (P < 0,01) a favor de los tratamientos 2 y 3 (6,41; 7,12 y 6,67; 7,17 mg de P/dL respectivamente) en comparación con el tratamiento 1 (5,44 y 6,44 mg de P/dL).
La cuarta semana no reveló diferencias estadísticas, las cuales si fueron evidenciadas en la quinta semana, donde el tratamiento 2 refirió las concentraciones más bajas de fósforo sanguíneo (5,85 mg de P/dL) vs. los tratamientos 1 y 3 (6,11 y 6,83 mg de P/dL, respectivamente). Con respecto al efecto de las semanas sobre las concentraciones sanguíneas de fósforo dentro de los tratamientos, se observó que las mautas presentes en los tres tratamientos estudiados, incrementaron significativamente (P < 0,05) los niveles sanguíneos de fósforo, a partir de la semana 2 del periodo peripuberal. Estos niveles se mantuvieron hasta la quinta semana, excepto para el tratamiento 2, donde en la semana 5 fue evidente una reducción del fósforo sanguíneo. Resulta importante destacar que los tratamientos 2 y 3, presentaron en la tercera semana (inicio la pubertad) las más altas concentraciones de fósforo en suero sanguíneo. En la semana 1, las mautas de los tres tratamientos estudiados mostraron los valores más bajos de fósforo sanguíneo. Poca literatura describe los niveles de fósforo sanguíneo durante el periodo peripuberal, no obstante existe reportes en otras etapas reproductiva del ganado bovino que permiten comprender la participación del fósforo en la reproducción. Al respecto, Small y col. [30] observaron un incremento significativo en la concentración del fósforo sanguíneo al día 21 del ciclo estrual, en vacas adultas y novillas. Estos mismos autores sugieren que las concentraciones de fósforo sanguíneo en ambas, son indicativas de una hiperfosfatemia, sin embargo los niveles de fósforo en novillas fueron mayores que las vacas adultas. Littledike y Goff [16], indican una elevación del fósforo sanguíneo 2 a 3 días antes del parto, para luego descender bruscamente en niveles por inferiores a los considerados normales [31].
Concentraciones sanguíneas de magnesio afectadas por el tipo de dieta y la semana, durante el periodo peripuberal
Las concentraciones de magnesio en suero sanguíneo observadas en las mautas de los tres tratamientos evaluados durante las cincos semana del periodo peripuberal, son mostradas en la TABLA VI.

En la primera y quinta semana del periodo peripuberal, los tratamientos estudiados no muestran diferencias significativamente en los niveles de magnesio sanguíneos. Sin embargo, en las semanas 2, 3 y 4, se detectaron diferencias estadísticas (P < 0,01) a favor de los tratamientos 2 y 3 (2,48; 2,53; 2,86 y 2,84; 2,67; 2,99 mg de Mg/dL, respectivamente), siendo el tratamiento 1 el cual refirió los más bajos valores de magnesio sanguíneo (1,96; 2,36 y 2,26 mg de Mg/dL, respectivamente). No obstante, en la semana 3 los tratamientos 1 y 2, no presentaron diferencias estadísticas. En relación al efecto de las semanas sobre los valores promedios de magnesio en suero sanguíneo dentro de los tratamientos, se observó un incremento significativo (P < 0,01) de las concentraciones sanguíneas de magnesio a partir de la segunda semana en los tres tratamientos estudiados, concentraciones éstas que se mantuvieron hasta la quinta semana. Los resultados anteriores indican que las mautas pertenecientes a los tres tratamientos, aumentaron los valores de magnesio sanguíneo una semana antes de la pubertad, para posteriormente mantenerse una vez iniciada la pubertad. Al igual que el calcio y fósforo, hay pocas evidencias literarias que muestre los niveles sanguíneos de magnesio en el periodo peripuberal. Sin embargo, algunos reportes en novillas y vacas adultas durante el periodo estrual, refieren una disminución de magnesio sanguíneo aun en animales suplementados [30]. Estos hallazgos difieren de los resultados obtenidos en el presente ensayo, donde es evidente un incremento del magnesio sanguíneo. Otras fluctuaciones de este elemento mineral son detectadas en vacas adultas antes y en el parto. Existen evidencias que los niveles sanguíneos de magnesio 7 días antes del parto se encuentra por debajo de los valores normales pero superando los 0,85 mmo L-1 [28]. En el parto, la concentración sanguínea de magnesio incrementó, tanto en vacas adultas como en novillas [31].
CONCLUSIONES
Las concentraciones de calcio, fósforo y magnesio sanguíneo, permanecieron dentro de los valores considerados normales para la especie. Sin embargo, una semana antes de la pubertad, las mautas experimentaron un aumento en las concentraciones sanguíneas de los tres elementos minerales antes citados, siendo estos valores sanguíneos mantenidos hasta el final del periodo peripuberal.
Las dos estrategias alimenticias evaluadas (alimento balanceado comercial y un suplemento alimenticio que contenía Gliricidia sepium), fueron efectivas en incrementar los niveles sanguíneo de calcio, fósforo y magnesio.
RECOMENDACIONES
El aumento en las concentraciones sanguíneas de calcio, fósforo y magnesio una semana antes de la pubertad, sugiere cambios en los programas alimenticios de las mautas destinadas al reemplazo del rebaño de producción. Estos cambios involucran la utilización de suplementos alimenticios que contengan fuentes proteicas y de minerales, de bajo costo y fácil adquisición, como por ejemplo Gliricidia sepium.
AGRADECIMIENTO
Los autores agradecen al Consejo de Desarrollo Científico y Humanístico (CONDES) de la Universidad del Zulia y laboratorio de Nutrición Animal de la Cátedra Nutrición y Alimentación Animal, Facultad de Ciencias Veterinarias, por el soporte logístico y financiero otorgado para el desarrollo de esta investigación.
REFERENCIAS BIBLIOGRÁFICAS
1. BUTLER, W.R. Nutritional interactions with reproductive performance in dairy cattle. Anim. Reprod. Sci. 60: 449-457. 2000. [ Links ]
2. CALL, J.W.; BUTCHER, J. E.; BLAKE, J.T.; SMART, R.A.; SHUPE, J.L. Phosphorus influence on growth and reproduction of beef cattle. J. Anim. Sci. 47: 216-225. 1978. [ Links ]
3. CIPAGAUTA, M.; LAREDO, M.; PALADINES, O. Suplementación de ganado bovino con tres fuentes de fósforo, en los llanos orientales de Colombia. Rev. ICA. 27: 333-345. 1992. [ Links ]
4. DEVENPORT, G.M.; BOLING, J.A.; GAY, N. Bioavailability of magnesium in beef cattle fed magnesium oxide or magnesium hydroxide. J. Anim. Sci. 68: 3765-3772. 1990. [ Links ]
5. ESPINOZA, J.E.; MCDOWELL, L.R.; WILKINSON, N.S.; CONRAD, J.H.; MARTIN, F.G.; WILLIAMS, S. N. Effect of dietary phosphorus level on performance and mineral status of grazing cattle in a warm climate region of central Florida. Livest. Res. for Rural Develop. htt://www. cipav.org.co/lrrd/. 3(1). 1991. [ Links ]
6. FICK, K.R.; MILLER, S. M.; FUNK, J.D.; McDOWELL, L.R.; HOUSER, R.H. Methods of mineral analysis fro plant and animal tissues. University of Florida. Animal Science Department. Gainesville, Florida, USA. 502 pp. 1979. [ Links ]
7. FISKE, C.H.; SUBBAROW, Y. The colormetric determination of phosphorus. J. Biol. Chem. 66: 375-384. 1925. [ Links ]
8. GOFF, J.P. Pathophysiology of calcium and phosphorus disorders. Veterinary Clinics of North America: Food Anim. Pract. 16(2): 319-337. 2000. [ Links ]
9. GONZALEZ, D.; QUINTERO, A.; PALOMARES, R.; ROJAS, N.; ARAUJO, O.; SOTO, G. Uso de la Gliricidia sepium en la suplementación de mautas mestizas y su efecto sobre el crecimiento y la pubertad. Rev. Científ. FCV-LUZ XII (1): 45-52. 2003. [ Links ]
10. GONZALEZ, I.; FARIA, J.; MORRILLO, D.; MAVAREZ, O.; NOGUERA, N.; FUENMAYOR, E. Efecto de frecuencias de riego y corte sobre el rendimiento de materia seca en Leucaena leucocephala (Lam) De wit. Rev. Fac. Agron. (LUZ). 20: 263-272. 2003. [ Links ]
11. GRINGS, E.E.; STAIGMILLER, R. B.; SHORT, R.E.; BELLOWS, R.A.; MACNEIL, M, D. Effects of stair-step nutrition and trace mineral supplementation on attainment of puberty in beef heifers of three sire breeds. J. Anim. Sci. 77:810-815. 1999. [ Links ]
12. HANSEL, W.; McENTEE, K. Procesos reproductores en la hembra. En: Dukes, H. y Swenson, M. (Ed.), Fisiología de los animales domésticos. 4ta Ed. Ediciones Aguilar: 1593-1648 pp. 1981. [ Links ]
13. HORST, R.L. Regulation of calcium and phosphorus homeostasis in the dairy cow. J. Dairy Sci. 69: 604-616. 1986. [ Links ]
14. HURLEY, L.W.; DOANE, W. L. Recent developments in the roles of vitamins and minerals in reproduction. J. Dairy Sci. 72: 784-804. 1989. [ Links ]
15. LEHNINGER, A. Bioquímica; Las bases moleculares de la estructura y función celular. 2ª Ed. Ediciones Omega, S.A. 1117 pp. 1985. [ Links ]
16. LITTLEDIKE, E.T.; GOFF, J. Interactions of calcium, phosphorus, magnesium and vitamin D that influence their status in domestic meat animals. J. Anim. Sci. 65: 1727-1743.1986. [ Links ]
17. MAYNARD, L.A.; LOOSLI, J. K.; HINTZ, H.F.; WARNER, R.G. Nutrición Animal. 7ma Ed. McGraw-Hill. 640 pp. 1981. [ Links ]
18. MCDOWELL, L.R.; CONRAD, J.H.; ELLIS, G.L. Rumiant mineral deficiencies: Radioisotopic and other techniques of delection. In: IESA-SR. 151-169 pp. 1985. [ Links ]
19. MCDOWELL, L.R.; VELÁSQUEZ, J.; VALLE, G. Minerales para rumiantes en pastoreo en regiones tropicales. Departamento de Zootecnia. Centro de agricultura tropical. Universidad de Florida, Gainesville. 3ra Ed. 84 pp. 1997. [ Links ]
20. MINSON, D.J. Forage in ruminant nutrition. Academic Press Inc. 483 pp. 1990. [ Links ]
21. MIRANDA, S.; LOPEZ, R.; DEAN, D.; ROMAN, R.; VILLALOBOS, G.; NAVAS, Y.; Respuesta al crecimiento y concentraciones sanguíneas de calcio, fósforo, magnesio y potasio en novillos mestizos (Taurus-Indicus) suplementados con una mezcla mineral completa. Rev. Científ. FCV-LUZ XIII (3): 221-228. 2003. [ Links ]
22. MORAN, C.; QUIRKE, J.F.; ROCHE, J. F. Puberty in heifers: a Review. Animal Reproduction Science. 167-182 pp. 1989. [ Links ]
23. NAOR, Z.; CATT, K. J. Mechanism of action of gonadotropin-releasing hormone. J. Biol. Chem. 256: 2226-2237. 1981. [ Links ]
24. NEATHERY, M.W.; CROWE, N.A.; MILLER, W.J.; CROWE, C.T.; VARNADOE, J. L.; BLACKMON, D. M. Effects of dietary aluminum and phosphorus on magnesium metabolism in dairy calves. J. Anim. Sci. 68:1133-1138. 1990. [ Links ]
25. NORMAN, A.W.; LITWACK, G. Steroid hormones: Chemistry, biosynthesis, and metabolism. In: Hormones. 2nd Ed. Academic Press. 58-75 pp. 1997. [ Links ]
26. PERKIN-ELMER. Analytical methods for atomic absorption espectrophotometry. Norwalk, Conneticut. U.S.A. 82 pp. 1982. [ Links ]
27. POPPI, D.; McLENNAN, S. Protein and energy utilization by rumiants at pasture. J. Anim. Sci. 73: 278-290. 1995. [ Links ]
28. SANSOM, B. F.; MANSTON, R.; VAGG, M. J. Magesium and milk fever. Vet. Rec. 112: 447-449. 1983. [ Links ]
29. SCHILLO, K.K.; HALL, J.B.; HILEMAN, S. M. Effects of nutrition and season on the onset of puberty in the beef heifer. J. Anim. Sci. 70:3994-4005. 1992. [ Links ]
30. SMALL, J.A.; CHARMLEY, E.; RODD, A.V.; FREDEEN, A.H. Serum mineral concentrations in relation to estrus and conception in beef heifers and fed conserved forage. Can. J. Anim. Sci. 77: 55-62. 1997. [ Links ]
31. SMALL, J.A. Serum mineral concentrations in relation to parturition in beef heirfs and cows fed conserved forage. Can. J. Anim. Sci. 77: 63-68. 1997. [ Links ]
32. SMITH, O.B.; AKINBAMIJO, O.O. Micronutrients and reproduction in farm animals. Anim Reprod Sci. 60-61:549-60. 2000 [ Links ]
33. STATISTICAL ANALISYS SYSTEM INSTITUTE (S.A.S). Users Guide Basics. Cary, N.C. Version 8,1. 1999. [ Links ]
34. SUTTLE, N. F. Predicting the risk of mineral deficiencies in grazing animals. South Afric. J. of Anim. Sci. 18(1):15-22. 1988. [ Links ]
35. SWANSON, L.V. Discussion-Interactions of nutrition and reproduction. J. Dairy Sci. 72: 805-814. 1989. [ Links ]
36. TIFFANY, M.E.; MCDOWELL, L.R.; O´CONNOR, G.A.; MARTIN, F. G.; WILKINSON, N.S.; PERCIVAL, S.S.; RABIANSKY, P.A. Effects of residual and reapplied biosolids on performance and mineral status of grazing beef steers. J. Anim. Sci. 80:260-269. 2002. [ Links ]
37. UNDERWOOD, E.J. The mineral nutrition of livestock Second. Commonwealth Agriculture Bureaw. Edition London. 210 pp. 1981. [ Links ]
38. VELDHUIS, J.D.; KLASE, P. A. Mechanisms by which calcium ions regulate the steroidogenic actions of luteinizing hormone in isolated ovarian cells in vitro. Endocrinol. 111: 1-9. 1982. [ Links ]
