











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista Científica
versión impresa ISSN 0798-2259
Rev. cient. (Maracaibo) v.20 n.4 Maracaibo jul. 2010
RELATIONSHIP OF MILD FATTY LIVER, b–CAROTENE, VITAMINS A AND E STATUS OF PERIPARTURIENT HOLSTEIN COWS
Oswaldo Rosendo 1*, Lee R. McDowell 2, Charles Staples 2, Jean K. Shearer 3,
Nancy S. Wilkinson 2 and William M. Seymour 4
1* Decanato de Ciencias Veterinarias. Universidad Centroccidental Lisandro Alvarado, Tarabana, Venezuela.
2 Departmentof Animal Sciences, University of Florida, Gainesville 32611, Fl. USA.
3 College of Veterinary Medicine, University of Florida, Gainesville 32611, Fl. USA. 4 DSM Vitamins Inc., Parsippany, NJ 07054. * orosendo@ucla.edu.ve. Phone: 58251/2592687.
Abstract
The objectives of this study were to determine the association between mild fatty liver and b-carotene (BC), retinol (ROL), and a-tocopherol (AT) status. Multiparous Holstein cows were defined as having mild fatty liver using a threshold of 2% liver triacylglycerol (TAG, wet basis) at day (d) 2 after calving. Cows with mild fatty liver decreased dry matter intake (DMI) as a % of body weight (BW) from calving to 5th week (wk) of lactation (3.47 vs. 3.99, P = 0.02) and increased plasma nonesterified fatty acids (NEFA) concentration (345.0 vs. 266.0 µeq/dL, P = 0.10) as compared to normal cows. Fatty liver tended to lower plasma ROL (0.20 vs. 0.23 µg/mL, P = 0.10), but had no effect on plasma BC, plasma AT, liver BC, and liver ROL (P > 0.12) concentrations. However, depression in liver BC at calving was very small (8.0 vs. 68.0%) for fatty liver than for normal cows. Liver AT concentrations were greater (5.5 vs. 4.0 µg/g, P = 0.06) for cows with more than 2% liver TAG concentration. Concentration of plasma NEFA was positively correlated with plasma BC concentrations (r = 0.18, P < 0.05) and negatively correlated with concentrations of plasma ROL (r = -0.36, P < 0.0001) or AT (r = -0.35, P < 0.0001). A positive direct relationship between DMI and plasma vitamins status was found to exist only for ROL and AT, which suggests that stores of ROL or AT are less labile than stores for BC. At different times from calving, liver BC concentrations were positively correlated with liver TAG concentrations in a consistent manner but, correlations among liver ROL or liver AT concentrations and liver TAG concentrations were nonsignificants. The results suggest that the regulatory role of the liver in maintaining retinol concentrations in plasma may be compromise in cows with fatty liver.
Key words: Fatty liver, dairy cow, vitamins.
Relación del hígado graso leve y el estatus de b–caroteno, vitaminas A y E en vacas Holstein durante el periparto
RESUMEN
Este estudio fue realizado con los objetivos de determinar la asociación entre hígado graso leve (HGL) y estatus de b-caroteno (BC), retinol (ROL) y a-tocoferol (AT). Para definir HGL en vacas Holstein multíparas se empleó como valor crítico el 2% de triacilgliceroles (TAG, base fresca) en el hígado al segundo día después del parto. El consumo de materia seca (CMS), como porcentaje del peso vivo, disminuyó (3,47 vs. 3,99; P = 0,02) mientras que la concentración de ácidos grasos no-esterificados (AGNE) incrementó (345,0 vs. 266,0 µeq/dL; P = 0,10), desde el parto hasta la 5ta semana de lactancia en las vacas con HGL en comparación con las vacas control. El hígado graso tuvo la tendencia a disminuir el ROL plasmático (0,20 vs. 0,23 µg/mL, P = 0,10), pero no afectó las concentraciones de BC y AT en plasma, ni las de BC y ROL hepático (P > 0,12). Sin embargo, en el parto, la caída del BC hepático fue muy pequeña (8,0 vs. 68,0%) en las vacas con HGL. Las concentraciones de AT hepático fueron mayores (5,5 vs. 4,0 µg/g, P = 0,06) en las vacas con más de 2% de TAG hepático. La concentración plasmática de AGNE tuvo una correlación positiva con las concentraciones plasmáticas de BC (r = 0,18; P < 0,05) pero negativa con las concentraciones de ROL (r = -0,36: P < 0,0001) o AT (r = -0,35; P < 0,0001) plasmático. Solo el estatus plasmático de ROL y AT evidenciaron una relación directa positiva con el CMS, lo que sugiere que los depósitos de ROL o AT son menos lábiles que los de BC. En días diferentes del postparto, las concentraciones hepáticas de BC fueron correlacionadas positivamente con las concentraciones hepáticas de TAG de una manera consistente, pero las correlaciones entre las concentraciones de ROL o AT hepático vs. las de TAG hepático no fueron significativas. Los resultados sugieren que la función reguladora del hígado para mantener las concentraciones plasmáticas de retinol puede estar comprometida en las vacas que sufren hígado graso.
Palabras clave: Hígado graso, vaca lechera, vitaminas.
Recibido: 29 / 04 / 2009. Aceptado: 01 / 03 / 2010.
Introduction
During the transition from the nonlactating to the lactating state in the dairy cow (Bos taurus indicus), changes in b-carotene (BC) and vitamins A and E status occur but causes are still unclear. Fatty liver can develop during the transition period, and changes on status of these vitamins may be closely related to changes in liver triacylglycerol (TAG) concentration.
A negative association between decreased plasma BC and body fat mobilization at the time of parturition has been suggested [5, 10]. However, based on recent estimates, an increment on plasma nonesterified fatty acids (NEFA) during the periparturient period would be associate to a negligible reduction (an increase of 100 µeq/dL in NEFA would be expected to reduce BC by only 0.02 µg/mL) of plasma BC [14]. Cows with fat cow syndrome or severe fatty liver had lower plasma retinol (ROL) and plasma a-tocopherol (AT) concentrations than periparturient healthy cows [11]. In contrast, Shaw [27] did not observe a relationship between ketosis, a disorder led by fatty liver, and low plasma vitamin A concentrations. In cows with mild fatty liver (> 2% liver TAG concentration), plasma AT concentrations decreased whereas liver AT concentrations tended to increase [19].
Cows that develop fatty liver during the transition period could have a higher risk for retained placenta, metritis, and mastitis [1], conditions all associated with a lower AT, ROL or BC status [13, 14, 16]. However, the dynamics of all BC, ROL, and AT, in both plasma and liver, in relation to fatty liver development have not been elucidated for periparturient dairy cows. Therefore, the objectives of this study were to evaluate the relationships between mild fatty liver, plasma and liver BC, ROL, and AT concentrations.
Materials and Methods
Management of cows
This study was conducted at the University of Florida, United State of America (USA), with the approval of the Animal Care and Use Committee. Thirty-four multiparous and pregnant Holstein cows were monitored from 16 days (d) prepartum (d -16) to 35 d postpartum (d 35). Mean 305-d milk production in the previous lactation was 10,533 ± 1,489 kg, and mean body weight was 648 ± 76 kg at the start of the experiment, 16 d before expected calving day. During the close-up dry period, cows were housed in shaded pens, fed individually, and had free access to tap water. A total mixed ration (DM basis) of corn (Zea maiz) silage (38.4%), bermudagrass (Costal bermudagrass) hay (13.6%), citrus (Citrus spp.) pulp (3.8%), and concentrate (44.2%) that provided 1.53 Mcal/kg of net energy of lactation (NEL), 13.4% of crude protein (CP), and 36.1% of neutral detergent fiber (NDF) (dry matter basis) was offered during the close-up dry period. These cows were supplemented (based on feed chemical composition of mineral and vitamin mix) with 80,000 IU of vitamin A/d, 10,000 IU of vitamin D/d, and 350 IU of vitamin E/d. After calving cows were housed in a free-stall barn and fed a diet based (DM basis) on corn silage (26.6%), alfalfa (Medicago sativa) hay (10.0%), citrus pulp (9.5%), cottonseed (Gossypium herbaceum) hulls (5.1%), and concentrate (48.8%). This diet consisted of 1.63 Mcal/kg of NEL, 17.3% of CP, and 34.3% of NDF (DM basis). Lactating cows were supplemented (based on feed chemical composition of mineral and vitamin mix) with 60,000 IU of vitamin A/d, 18,000 IU of vitamin D/d, and 350 IU of vitamin E/d. Dry matter content of corn silage was determined weekly at 55°C to adjust as-fed ratios of forage and concentrate. Feed offered and refused was recorded daily from d -16 prepartum to d 35 postpartum. The cows were milked three times a day.
Sampling and chemical analyses
Body weight was determined on approximately d 20 (pretreatment) before expected calving day, and on the same day each week throughout the trial after the morning milking. Blood samples were collected via the coccygeal artereo-vein at 5 h after the morning feeding on week -2, -1, 0, 1, 2, 3, 4, 5 relative to week of calving using evacuated tubes containing sodium heparin (Vacutainer®; Becton Dickinson, Franklin Lakes, NJ, USA). Tubes were immediately placed on ice and centrifuged (Superspeed Refrigerated Centrifuge, RC-5, Sorvall, USA) within 4 h, at 4°C for 30 min at 3,000 x g. Plasma was harvested and stored at -20°C until analyzed for nonesterified fatty acids (NEFA) using the NEFA-C kit (Wako Chemicals USA Inc., Richmond, VA, USA) with the modifications of Johnson and Peters [12]. Plasma glucose was analyzed with an automated analyzer (Bran + Luebbe, Model II, Bran + Luebbe Analyzing Technologies, Elmsford, NY, USA), following a colorimetric procedure [4]. Liver biopsy samples were collected with trocar and cannula [23] at the 11th intercostals space at d 16 (10 to 30 d) before calving date, and on d 2 (1 to 4), 16 (14 to 18 d), and 30 (27 to 34 d) postpartum on the same cows. Upon collection the liver tissue was rinsed with saline to removed excess blood and immediately placed into liquid nitrogen. After freezing, samples were placed on dry ice for up to 2 h. Samples were stored at -70°C until analyzed for triacylglycerol [23]. A cow was classified as having fatty liver when liver TAG concentration was more than 2% (wet weight) at d 2 after calving. None of the cows had severe fatty liver (more than 10% liver TAG).
Concentrations of plasma and liver BC, ROL, and AT were determined by High Performance Liquid Cromatografy (HPLC) after double extraction with petroleum ether. The procedures have been described previously for measuring AT [21], plasma BC and ROL [2] and liver BC and ROL [28].
Statistical analysis
Plasma, liver, and performance data were analyzed using PROC MIXED of SAS [26] according to the following model:
Yijk = µ + Ti + Cji + Dk + TDik + eijk
Where:
Yijk is the dependent variable,
µ is the overall mean of the population,
Ti is the effect of Liver Condition i (normal or fatty),
Cji is the effect of Cow j on Liver Condition i,
Dk is the effect of Time Period k of sampling relative
to parturition,
TDik is the interaction effect of Liver Condition i by Time
Period k relative to parturition,
eijk is the unexplained residual assumed to be
independent and identically distributed N (0, s2).
Cow nested within liver condition was used in the random statement as the error term to test for treatment effects. Prepartum and postpartum data were analyzed separately for dry matter intake (DMI), body weight (BW), BW change, and DMI as a percentage of BW. Time period represented week relative to calving for production and plasma data (week 0 represents the week of calving) but represented day relative to calving for liver data.
The repeated statement including d or week relative to calving but no random statement was used in the program. Satterth option (performs a general Satterhwaite approximation for the denominator degrees of freedom) was included in the model statement.
For each analyzed variable, cow nested within treatment was subjected to four covariance structures: compound symmetric, heterogeneous compound symmetry, autoregressive order one, and unstructured variance components. The covariance structure that yielded the better fit statistics criteria was considered to be the most desirable analysis. Treatment differences or treatment by time interactions were considered to be significant at P < 0.05, to have a tendency toward significant at P = 0.05 to P = 0.12, and to be nonsignificant at P > 0.12. When treatment by time interactions were significant or tended to be significant, the slice option for LS MEANS statement in PROC MIXED [26] was used to determine for which time period means were different. Correlation analysis was performed to relate BC, ROL, and AT status with body fat mobilization. Pearson correlation coefficients were obtained using the PROC CORR procedure of SAS [26].
Results and Discussion
Mild fatty liver and feed intake
The number of cows with more than 2% TAG (wet weight) at d two postcalving was 19 among a cohort of 34 cows which gives a mild fatty liver prevalence of 56%. Mean concentration of TAG in the liver was 2.7 and 1.7 for fatty liver and non-fatty liver classifications (P = 0.008) (TABLE I). The pattern (interaction of liver condition by time from calving, P = 0.04) of liver TAG concentration was different between the two groups of cows (FIG. 1). At d 16 before calving, no differences in liver TAG concentration were observed between groups, but liver TAG concentration markedLy increased in the fatty liver group at d two and d 16 after calving.
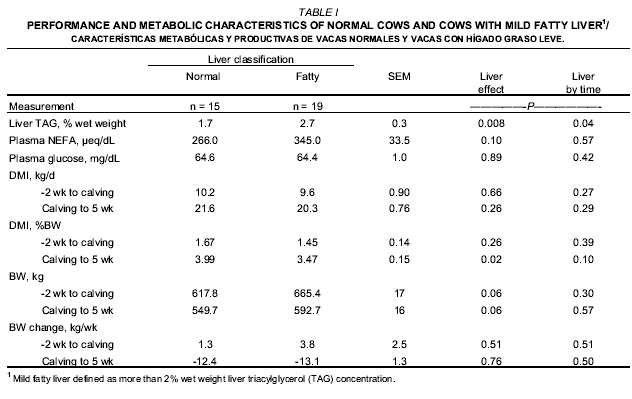

In periparturient cows, Muylle et al. [20] defined mild fatty liver as more than 2% liver TAG (wet basis). More recently, Bobe et al. [1] define mild fatty liver as 1 to 5% liver TAG, elevated concentrations of urinary ketones and decreased health status and reproductive performance. Although liver TAG concentration at d 16 before calving was similar, cows that developed fatty liver in this study had increased their liver TAG concentration by 125 and 98% by d two and 16 postpartum, respectively. The peak of liver TAG was observed at d two after calving which has been described previously [7, 9]. The persistence of fatty liver for the first two week of lactation was similar to previous reports by some [9, 27] but not by others [8, 24].
Plasma NEFA (P = 0.10) but not plasma glucose (P > 0.12) concentrations during the periparturient period tended to be greater for cows developing fatty liver (TABLE I). As previously described [20, 25], cows with fatty liver tended to have greater plasma NEFA concentration than normal cows. This means that cows with fatty liver had an increased rate of adipose tissue lipolysis than normal cows, even though BW changes did not vary between groups, before or after calving (TABLE I). The results of this study also showed that the plasma NEFA concentration was positively correlated with the liver TAG concentration at d two (r = 0.65; P < 0.001; not shown on Tables). Plasma glucose concentration was not different between fatty liver and non-fatty liver cows as previously reported [25].
Mean dry matter intake (kg/d) was not affected by the fatty liver condition as defined in this study during the peripartum period (-2 week to 5 week related to calving). Cows with fatty liver tended to have greater mean BW than cows with normal livers (P = 0.06) but no difference in body weight change (BWC) was observed between cow groups from two week before calving to five week of lactation (TABLE I). Cows with mild fatty liver consumed less DM as a percentage of BW than normal cows from calving to five week of lactation (P = 0.02), but no difference between groups were observed at week 1 and week 2 after calving (liver condition by week of lactation interaction, P = 0.10) (FIG. 2). As reviewed by Bobe et al. [1], no effects of mild fatty liver on DMI had been observed previously.

Plasma vitamins status
Because of decreased DMI as a %BW, increased liver TAG concentration, and increased plasma NEFA concentration observed in cows defined as having mild fatty liver, the changes in BC, ROL, and AT status could be associated with increased body fat mobilization and fatty liver development.
Similar to previous reports [5, 16], plasma concentrations of BC were decreased at calving and continued decreasing slightly during the first four week of lactation in both groups (FIG. 3). However, plasma BC concentrations were not different between groups of cows (P = 0.17) (TABLE II). The effect of lipid mobilization on plasma BC has been addressed before. Contrary to the obtained results, Haraszti et al. [10] reported a marked elevation of plasma NEFA (> 0.60 mmol/L) with a simultaneous decrease of plasma total carotene that exceeded 50% in cows between 30 to 45 d postpartum.


In the present study, plasma ROL concentrations increased slightly with week of lactation (FIG. 4) in both groups, but cows with mild fatty liver tended to have lower plasma ROL (P = 0.10) (TABLE II), as observed by others [10, 11] in cows with severe fatty liver. Plasma ROL concentrations were similar during the prepartum period but tended to be lower at week 0 and week two related to calving for cows with fatty liver as compared to normal cows (FIG. 4). Mean plasma ROL concentrations observed in this study are below concentrations (0.4 to 0.5 µg/mL) reported as typical when cows are fed at least 50,000 IU of vitamin A/d [28]. However, plasma concentrations of ROL are usually lower during the peripartum period [6, 16] as compared with other stages of the cycle. Recently, values under 0.2 µg/mL have been reported for periparturient Jersey cows fed 125,000 IU of supplemental retinyl palmitate [5] and for periparturient cows from different commercial herds [14]. Therefore, a poor relationship between intake of vitamin A and plasma ROL concentrations is frequently observed as pointed out by Weiss [28]. It has been suggested that the accumulation of TAG in the liver may reduce the synthesis of retinol-binding protein [15] and plasma albumin [18] which are both carriers for ROL from the liver to plasma.

Mudron et al. [19] have reported that plasma AT concentrations decreased in cows with mild fatty liver, but in the present study, plasma AT concentrations were not different between groups of cows (P = 0.97) (TABLE II). In both groups, plasma AT concentrations increased with wk of lactation (FIG. 5) as previously reported [6, 29].

Mean plasma AT concentrations (4.4 µg/mL) observed in this study were above the critical concentrations of 3.0 to 3.5 µg/mL [29] or 2.0 µg/mL [13] suggested for periparturient cows.
Liver vitamins status
No differences in mean liver BC and liver ROL between groups were observed (TABLE II). Liver BC (FIG. 6) values decreased over time in both groups of cows but the depression in liver BC at calving was more pronounced (68.0 vs. 8.0%, not shown in Tables) for normal than for fatty liver cows. At d 2 postpartum, liver BC concentration was greater (P = 0.04) in cows with fatty liver. Liver ROL concentrations (P = 0.10) tended to be greater in cows with fatty liver at -16 and 2 d relative to calving but no differences were observed at 16 and 30 d after calving (FIG. 7).


During the transition period, the ratio of liver ROL to liver BC was 174 and 165 for normal and fatty liver cows respectively, with results even higher than values observed in steers deprived of dietary BC [17].
Similar to findings reported by Mudron et al. [19] liver AT concentrations were higher (P = 0.06) for cows with more than 2% liver TAG concentrations (TABLE II). At d -16 (P = 0.03) and d two (P = 0.04) related to calving, liver AT concentrations were greater in cows with fatty liver but no differences were observed at 16 and 30 d after calving. In normal cows, liver AT values remains almost steady during the periparturient period (FIG. 8). For both groups of cows, liver stores were much higher for AT than for BC.

Dynamics of vitamins status and body fat mobilization
Plasma BC concentrations were related positively whereas both plasma ROL and plasma AT were related negatively to BW (P < 0.0001). Dry matter intake as a percentage of BW was highly and negatively related to plasma BC concentration but positively related to plasma ROL and plasma AT concentrations (P < 0.0001). A low but significant positive correlation between plasma NEFA and plasma BC concentrations (r = 0.18; P < 0.05) was detected in the present study. Both plasma ROL (r = -0.36) and plasma AT (r = -0.35) concentrations were related negatively to plasma NEFA concentration (P < 0.0001) (TABLE III).

Contrary to the obtained results, Haraszti et al [10] reported a negative correlation (r = -0.388 to -0.503) between plasma NEFA and plasma total carotene in cows at -5 to 10 d from calving. However, the correlation found between ROL and plasma NEFA in the present study was similar to Haraszti et al [10] whom reported a negative correlation (r = -0.360 to -0.430) between plasma NEFA and plasma vitamin A in cows at 0 to 45 d from calving. Similarly, at one week before calving, LeBlanc et al [14] found an inverse and significant association of plasma NEFA concentrations with serum ROL but not with serum BC concentrations. An increment of 100 µeq/dL in plasma NEFA would be expected to decrease ROL by 60 ng/mL.
In the present study, plasma BC concentrations seems to be closely related to degree of body fat mobilization as evidenced by a low but positive correlation between NEFA and plasma BC concentrations. Plasma concentrations of BC are related to forage intake, but no effect of feeding level (high vs. low energy intake) was recently observed on plasma BC concentrations of cows at mid lactation [22]. The negative correlation found between DMI, as percentage of BW, and plasma BC concentrations means that cows with lower DMI during the transition period have higher plasma BC concentrations.
A positive direct relationship between feed intake and plasma status of vitamins was found only for ROL (r = 0.55; P < 0.0001) and AT (0.46; (P < 0.0001), which suggests that stores of ROL or AT are less labile than stores for BC. For cows at mid lactation, the contribution of adipose BC pool to plasma BC was very small [22]. Based on present data and assuming a liver weight of 6 kg, the contribution of mobilized liver BC to plasma ROL (1 mg BC = 120 µg ROL) would be 1044 µg for normal vs. 115 µg for fatty liver cows, at calving.
The regulatory mechanisms of the liver in maintaining plasma ROL concentrations, particularly in dairy cows during the transition period are unknown. The lower liver BC and greater plasma ROL concentrations observed in this study for normal cows at calving suggests that stored ROL in liver first forces available BC stores to be converted into ROL, thus decreasing the BC concentration but not the ROL concentration in liver. However, these coordinated changes might not operate for cows with fatty liver.
There was a substantial variation in correlation coefficients among liver nutrients at each time from calving; therefore, a correlation coefficient for each time from calving was computed. Liver BC and liver ROL concentrations had a positive and significant correlation at d 30 (0.53; P < 0.05) but not at other times (P > 0.12) (TABLE IV) which suggests that the relationship between liver stores of BC and ROL appeared only after liver TAG infiltration declined (approximately 4 wk after calving). Positives correlations (range: 0.170 to 0.499) between liver total carotenes and liver vitamin A have been found in Hereford calves [3].
At d two from calving only, a positive correlation between liver BC and liver AT was found (P < 0.01) whereas no association between liver AT and liver ROL concentrations was observed (P > 0.12) at any time (TABLE IV).

At all times after calving, liver BC concentrations were positively (P < 0.12) associated with liver TAG concentrations but, liver ROL concentrations were not associated with liver TAG concentrations (TABLE IV). An association between liver AT and liver TAG concentrations was observed (0.43; P < 0.05) at d 30 from calving, only (TABLE IV).
No information on the relationship among liver BC, liver ROL and liver TAG was found in the literature and only one study has reported the relationships between liver AT and liver TAG [19]. The findings of the present study suggest that liver BC concentrations are related to liver TGA infiltration and fatty liver development.
Conclusions E IMPLICATIONS
During the transition from the nonlactating to the lactating state in the dairy cow, changes in b-carotene (BC) and vitamins A and E status occur. Late prepartum and early postpartum Holstein cows received a corn silage and alfalfa hay based diets. Changes in BC and vitamins A and E status in cows were associated with increased body fat mobilization and fatty liver development. Both plasma vitamins A and E had positives relationships with feed intake, which suggests that stores of these vitamins are less labile than stores for BC. Less liver BC was mobilized in cows with mild fatty liver at calving.
The changes in BC, ROL, and AT status in periparturient dairy cows were associated with increased body fat mobilization and fatty liver development. Plasma and liver BC would be expected to increase whereas both plasma ROL and AT would decrease with lipolysis. Both plasma ROL and AT had positives relationships with feed intake, which suggests that stores of ROL or AT are less labile than stores for BC.
The lower liver BC and greater plasma ROL concentrations found in this study for normal cows at calving suggest that stored ROL in liver first forces available BC stores to be converted into ROL, thus decreasing the BC concentration in liver without decreasing ROL concentration in liver. However, these coordinated changes might not operate for cows with fatty liver because less liver BC was mobilized in cows with mild fatty liver at calving. More information on the mechanisms of the liver for controlling homeostasis of plasma ROL is needed, particularly in dairy cows during the transition period.
ACKNOWLEDGMENT
This research was supported by the Florida Agricultural Experiment Station and a grant from DSM Vitamins Inc., Parsippany, NJ. Additional support provided by the Florida Dairy Checkoff.
Bibliographic references
1. BOBE, G.; YOUNG, J.; BEITZ, D. Pathology, etiology, prevention, and treatment of fatty liver in dairy cows. J. Dairy Sci. 87:3105-3124. 2004. [ Links ]
2. BROCAS, C.; RIVERA, R.; PAULA-LOPEZ, F.; MCDOWELL, L.; CALHOUN, M.; STAPLES, C.; WILKINSON, N.; BONING, A.; CHENOWETH, P.; HANSEN, P. Deleterious actions of gossypol on bovine spermatozoa, oocytes, and embryos. Biol. Reprod. 57:901-907. 1997. [ Links ]
3. DIVEN, R.; PAHNISH, O.; ROUBICEK, C.; ERWIN, E.; PAGE, H. Vitamin A and carotenoid inter-relationships in bovine plasma and liver. J. Dairy Sci. 43:1632-1638. 1960. [ Links ]
4. GOCHMAN, N.; SCHMITZ, J. Application of a new peroxide indicator reaction to the specific automated determination of glucose with glucose oxidase. Clin. Chem. 18:943-950. 1972. [ Links ]
5. GOFF, J.; KIMURA, K.; HORST, R. Effect of mastectomy on milk fever, energy, and vitamins A, E, and b–carotene status at parturition. J. Dairy Sci. 85: 1427-1436. 2002.
6. GOFF, J.; STABEL, J. Decreased plasma retinol, a-tocopherol, and zinc concentration during the periparturien period: Effect of Milk fever. J. Dairy Sci. 73:3195-3199. 1990. [ Links ]
7. GREENFIELD, R.; CECAVA, M.; DONKIN, S. Changes in mRNA expression for gluconeogenic enzymes in liver of dairy cattle during the transition to lactation. J. Dairy Sci. 83:1228-1236. 2000. [ Links ]
8. GRUM, D.; DRACKLEY, J.; HANSEN, L.; CREMIN, JR.; J. Production, digestion, and hepatic lipid metabolism of dairy cows fed increased energy from fat or concentrate. J. Dairy Sci. 79:1836-1849. 1996. [ Links ]
9. Grummer, R. R.; Bertics, S. J.; Hackbart, R. A. Short communication: Effects of prepartum milking on dry matter intake, liver triglyceride, and plasma constituents. J. Dairy Sci. 83:60-61. 2000. [ Links ]
10. Haraszti, J.; Huszenicza, g.; Molnár, l.; Blaskovits, a. Effect of periparturient lipid mobilization on serum total carotene and vitamin A concentrations in cattle. Acta Vet. Hungar. 32:193-203. 1984. [ Links ]
11. Hidiroglou, M.; Hartin, k. e. Vitamins A, E and selenium blood levels in the fat cow syndrome. Can. Vet. J. 23: 255-258. 1982. [ Links ]
12. Johnson, M. M.; Peters, J. P. Technical note: an improved method to quantify nonesterified fatty acids in bovine plasma. J. Anim. Sci. 71:753-756. 1993. [ Links ]
13. LeBlanc, S. J.; Duffield, T. F.; Leslie, K. E.; Bateman, K. G.; TenHag, J.; Walton, J. S.; Johnson, W. H. The effect of prepartum injection of vitamin E on health in transition dairy cows. J. Dairy Sci. 85:1416-1426. 2002. [ Links ]
14. LeBlanc, S. J.; Herdt, T. H.; Seymour, W. M.; Duffield, T. F.; Leslie, K. E. Peripartum serum vitamin E, retinol, and beta-carotene in dairy cattle and their associations with disease. J. Dairy Sci. 87:609-619. 2004. [ Links ]
15. Lindberg, L. A.; Sinkkonen, H.; Poso, A. R.; Tesfa, A. T.; Schroder, J. Production of monoclonal antibodies and enzyme immunoassay (EIA) to bovine retinol-binding protein (RBP) and determination of RBP serum levels and retinol concentrations in serum and liver in dairy cows before and after parturition. Res. Vet. Sci. 68:169-173. 1999. [ Links ]
16. Michal, J. J.; Heirman, L. R.; Wong, T. S.; Chew, B. P.; Frigg, M.; VolkER, L. Modulatory effects of dietary b-carotene on blood and mammary leukocyte function in periparturient dairy cows. J. Dairy Sci. 77:1408-1421. 1994.
17. Mora, O.; Romano, J. L.; González, E.; Ruiz, F. J.; Gómez, R.; Shimada. A. Presence of fed â-carotene in digesta, excreta, blood, and hepatic and adipose tissues of Holstein steers. Can. J. Anim. Sci. 81:133-139. 2001. [ Links ]
18. Mudron, P.; Rehage, J.; Qualmann, K.; Sallmann, H. P.; Scholz, H. A study of lipid peroxidation and vitamin E in dairy cows with hepatic insufficiency. J. Vet. Med. A 46: 219-224. 1999. [ Links ]
19. Mudron, P.; Rehage, J.; Sallmann, H. P.; Mertens, M.; Scholz, H.; Kovac, G. Plasma and liver a-tocopherol in dairy cows with left abomasal displacement and fatty liver. J. Vet. Med. A. 44: 91-97. 1997. [ Links ]
20. Muylle, E.; Van den Hende, C.; Sustronck, B.; Deprez, P. Biochemical profiles in cows with abomasal displacement estimated by blood and liver parameters. J. Vet. Med. A. 37: 259-263. 1990. [ Links ]
21. Njeru, C. A.; McDowell, L. R.; Shireman, R. M.; Wilkinson, N. S.; Rojas, L. X.; Williams, S. N. Assessment of vitamin E nutritional status in yearling beef heifers. J. Anim. Sci. 73:1440-1448. 1995. [ Links ]
22. Nozière, P.; Grolier, P.; Durand, D.; Ferlay, A.; Pradel, P.; Martin, B. Variations in carotenoids, fat-soluble micronutrients, and color in cows plasma and milk following changes in forage and feeding level. J. Dairy Sci. 89:2634-2648. 2006. [ Links ]
23. Rosendo, O.; McDowell, L. R. Liver dry matter and liver lipids in periparturient dairy cows. Acta Vet. Brno. 72: 541-546. 2003. [ Links ]
24. Rukkwamsuk, T.; Wensing, T.; Geelen, M. J. H. Effect of fatty liver on hepatic gluconeogenesis in periparturient dairy cows. J. Dairy Sci. 82:500-505. 1999a. [ Links ]
25. Rukkwamsuk, T.; Wensing, T.; Kruip, T. A. M. Relationship between triacylglycerol concentration in the liver and first ovulation in postpartum dairy cows. Theriogenol. 51: 1133-1142. 1999b. [ Links ]
26. STATISTICAL ANALYSIS SYSTEM INSTITUTE (SAS). Software Statistics, Version 8.2 Edition. Cary, NC. 2001.
27. Shaw, J. C. Studies on ketosis in dairy cattle. The effect of a vitamin A deficiency. J. Dairy Sci. 33:486-495. 1950. [ Links ]
28. Weiss, W. P. Requirements of fat-soluble vitamins for dairy cows: A review. J. Dairy Sci. 81:2493-2501. 1998. [ Links ]
29. Weiss, W. P.; Todhunter, D. A.; Hogan, J. S.; Smith, K. L. Effect of duration of supplementation of selenium and vitamin E on periparturient dairy cows. J. Dairy Sci. 73:3187-3194. 1990. [ Links ]
