










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Investigación
versión impresa ISSN 1010-2914
Revista de Investigación vol.33 no.68 Caracas dic. 2009
Las Costras Microbióticas del Suelo
The Microbiotic Crust On Soil
Valentina Toledo1 Adriana Florentino2
(1) Universidad Pedagógica Experimental Libertador. Instituto Pedagógico de Caracas. Caracas, Venezuela. toledo.valentina@gmail.com
(2) Universidad Central de Venezuela. Instituto de Edafología, Facultad de Agronomía. Caracas, Venezuela. florentinoa@agr.ucv.ve
RESUMEN
La presente investigación tuvo como propósito presentar el estado del conocimiento de un tópico relacionado con la ecología microbiana del suelo como son las costras microbióticas. Se refiere a una investigación documental partiendo de la terminología empleada para describirla, su formación y funciones en el suelo, así como tema de investigación en diferentes áreas de la ciencia. Dentro de las conclusiones tenemos que las costras microbióticas mitigan la degradación de los suelos y tienen un efecto de biofertilizante natural. Su papel ecológico es reseñado en diferentes investigaciones y en distintas áreas de la ciencia. Sin embargo, poco se conoce en el ámbito educativo. Escasamente se discuten resultados de suelos recuperados por inoculación de microorganismos o por transferencia de fragmentos de costras microbióticas, lo cual se perfila como una oportunidad para desarrollarla a mediano plazo en nuestro país.
Palabras clave: Costra microbiótica; cianobacterias; musgos; líquenes; suelos
ABSTRACT
The present investigation present the state of the knowledge of a topic related with the microbial ecology in the soil like they are the microbiotic soil crust. The work is developed through a documental investigation leaving of the terminology used to describe it, its formation and functions in the soil, as well as investigation topic in different areas of the science. Inside the summations we have that the microbiotic soil crust mitigate the degradation in the soil and they have an effect of natural biofertility. Its ecological rol is reviewed in diverse investigations and different areas from science. However, little it is known in the educational environment. Scarcely results of recovered in the soil are discussed by inoculation of microorganisms or for transfer of fragments of microbiotic soil crust, that which is profiled like an opportunity to develop it to medium term in our country.
Key words: Microbiotic soil crust; cyanobacteria; mosses; lichens; soil
INTRODUCCIÓN
Las costras microbióticas son diminutas comunidades pero organismos importantes que pueden incluir cianobacterias, algas, musgos, líquenes y otros, los cuales se encuentran asociados íntimamente con las partículas minerales de la superficie del suelo, creando una capa delgada, cohesiva y horizontal (Belnap y Lange, 2001). Estas comunidades son comunes en ecosistemas con alta entrada de luz hacia la superficie del suelo, tal como ocurre en las zonas áridas del mundo (Bowker, 2007).
En contraste con las costras físicas o inorgánicas, las cuales se forman por el impacto de las gotas de lluvia sobre la superficie, formando un sello superficial que impide la infiltración y promueve la escorrentía, con la consecuente erosión y pérdida de nutrientes del suelo, las costras microbióticas enfatizan la calidad del suelo por: 1) agregación de las partículas del suelo, reduciendo así la erosión hídrica y eólica; 2) mejora la infiltración del agua a través del suelo debido al incremento de la estabilidad de los agregados (unidad de partículas coherente del suelo) y 3) aumenta la fertilidad del suelo por fijación de nitrógeno y carbono.
La costra microbiótica ha sido reportada en las regiones áridas y semiáridas de diferentes partes del mundo: USA (Rychert y Skujins, 1974; Klubek y Skujins, 1980; Cole, 1990; Beymer y Klopatek, 1991; Belnap y Gardner, 1993), en Australia (Eldridge, 1993), en China (Reynaud y Lumpkin, 1988), en Israel (Verrecchia et al., 1995), en Africa Saheliana (Malam, et al., 1999; Malam et al., 2001a), en Brasil (Trindade et al., 2001) y en el Ecuador (Pérez, 1997). Aunque las costras han sido encontradas en los ecosistemas árticos, boreales y áridos, es en los últimos donde aparentemente tienen mayor importancia ecológica, ya que pueden llegar a cubrir hasta 70% de la superficie (representando más del 30% de la superficie terrestre) (Belnap y Lange, 2001).
Por otra parte, se evidencia a través de la bibliografía consultada, un incremento importante en las investigaciones sobre los roles de estos macro y microorganismos en los diversos ecosistemas a nivel mundial. Dentro de los beneficios ecológicos que aporta la presencia de la costra en el suelo, diversos autores incluyen, desde su efecto como biofertilizante natural pasando como acondicionador del suelo, biorremediador, hasta la recuperación de suelos degradados en fase intermedia por inoculación de microorganismos (Belnap, Hawkes y Firestone, 2003; Bowker, Belnap, Davidson y Goldstein, 2006).
Actualmente, el deterioro progresivo de los suelos como consecuencia del mal manejo de la tierra ante la demanda de consumo de alimentos, pone de manifiesto la degradación de los suelos, representado en la desertificación en diferentes zonas. Este problema avanza de forma muy rápida y afecta con mayor intensidad las zonas áridas y semiáridas del mundo. A pesar de esto, escasamente se discuten resultados de suelos recuperados por inoculación de microorganismos o por transferencia de fragmentos de costras microbióticas, lo cual se perfila como una oportunidad para desarrollarla a mediano plazo.
En consecuencia, el propósito de este trabajo es presentar las costras microbióticas como un tema relacionado con ecología microbiana del suelo, así como también los beneficios obtenidos de su estudio en diversos ecosistemas, los cuales son escasos en nuestro país pero ampliamente distribuido a nivel mundial y finalmente, sugerir futuras investigaciones de las costras microbióticas en Venezuela.
MÉTODO
Esta investigación está sustentada en un estudio documental, con el propósito de presentar el estado del conocimiento de un tópico, las costras microbióticas del suelo. Para focalizar el progreso de la investigación actual, reflexiones y recomendaciones, se partió de la revisión de trabajos previos, entre ellos libros, investigaciones y artículos científicos divulgados por medios impresos y electrónicos. La selección de estas fuentes documentales, evidencia la evolución de los estudios de costras microbióticas en el suelo, la cual esta densamente documentada en las regiones de América del Norte, Europa, en la India, poco en Australia y escasamente en Venezuela. Las investigaciones de los roles de las costras microbióticas del suelo en el contexto Venezolano, aparecen fragmentadas o discontinuas en el tiempo y con poco énfasis en el desarrollo ecológicamente sustentable en las regiones áridas y semiáridas en Venezuela.
La revisión de la amplia bibliografía consultada permitió, en primera instancia, tener un marco de referencia hacia donde se han orientado las investigaciones en costras microbióticas y posteriormente destacar las oportunidades inexploradas para sus futuros estudios en Venezuela.
Terminología empleada para describir la costra macrobiótica.
La costra microbiótica está formada por organismos vivos y sus productos, creando una costra de partículas inorgánicas atrapadas en la superficie del suelo junto con materiales orgánicos (USDA, 1997). Es una superficie biológica dominante en los paisajes áridos y semiáridos, siendo un complejo ensamblaje de musgos, cianobacterias, líquenes, hongos y plantas no vasculares (Eldridge y Greene, 1994). (Figura 1). También es conocida como costra microfloral (Loope y Gifford, 1972), criptogámica (Kleiner y Harper,1977), microfísica (West, 1990) y criptobiótica (Harper y Pendleton, 1993; Belnap, 1993), lo cual ha generado confusión en la terminología empleada. También se conocen como biofilm, tapetes microbianos o costra biológica. Todos los nombres encontrados indican las formas de los organismos comunes que componen la costra, inclusive, el término más actualizado es "microbiótica", en donde Downing y Selkirk (1993), la refieren al pequeño tamaño de los organismos sin limitar los componentes de la costra y sus funciones. No obstante, independientemente del nombre que se use, en todas estas definiciones se establece una clara distinción entre estas formaciones y las costras físicas o físico-químicas.
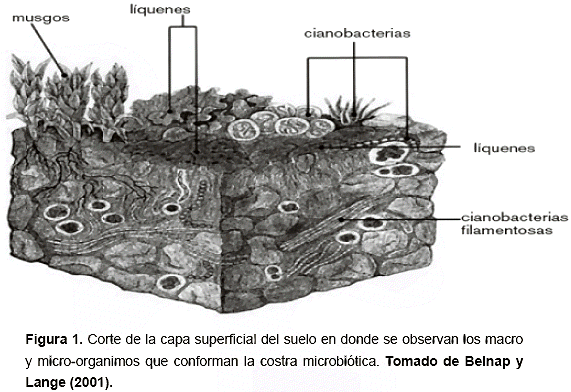
La costra microbiótica está formada por organismos vivos y por sus productos, es decir, por materiales orgánicos; mientras que las costras físicas y químicas son fundamentalmente inorgánicas. Por tanto, la costra microbiótica resulta de la colonización de la superficie del suelo por comunidades de microorganismos dominadas por cianobacterias y algas eucarióticas, pero también es común la presencia de plantas no vasculares o macroorganismos, musgos y hongos.
La confusión sobre la definición de los diferentes tipos de costra microbiótica se debe posiblemente a la amplia disparidad de los resultados reportados en la bibliografía australiana, particularmente en el rol que ésta desempeña en el proceso de infiltración. No obstante, Eldridge y Greene (1994), proponen un sistema alternativo para clasificar las costras microbióticas sin considerar el sistema de clasificación de planta /animal, considerando tres tipos de forma: hipermórfi cas, perimórfi cas y criptomórficas (Figura 2).

La costra hipermórfica incluye musgos, los cuales ocurren en microhábitas húmedos de las regiones semiáridas, así como debajo de los arbustos, siempre asociadas con algas. Son comunes en pendientes estables con microrrelieve pronunciado y son más vulnerables a las perturbaciones como el pastoreo y el fuego. Su influencia está limitada a las áreas sobre la superficie del suelo, de allí su habilidad moderada contra el efecto del viento y la acción del impacto de las gotas de lluvias.
La costra perimórfica compuesta por líquenes foliáceos (como una hoja) y crustáceos (como una corteza), los cuales constituyen una asociación simbiótica de hongos y microalgas eucarióticas (aplicado o relativo a las células que tienen núcleo con membrana, aparato de Golgi y mitocondrios). Su área de influencia está repartida por encima y por debajo de la superficie del suelo y proporciona protección al suelo contra la erosión por el viento y el agua. La asociación de hifas de hongos con el liquen crea una red debajo de la superficie y fortalece el desarrollo de agregados estables al suelo.
La costra criptomórfica microscópica está compuesta de microalgas eucarióticas y filamentos de hongos; se encuentra en lugares perturbados y es menos susceptible al fuego. Su mayor actividad está concentrada (en grupos) debajo de la superficie donde ellas contribuyen a mejorar la estructura del suelo y el status nutricional.
Otros sistemas de clasificación para las costras han sido propuestos por (Belnap, Kaltenecker, Rosentreter, Williams, Leonard y Eldridge, 2001); este último está basado en la morfología externa de la costra, siendo más fácil distinguirla en forma visual y su efecto en mayor proporción sobre las funciones de muchos ecosistemas. En esta clasificación, se reconocen cuatro grupos, a saber: lisas, rugosas, enrolladas y en pináculos.
Formación de la costra microbiótica
Las costras microbióticas y las físicas constituyen los principales tipos de costras del suelo en las regiones áridas y semiáridas. Las costras microbióticas ocurren comúnmente en asociación con las costras físicas del suelo, las cuales constituyen el soporte inicial para que los microorganismos inicien la colonización (Toledo, 2006). Estos tapetes microbianos están constituidos por densas comunidades de microorganismos que se disponen siguiendo un patrón de estratificación vertical (Figura 3). Dicha estratificación se hace aparente a menudo por la diferente coloración de las distintas capas, que es consecuencia del tipo de pigmentos de los organismos situados en ellas. Las comunidades microbianas se entremezclan en los tapetes con partículas de sedimento que son atrapadas o precipitadas por su propios productos (Hu, Zhang, Huang y Liu, 2003; Camacho y Fernández-Valiente, 2005; Toledo 2006).

En general, la apariencia de las costras en términos de color, superfi cie topográfica y cobertura superficial varía en diferentes regiones. Su presencia está asociada a la textura del suelo, conductividad eléctrica, pH, humedad y posiblemente temperatura (USDA, 1997). Por otro lado, la morfología externa de la costra está determinada por múltiples factores, dentro de los cuales el clima y el registro histórico de uso del suelo son probablemente los más importantes, seguidos por la textura y la composición química del suelo (Belnap, 2001).
Además, diversos estudios sugieren que los cambios microclimáticos y edáficos promovidos por el desarrollo de la vegetación pueden favorecer el desarrollo de determinados componentes de la costra microbiótica, que de otra forma no podría sobrevivir a las adversas condiciones climáticas presentes en las zonas desnudas (Belnap et al., 2001, Maestre, 2003). Alternativamente, los macroorganismos (briofitasmusgos y líquenes) presentan adaptaciones morfológicas tolerantes a la desecación. En particular, las briofitas que habitan áreas áridas y semiáridas exhiben estrategias para resistir la sequía, abarcando una variedad de ajustes estructurales para la retención de agua y la manutención de la fotosíntesis durante períodos de aridez (Kürschner, 2003). En este sentido, los componentes de la costra pueden permanecer en estado latente ante eventos extremos de sequía sin presentar efectos negativos y activar sus funciones metabólicas rápidamente con pequeñas cantidades de humedad atmosférica (punto de rocío) o con cierto contenido de agua de la superficie del suelo, de allí que se conocen como organismos poiquilohídricos (Belnap, Kaltenecker, Rosentreter, Williams, Leonard y Eldridge, 2001).
Funciones de la costra microbiótica en el suelo
Diversas investigaciones destacan el rol que la costra microbiótica desempeña en la agregación de las partículas del suelo con la consecuente protección contra la erosión hídrica y eólica (Campbell et al., 1989; Belnap y Gardner, 1993; Malam et al., 2001a; Trindade et al., 2001), en la infiltración del agua y escurrimiento superficial (Eldridge y Greene, 1994). Con relación a esto, Loope y Gifford (1972) reportaron que la costra microbiótica contribuye a la infiltración del agua y disminuye la escorrentía. No obstante, Brotherson y Rushforth (1983), Verrechia et al (1995), Kidron et al. (1999) y Cerdá (2002), sugieren lo opuesto. La costra también aumenta la retención de humedad del suelo (Belnap y Gardner, 1993; Verrecchia et al., 1995), incrementa la disponibilidad de nutrientes al concentrar ciertos elementos esenciales, así como también el N, el cual es fijado por las cianobacterias (Harper y Pendleton, 1993; Stal, 1995; Zaady et al., 1998; Malam et al., 2001b), y el carbono (Zaady et al., 2000) y favorece la germinación de semillas (Zaady et al.,1997). Actualmente, las costras microbióticas están siendo utilizadas en la biorremediación de zonas afectadas por vertidos accidentales de petróleo (Martínez-Alonso, Gaju, 2005). En consecuencia, son aprovechadas como bioindicador de la salud del suelo no sólo por las ventajas de su uso en la recuperación de zonas deterioradas o degradadas sino también por sus efectos como fertilizante natural.
La costra microbiótica, como tópico de investigación, en diversas áreas de la ciencia
Los factores más importantes que determinan el establecimiento y mantenimiento de la costra microbiótica son: el sustrato, el microrelieve y el patrón de perturbación, mientras que su distribución está relacionada principalmente con los grupos morfológicos o especímenes que la componen (Eldridge y Rosentreter, 1999; DeFalco et al., 2001).
Basado en lo anteriormente expuesto, un número creciente de investigaciones se ha orientado a evaluar la costra microbiótica en los ecosistemas áridos y semiáridos, abarcando desde su composición, pasando por las distintas funciones ecológicas y finalmente, la estimulación de su formación mediante la inoculación con microorganismos autóctonos; esto último, con el fin de promover el crecimiento de la costra sobre los suelos degradados por la erosión hídrica (Kumar y Thomas 1997; Zaady, et al., 1997; Shem-Tov et al., 2002).
Asimismo, a través de la revisión bibliográfica se constata que la costra microbiótica aparece como tópico de investigación en diversas áreas de la ciencia, particularmente en biología, botánica, ecología y física de suelos. Algunos trabajos resaltan la actividad biológica de un determinado microorganismo o macroorganismo (San José y Bravo, 1991), mientras que otros destacan algunas de las funciones de la costra en el suelo (Budel, Luttge, Stelzer, Huber, y Medina, 1994; Gorbushina, Boettcher, Krumbein y Vendrell-Sanz, 2001). En algunos casos, los distintos métodos empleados para su estudio son similares a los utilizados para diagnosticar propiedades físicas, químicas o biológicas en suelos de uso agrícola, sin costra microbiótica y también en suelos naturales. Sin embargo, cada vez más los métodos para su estudio se perfilan hacia el uso de técnicas que permiten su observación en dos (2D) y principalmente en tres (3D) dimensiones; entre estas técnicas, interesa destacar la de microscopía electrónica de barrido (MEB) y microanálisis de rayos X (EDX), las cuales permiten un estudio submicroscópico instrumental, logrando con esto explicar el funcionamiento de este ecosistema a pequeña escala para predecir y describir procesos que ocurren a escalas mayores (Asunción de los Ríos et al., 2004; Toledo y Urbina de Navarro, 2008).
Por otro lado, son pocos los trabajos de investigación que articulan las propiedades biológicas y bioquímicas con las físicas en los estudios de la costra microbiótica, siendo los dos primeros grupos de propiedades los que van a resultar más sensibles a los cambios que se producen en el suelo, y este es un aspecto a tener en cuenta cuando se manejan suelos de clima semiárido sometidos a procesos de degradación o desertificación (Toledo, 2008). Así mismo, la mayoría de las investigaciones destacan una determinada función de la costra microbiótica, la cual responde frecuentemente a un macroorganismo o microorganismo que la compone pero muy poca veces aparece descrita una costra compuesta por varios macroorganismos o microorganismos (Toledo, 2006).
Como resultado, en algunos desiertos norteamericanos, como el de Mojave y la Meseta de Colorado, se ha estimado que las cianobacterias y los líquenes tardan en recuperarse de 35 a 65 años y de 45 a 85 años respectivamente, mientras que el tiempo necesario para los musgos puede ser hasta de 250 años (Eldridge y Rosentreter, 1999). Probablemente, sea esta la razón como la señala Bowker (2007), que raramente se discute en las investigaciones la restitución de la costra microbiótica como una técnica en la rehabilitación de suelos expuestos a diferentes perturbaciones tales como: degradación por erosión hídrica o eólica, pastoreo, actividad agrícola, que implica pérdida de la calidad del mismo.
Aún así, su presencia en suelos desnudos y de forma muy discontinua ofrecerá mayor beneficio para el suelo y su preservación como manto orgánico a que no existiera, pues actúa de trampa de propágalos y promueve la concentración de recursos, lo cual actúa como un fertilizante natural en los ecosistemas más precarios (Li, Wang, Ti, Zhang, 2002; Bowker et al. 2006).
Futuras investigaciones
La información de la costra microbiótica en Venezuela esta basada en una pequeña cantidad de investigaciones en diversos ecosistemas. Se necesita más estudios orientados a: 1) Cómo la composición de la costra y sus funciones varía con el clima y la química del suelo. Para ello es necesario disponer de estaciones climatológicas dotadas con instrumentos para el registro de parámetros microclimáticos (punto de rocío, humedad específica, entre otros), los cuales permitirán reconocer la importancia de los umbrales críticos en la disponibilidad de un recurso limitante, en este caso el agua y para la conservación de la costra microbiótica; 2) Evaluar el valor potencial de la costra microbiótica como biofertilizante para acelerar la recolonización del suelo en zonas con evidencia de desertifi cación a través de invernaderos experimentales (en laboratorio), bien en forma de translocación (transferencia de fragmentos de costra) o mediante la dispersión de inóculos. 3) Fomentar la importancia de la costra microbiótica para las zonas áridas y semiáridas intertropicales a través de la divulgación de documentos técnicos y científi cos, zonificando el área donde ésta se encuentra, como un lugar de experiencia de laboratorio en campo.
CONCLUSIONES
Las costras microbióticas figuran como una alternativa para la recuperación de estas zonas deterioradas ya que crecen de forma natural, preferiblemente en los suelos desnudos, por lo que pueden contribuir a estabilizar el suelo y disminuir el riesgo de la erosión, en sectores con evidencias de desertificación.
Las costras microbióticas son importantes para el ciclo de nutrientes y protección del suelo contra la erosión hídrica y eólica. Son zonas de concentración de elevados recursos (nutrientes, materia orgánica) o propágulos (esporas en los musgos, esporas de hongos, etc.) y en la progresiva formación de islas o parches de fertilidad.
En la actualidad, la mayor amenaza para la Costra Microbiótica es la pérdida de extensión, debido a que las áreas intactas remanentes se están convirtiendo en terrenos para la agricultura y el comercio.
El conocimiento acerca de la composición específica y la ecología de la costra microbiótica en nuestro país es aún incipiente. El estudio de éstos y otros aspectos relacionados con el monitoreo de estas comunidades biológicas es esencial, no sólo para entender el funcionamiento de estas en cualquier ecosistema sino también para contribuir a su conservación.
REFERENCIAS
1. Asunción de Los Ríos, C., Wierzchos, J. Valiente, E. y Quesada, A. (2004). Microstructural characterization of cyanobacterial mats from the McMurdo Ice Shelf, Antarctica. Applied and Environmental Microbiology, 70(1), 569-580. [ Links ]
2. Belnap, J. (1993). Recovery rates of cryptobiotic crusts: inoculant use and assessment methods. Great Basin Naturalist, 53, 89-95. [ Links ]
3. Belnap, J. y Gardner, J. (1993). Soil microstructure in soils of the Colorado Plateau: the role of the cyanobacterium Microcoleus vaginatus. Great Basin Naturalist, 53, 40-47. [ Links ]
4. Belnap, J. (2001). Comparative structure of physical and biological crusts. In J. Belnap y O.L. Lange (ed.), Ecological Studies: Vol. 150. Biological Soil Crusts: Structure, Function and Management (pp. 177-191). New York: Springer-Verlag. [ Links ]
5. Belnap, J. y Lange, O. (2001). Structure and functioning of biological soil crusts: a synthesis. In J. Belnap y O.L. Lange (ed.), Ecological Studies: Vol. 150. Biological Soil Crusts: Structure, Function and Management (pp. 471-479). New York: Springer-Verlag. [ Links ]
6. Belnap, J., Kaltenecker, J. Rosentreter, R. Williams, J. Leonard, S. y Eldridge, D. (2001). Biological soil crusts: ecology and management. BLM Technical Reference 1730-2. Washington, D.C: United States Bureau of Land Management. [ Links ]
7. Benalp, J., Hawkes, C. y Firestone, M. (2003). Boundaries in miniature: Two examples from soil. BioScience, 53(8), 739-749. [ Links ]
8. Beymer, R. y Klopatek, J. (1991). Potencial contribution of carbon by microphytic crusts in Pinyon-Juniper woodlands. Arid Soil Research and Rehabilitation, 5:187-198. [ Links ]
9. Bowker, M., Belnap, J., Davidson, D. y Goldstein, H. (2006). Correlates of biological soil crust abundance across a continuum of spatial scales: support for a hierarchical conceptual model. Journal of Applied Ecology, 43, 152-163. [ Links ]
10. Bowker, M. (2007). Biological soil crust rehabilitation in theory and practice: An underexploited opportunity. Restoration Ecology, 15(1), 13-23. [ Links ]
11. Brotherson, R. y Rushforth, S. (1983). Influence of cryptogamic crust on moisture relationships of soils in Navajo National Monument, Arizona. Great Basin Naturalist, 43, 73-78. [ Links ]
12. Budel, B., Luttge, U. Stelzer, R. Huber, O. y Medina, E. (1994). Cynobacteria of rocks and soils of the Orinoco lowlands and the Guayana uplands, Venezuela. Journal of Plant Physiology, 144, 74-79. [ Links ]
13. Camacho A., y Fernández-Valiente E. (2005). Un mundo dominado por los microorganismos. Ecología microbiana de los lagos antárticos. Ecosistemas. [Revista en línea], 2. Disponible http://www.revistaecosistemas.net/articulo.asp?Id=109 & Id_Categoria=2 & tipo=portada (Consulta: 2007, Diciembre 6). [ Links ]
14. Campbell, S., Seeler, J. y Golubic, S. (1989). Desert crust formation and soil stabilization. Arid Soil Research and Rehabilitation, 3, 217-228. [ Links ]
15. Cerdá, A. (2002). Influencias de las costras biológicas en el comportamiento hidrológico y erosivo de los suelos en los cordones dunares de Nizzana, desierto del Negev, Israel (Documento en línea). Disponible: http://www.geot.unex.es/norba/files/10/p3.pdf (Consulta: 2003, Abril 23). [ Links ]
16. Cole, D. (1990). Tramplind disturbance and recovery of cryptogamic soil crust in Gran Canyon National Park. Great Basin Naturalist, 50, 321-325. [ Links ]
17. Defalco, L., Detling, J. Richard, C. y Warren, S. (2001). Physiological variation among and exotic winter annual plants associated with microbiotic crust in the Mojave Desert. Plant and Soil, 234, 1-14. [ Links ]
18. Downing, A. y Selkirk, P. (1993). Bryophytes of the calcareous soils of Mungo National Park, an arid area of southern central Australia. Great Basin Naturalist, 53(1): 13-23. [ Links ]
19. Eldridge, D. (1993). Cryptogams, vascular plants, and soil hydrological relations: some preliminary results from the semiarid woodlands of eastern Australia. Great Basin Naturalist, 53, 48-58. [ Links ]
20. Eldridge, D. y Greene, R. (1994). Microbiotic soil crust: A review of their roles in soil and ecological processes in the Rangelands of Australia. Australian Journal Soil Research, 32, 389-415. [ Links ]
21. Eldridge, D. y Rosentreter, R. (1999). Morphological groups: a framework for monitoring microphytic crusts in arid landscapes. Journal of Arid Environments, 41, 11-25. [ Links ]
22. Gorbushina, A., Boettcher, M. Brumsack, H. Krumbein, W. y Vendrell-Sanz, M. (2001). Biogenic forsterite and opal as a product of biodeterioration and lichen stromatolite formation in table mountain systems (Tepui) of Venezuela. Geomicrobiology Journal, 18 (1): 117-132. [ Links ]
23. Harper, K. y Pendleton. R. (1993). Cyanobacteria and cyanolichens: can they enhance availability of essential minerals for higher plants?. Great Basin Naturalist, 53, 59-72. [ Links ]
24. Hu, C., Zhang, D, Huang, Z. y Liu, Y. (2003). The vertical microdistribution of cianobacteria and green algae within desert crust and the development of the algal crust. Plant and Soil, 257, 97-111. [ Links ]
25. Kidron, G., Yaalon, D. y Vonshak, A. (1999). Two causes for runoff initiation on microbiotic crust: hydrophobicity and pore clogging. Soil Science, 164, 18-27. [ Links ]
26. Kleiner, E y Harper, K. (1977). Soil properties in relation to cryptogamic ground cover in Canyonlands National Park. Journal of Range Management, 30, 203-205. [ Links ]
27. Klubek, B. y Skujins, J. (1980). Heterotrophic N2 fixation in arid soil crust. Soil Biology and Biochemistry, 12, 229-236. [ Links ]
28. Kumar, S. y Thomas, J. (1997). Possible amelioration of coastal soil salinity using halotolerant nitrogen-fixing cyanobacteria. Plant and Soil, 189, 205-211. [ Links ]
29. Kürschner, H. (2003). Life strategies and adaptations in bryophytes from the near and middle east. Turk Journal Botanic, 28, 73-84. [ Links ]
30. Li, X., Wang, X. P. Li, T y Zhang, J. (2002). Microbiotic soil crust and its effect on vegetation and habitat on artificially stabilized desert dunes in Tengger Desert, North China. Biology and Fertility Soils, 35, 147-154. [ Links ]
31. Loope, W. y Gifford, G. (1972). Influence of a soil microfl oral crust on select properties of soils under Pinyon-Juniper in southeastern Utah. Journal Soil Water Conservation, 27, 164-167. [ Links ]
32. Malam, O., Trichet, J. Defarge, C. Coute, A. y Valentin, C. (1999). Morphology and microstructure of microbiotic soil crusts on a tiger bush sequence (Niger, Sahel). Catena, 37, 175-196. [ Links ]
33. Malam, O., Le Bissonnais, Y. Défarde, C. y Trichet, J. (2001a). Role of a cyanobacterial cover on structural stability of sandy soils in the Sahelian part of western Niger. Geoderma, 101, 15-30. [ Links ]
34. Malam, O., Stal, L. Défarge, C. Couté, A y Trichet, J. (2001b). Nitrogen fixation by microbial crust from desiccated Sahelian soils (Niger). Soil Biology and Biochemistry, 33, 1425-1428. [ Links ]
35. Maestre, F. (2003). Variaciones en el patrón espacial a pequeña escala de los componentes de la costra biológica en un ecosistema mediterráneo semiárido. Revista Chilena de Historia Natural, 76(1), 35-48. [ Links ]
36. Martínez-Alonso M. y Gaju, N. (2005). El papel de los tapetes microbianos en la biorrecuperación de zonas litorales sometidas a la contaminación por vertidos de petróleo. Ecosistemas. (Revista en línea), 2. Disponible: http://www.revistaecosistemas.net/articulo.aspId=122 & Id_Categoria=2 & tipo=portada (Consulta: 2007, Diciembre 6). [ Links ]
37. Pérez, F. (1997). Microbiotic crust in the high equatorial Andes, and their influence on paramo soils. Catena, 31, 173-198. [ Links ]
38. Reynaud, P. y Lumpkin, T. (1988). Microalgae of the Lanzhou (China) crytogamic crust. Arid Soil Research and Rehabilitation, 2, 145-155. [ Links ]
39. Rychert, R. y Skujins, J. (1974). Nitrogen fixation by blue-green alga-lichen crusts in the Great Basin Desert. Soil Science Society American Proceedings, 38, 768-771. [ Links ]
40. San José, J. y Bravo, C. (1991). CO2 Exchange in soil algal crusts ocurring in the trachypogon savannas of the Orinoco Llanos, Venezuela. Plant and Soil, 135, 233-244. [ Links ]
41. Shem-Tov, S., Zaady, E. y Gutterman, Y. (2002). Germination of Carrichtera annua (Brassicaceae) seeds on soil samples collected along a rainfall gradient in the Negev Desert of Israel. Israel Journal of Plant Sciences, 50, 113-118. [ Links ]
42. Stal, L. (1995). Physiological ecology of cyanobacteria in microbial mats and other communities. New Phytologist, 131, 1-32. [ Links ]
43. Toledo, V. (2006). Caracterización de la costra microbiótica y su influencia biológica y física en suelos de la región árida de Quibor, estado Lara. Tesis de doctorado no publicada. Universidad Central de Venezuela, Postgrado en Ciencia del Suelo. Facultad de Agronomía, Maracay. [ Links ]
44. Toledo, V y Navarro de Urbina, C. (2008). Estudio preliminar de la influencia de la costra microbiótica en los suelos de Quibor, estado Lara, Venezuela, mediante microanálisis de rayos –X (EDS). Acta Microscópica, 17(1), 77-84. [ Links ]
45. Toledo, V. (2008). Evaluación de las propiedades biológicas y bioquímicas de la costra microbiótica para la recuperación de suelos degradados en la región árida de Quibor, Venezuela. Trabajo de ascenso no publicado, Universidad Pedagógica Experimental Libertador, Instituto Pedagógico de Caracas, Caracas. [ Links ]
46. Trindade, E., Schaefer, C. Albuquerque, M. Abrahao, W. Mello, J. y Chagas, A. (2001). Costas biológicas em saprolitos de gnaisse: ciclagem biogeoquímica, micromorfologia e ensaio de colonizacao. Revista Brasileira do Ciencia do Solo, 25, 849-861. [ Links ]
47. USDA. (1997). Introduction to microbiotic crust. Washington, D.C.: Secretary of Agriculture, U.S. Department of Agriculture. 13. [ Links ]
48. Verrecchia, E., Yair, A. Kidron, G. y Verrecchia, K. (1995). Physical properties of the psammophile cryptogamic crust and their consequences on the water regime of sandy soils, North-western Negev Desert, Israel. Journal Arid Environment, 29, 427-437. [ Links ]
49. West, N. (1990). Structure and function of microphytic soil crusts in wildland ecosystems of arid to semi-arid regions. Advance Ecology Research, 20, 179-223. [ Links ]
50. Zaady, E., Groffman, P. y Shachak, M. (1998). Nitrogen fixation in macro and microphytic patches in the negev desert. Soil Biology and Biochemistry, 30, 449-454. [ Links ]
51. Zaady, E., Gutterman, Y. y Boeken, B. (1997). The germination of mucilaginous seeds of Plantago coronopus, Reboudia pinnata, and Carrichiera annua on cyanobacterial soil crust from the Negev Desert. Plant and Soil, 190, 247-252. [ Links ]
52. Zaady, E., Kuhn, U. Wlske, B. Sandoval-Soto, L y Kesselmeier, J. (2000). Patterns of CO2 exchange in biological soil crust of successional age. Soil Biology and Biochemistry, 32, 959-966. [ Links ]
