











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de la Sociedad Venezolana de Microbiología
versión impresa ISSN 1315-2556
Rev. Soc. Ven. Microbiol. v.22 n.1 Caracas ene. 2002
Efecto de inhibidores de ß-lactamasas sobre la evolución de la resistencia a ß-lactámicos en bacilos Gram-negativos.
Effect of ß-lactamases inhibitors on the evolution of resistance to ß-lactams in Gram-negatives bacilli.
Martín N., G.1; Carmona, O.2; Guzmán Blanco, M.3 y Grupo Venezolano de Vigilancia de la Resistencia Bacteriana (GVRB).
1. Cátedra de Farmacología, Escuela de Medicina "J. M. Vargas", Facultad de Medicina, Universidad Central de Venezuela, Caracas, Venezuela.
2. Cátedra de Microbiología, Escuela de Medicina "J. M. Vargas", Facultad de Medicina, Universidad Central de Venezuela, Caracas, Venezuela.
3. Unidad de Microbiología y Enfermedades Infecciosas del Hospital Vargas, Caracas, Venezuela.
Resumen
Los ß-lactámicos, primeros antibióticos introducidos para uso en clínica, siguen siendo los más usados. También es frecuente el desarrollo de resistencia a ellos, la cual fue identificada inmediatamente después de la introducción de la penicilina. El mecanismo de resistencia más frecuente en bacilos Gram-negativos a los ß-lactámicos es la producción de ß-lactamasas, y su forma de transmisión es a través de plásmidos y cromosomas. Los bacilos Gram-negativos aeróbicos son los principales responsables de infección nosocomial. Hemos publicado los valores de resistencia de éstos ante ß-lactámicos, haciendo un diagnóstico de la situación a nivel nacional. Además de seguir su evolución durante la década 1988-1998, estudiamos los cambios producidos sobre la evolución de la resistencia, en presencia de inhibidores de ß-lactamasas.
Desde 1988, el Grupo Venezolano de Vigilancia de la Resistencia Bacteriana, cuyos miembros pertenecen a 29 instituciones de salud de siete estados, está a cargo de analizar y publicar resultados de resistencia bacteriana a antimicrobianos de bacterias aisladas de pacientes con infecciones hospitalarias y de la comunidad.
Se usó el método de difusión de disco, de acuerdo a las normas de la NCCLS. Se siguió el programa software WHONET (World Health Organization Net).
Las diferencias de sensibilidad entre el ß-lactámico y el ß-lactámico más el inhibidor de ß-lactamasas son:
1. Piperacilina versus piperacilina/tazobactam: entre el 10 y el 30% para la mayoría de los gérmenes estudiados, excepto para E. coli (45%) y Serratia sp. (60%).
2. Ampicilina versus ampicilina/sulbactam: entre el 10 y el 30%.
3. Cefoperazona versus cefoperazona/sulbactam: entre el 5 y el 25%.
Como era de esperar, la resistencia de bacilos Gram-negativos aeróbicos a betalactámicos en presencia del inhibidor de ß-lactamasas es menor que ante el ß-lactámico solo; además, esta diferencia se hace mayor con el tiempo. Las diferencias entre las dos series durante el primer año (1988) son menores que las encontradas entre las series durante el último año (1998) en muchas de las bacterias estudias.
Estos resultados son relevantes especialmente si recordamos el mecanismo de inductores de betalactamasas que se les ha atribuido.
Palabras-clave: ß-lactamasas, ß-lactámicos, bacilos Gram-negativos, resistencia bacteriana, inhibidores de ß-lactamasas, sulbactam, tazobactam.
Abstract
The ß-lactams antimicrobial were the first to be used, and today they still are the most frequently used. Among the bacteria responsible of high resistance to ß-lactams are gramnegative rods; the most frequent mechanism is the production of ß-lactamase We follow the trends of resistance of Gram-negative rods to ß-lactams alone and with ß-lactamase-inhibitors during the decade 1988-1998.
Since 1988, The Venezuelan Group of Vigilance of the Bacterial Resistance, with 29 health institution in the country; they identify, analyze and publish data on bacteria resistance of isolates from patients with bacterial infection coming from hospitals and the community.
It was used diffusion disk, according NCCLS. The software program WHONET (World Health Organization Net) was used. The difference in sensitivity among ß-lactam and ß-lactam/ß-lactamase-inhibitor are:
1. Piperacillin versus piperacillin/tazobactam: between 10 and 30% of resistance for most isolated, except for E. coli (45%) and Serratia sp. (60%).
2. Ampicillin versus ampicillin/sulbactam: between 10 and 30%.
3. Cefoperazone versus Cefoperazone/sulbactam: between 5 and 25%.
How is expected gramnegative rods resistance to ß-lactams with a betalactamase-inhibitor (ß-L-I), is lower than the ß-lactam alone; furthermore the difference between them, grows higher with time. This results are relevant, since ß-L-I are described as ß-lactamase inductors.
Introducción
Las cefalosporinas de tercera generación y los carbapenemos surgieron como una necesidad ante la presencia de bacilos Gram-negativos productores de ß-lactamasas, tanto cromosómicos como plasmídicos (1), capaces de inactivar los ß-lactámicos en uso.
Las ß-lactamasas aparecieron gradualmente, pero luego aumentaron en forma alarmante (2, 3, 4). Actualmente se describen unas doscientas betalactamasas, entre las cuales se encuentra un gran número de ß-lactamasas de espectro expandido (5), capaces de inactivar los nuevos grupos de ß-lactámicos. Las descritas más recientemente son producidas por un gran número de bacterias, especialmente por Gram-negativos. Algunas muestran actividad ante los carbapenemos; entre ellas se incluyen las metalo-ß-lactamasas (6).
Los mecanismos de resistencia a los ß-lactámicos por parte de los bacilos Gram-negativos son diversos; también lo son sus mecanismos de transmisión (7, 8). Otra característica es la de que estos bacilos con gran frecuencia son responsables de resistencia ante los betalactámicos. Existen resultados de hospitales de Venezuela durante una década donde se evidencia la alta frecuencia de resistencia de éstos ante betalactámicos (9).
Los ß-lactámicos siguen siendo los antimicrobianos más usados; existen varias razones que lo explican: espectro de actividad antimicrobiana, seguridad, eficacia y baja toxicidad. Sin embargo, por ser los primeros antibióticos introducidos en clínica, la resistencia bacteriana ante estos fármacos se ha constituido en un problema por más de 40 años. Hemos revisado con algún detalle los mecanismos de resistencia bacteriana en general (10) y los mecanismos de resistencia ante betalactámicos (11) y, aunque las bacterias tienen una gran creatividad, han desarrollado un limitado número de mecanismos de resistencia.
No obstante, se sabe actualmente que el mecanismo más importante de resistencia antimicrobiana por parte de estos fármacos es la producción de ß-lactamasas y, esencialmente, la ruptura del anillo ß-lactámico. Estas enzimas son producidas por algunas bacterias Gram-positivas y todas las Gram-negativas (2, 3, 4).
Cerca de doscientas ß-lactamasas bioquímicamente diferente han sido identificadas en bacterias Gram-negativas. Estas enzimas, aunque presentan la misma esencial función antes descrita, presentan diferentes propiedades. Así nació la necesidad de agruparlas; Richmond & Sykes, en 1973 (12), clasificó las ß-lactamasas de bacterias Gram-negativas en cinco grupos, sobre la base de su espectro general de actividad (penicilinas o cefalosporinas y otros ß-lactámicos) y su sensibilidad a ser inhibidas por cloxacilina.
Luego aparecieron otras clasificaciones. Recientemente, aplicando estos criterios y basados en la clasificación de Richmond & Sykes, Bush propuso una amplia clasificación, que incluye ß-lactamasas de Gram-negativos y Gram-positivos (2, 3, 4). Esta clasificación de ß-lactamasas tiene cuatro grupos.
Posteriormente (1995) apareció una clasificación, denominada funcional, de Bush-Jacoby-Medeiros (13), la cual se basa en la clasificación original de Bush.
La producción de ß-lactamasas puede ser constitutiva o inducible, al ser afectada por la exposición al antimicrobiano. La constitutiva mantiene un nivel estable y basal, independientemente del estimulo externo. Las ß-lactamasas inducible se produce en gran cantidad después de la exposición a un determinado ß-lactámico inductor. La inducción por ß-lactámicos puede aumentar la producción por ß-lactamasas tanto como mil veces.
Las ß-lactamasas mediadas por plásmidos en Gram-negativos son comúnmente constitutivas.
Las ß-lactamasas cromosómicas de bacterias Gram-negativas pueden ser altamente inducibles en presencia de un particular ß-lactámico. Algunas cefalosporinas de segunda y tercera generación (cefoxitin, moxalactam, cefotaxima) son resistentes a la hidrólisis ante algunas ß-lactamasas, pero han demostrado habilidad para inducir la producción de estas enzimas por algunos microorganismos. De esta manera, existe potencial para el antagonismo entre ß-lactámicos si está presente uno que sea fuerte inductor de ß-lactamasas.
Las ß-lactamasas producidas por microorganismos Gram-negativos se concentran en el espacio periplásmico, y se producen en mucha menor cantidad, mientras que las producidas por las bacterias Gram-positivas se localizan en el exterior y en gran cantidad.
En Gram-negativos la perturbación de la síntesis de peptidoglicanos de la pared lleva a la acumulación de sus precursores en el espacio periplásmico, produciendo éstos la señal para la inducción enzimática (14) en el mismo espacio, donde se acumula la ß-lactamasa producida en menor cantidad que en Gram-positivos.
Es importante señalar que, aunque el mecanismo más importante de resistencia a los ß-lactámicos es la producción de ß-lactamasas, cualquier microorganismo puede desarrollar más de un mecanismo de resistencia a la vez, pudiendo uno de ellos ser el origen más importante de la expresión de resistencia o ser sólo un factor contribuyente que ayuda a la eficacia de la expresión de la misma (15).
El camino más obvio para acabar con la resistencia debida a las betalactamasas es desarrollar un compuesto que las inactive. La gran y frecuente producción de ß-lactamasas como mecanismo de resistencia condujo a la síntesis y purificación de sustancias que inhiben su actividad; éstos son los inhibidores de ß-lactamasas.
Los inhibidores de ß-lactamasas bloquean dichas enzimas, y representan el mecanismo más específico desarrollado para evadir la resistencia a los ß-actámicos. Esto constituyó un importante logro, pues con estos fármacos se recupera la actividad de ß-lactámicos clásicos como ampicilina, amoxicilina, piperacilina, ticarcilina, entre otros. Lamentablemente su eficacia clínica es limitada, pues sólo son capaces de inactivar algunas ß-lactamasas (clase mol. A, grupo 2 de Bush) (16).
La idea de que un ß-lactámico pueda inhibir una ß-lactamasa se describió ya con el uso de la meticilina (17). Posteriormente, algunos otros compuestos han sido estudiados. La penicilina anti-estafilocócica cloxacilina no es hidrolizada por la ß-lactamasas producidas por S. aureus; sin embargo, por su estrecho espectro de inhibición, su uso es restringido. El nuevo concepto de inhibidores específicos de ß-lactamasas comenzó con el reporte del ácido clavulánico en 1976 (18); fue el primero en ser efectivo in vitro e in vivo. Tiene una pobre actividad antibacteriana, y su uso es posible al ser combinado con penicilinas y cefalosporinas inductoras de ß-lactamasas y/o sensibles a ser inactivadas por ellas. Se ha combinado para su uso con amoxicilina y ticarcilina.
Más tarde apareció el sulbactam, molécula semisintética obtenida a partir del ácido 6-amino penicilánico. Tiene pobre actividad antibacteriana y es activo contra N. gonorrhoeae y Acinetobacter sp. Al igual que el ácido clavulánico, tiene utilidad terapéutica al combinarlo con otro ß-lactámico que induzca la producción de ß-lactamasas y/o que sea susceptible a la inactivación por ellas. El sulbactam se combina con ampicilina y cefoperazona. El sulbactam y el ácido clavulánico son excelentes inhibidores de ß-lactamasas pertenecientes a los grupos II, III y IV de Richmond y Sykes y el grupo 2 de Bush. El sulbactam es buen inhibidor de ß-lactamasas clase C, mostrando mayor actividad que el ácido clavulánico (19) ante algunas enzimas, pero este último tiene mayor permeabilidad; de esto resulta que los dos son similares en su efecto de inhibición de las ß-lactamasas.
En cuanto a su mecanismo de inhibición, éste se manifiesta a través de la formación de un complejo estable, irreversible (ß-lactamasa-inhibidor de ß-lactamasa), siendo posteriormente destruida la molécula, tanto la del ácido clavulánico como el sulbactam; por esta razón se denominan inhibidores suicidas (18).
Por otra parte, se ha descrito la unión del sulbactam a PBPs de Gram-negativos, específicamente a la PBP-1a, y esto confiere actividad a bacterias como E. coli, Klebsiella sp., P. vulgaris y Acinetobacter sp.
Existe otra molécula similar a estas en cuanto a su mecanismo de acción, de mas reciente adquisición, el tazobactam. Dicha molécula tiene mayor actividad que sus antecesores ante ß-lactamasas cromosómicas (grupo 1, 2a, 2b, 2b'), las cuales hidrolizan cefalosporinas de tercera generación, por lo que su espectro es mayor (13). Existen evidencias que muestran que se producen reacciones reversibles con cada enzima antes de que ocurra la completa inactivación (13). La piperacilina fue introducida en 1981, mientras que la piperacilina-tazobactam lo fue en 1993; desde entonces ambas han jugado un papel relevante en el tratamiento de las infecciones producidas por gérmenes Gram-negativos (20). El tazobactam tiene características farmacocinéticas y farmacodinámicas que lo hacen más activo como inhibidor de ß-lactamasas que sus predecesores (16). El tazobactam es también un inhibidor de ß-lactamasas pertenecientes al grupo 1 de Bush.
Por otra parte, el clavulanato es un inductor de ß-lactamasas del grupo I, mientras que el sulbactam y el tazobactam parecen tener menor capacidad de inducción (21). Para todas las enzimas clase A, el tazobactam es el más activo. Sin embargo, el clavulanato fue mas efectivo inhibiendo las ß-LEE, derivadas de las TEM.
En relación a los inhibidores de ß-lactamasas, y debido a los argumentos antes planteados debido a su potencial de inducir la producción de ß-lactamasas, es importante conocer el efecto de su uso sobre la evolución de la resistencia a ß-lactámicos por parte de los bacilos Gram-negativos aeróbicos; en este sentido, hemos publicado algunos resultados (22), y vamos a ampliarlos y discutirlos con mayor detalle.
Materiales y Métodos
En Venezuela, en 1988, se creó el Grupo Venezolano de Vigilancia de la Resistencia Bacteriana (GVRB), integrado por investigadores de 29 instituciones de salud o centros de enseñanza universitaria. Esta información se adapta al programa de software WHONET (23), lo que ha permitido la incorporación de Venezuela a la red internacional coordinada por la OMS.
Todos los centros hospitalarios públicos o privados participantes (desde clínicas privadas pequeñas hasta hospitales universitarios con más de 500 camas) registran los datos según un formato ad-hoc. En estos formatos se registra la información sobre cada una de las cepas bacterianas identificadas: datos del paciente, origen de la muestra, nombre de la bacteria, procedencia dentro del hospital y número de milímetros de los halos de inhibición.
Esta información es transcrita a computadoras IBM ubicadas actualmente en la Unidad de Microbiología e Infectología del Hospital Vargas de Caracas.
WHONET (red de la Organización Mundial de la Salud) es un programa software que acepta y analiza los resultados de las pruebas de sensibilidad in vitro a los antimicrobianos. La información es almacenada en archivos que aceptan resultados de diferentes regiones, para ser comparados de acuerdo con las recomendaciones de la OMS. También acepta resultados que van a ser analizados epidemiológicamente. Este programa integra los resultados de los laboratorios participantes a través de todo el mundo (24). La vigilancia de la resistencia a los antimicrobianos es esencial para identificar organismos resistentes y así lograr la adecuada orientación de la terapia antimicrobiana de infecciones y para mejorar los estándares de las pruebas de sensibilidad (24).
En todos los laboratorios participantes se empleó el método de difusión en agar, usando discos de reconocida calidad (Difco, BBL, Oxoid), siguiendo las normas originales de Bauer y Kirby (25). Se siguieron las normas de eficacia publicadas por la NCCLS (Comité Nacional de Estándares para Laboratorios Clínicos) (26).
Se comparan porcentajes de resistencia de bacilos Gram-negativos aeróbicos ante ß-lactámicos (ampicilina, cefoperazona y piperacilina), solos y en presencia de un inhibidor de ß-lactamasas (sulbactam y tazobactam respectivamente).
A. Se analizan y comparan resultados de resistencia ante piperacilina con piperacilina-tazobactam (1996-1998).
B. Se estudia la evolución de la resistencia durante una década, comparando: a) ampicilina con ampicilina-sulbactam, y b) se comparan resultados de resistencia de bacilos Gram-negativos aeróbicos ante cefoperazona con cefoperazona-sulbactam.
Los resultados obtenidos fueron presentados como porcentajes de todos los valores individuales (un mínimo de 50 cepas de cada especie). Para comparar los porcentajes de resistencia se aplicó la evaluación estadística por la formula general del cálculo del error estándar de la diferencia entre dos porcentajes, base del problema, que forma el numerador de la razón conocida como el "valor Z". El criterio de significación estadística fue de 5%.
Resultados
Comparación de la resistencia ante la piperacilina y la piperacilina/tazobactam (1996-1998). (Gráfico 1).
Compararemos las diferencias de la resistencia (D), correspondientes a los años 1996 y 1998, para los diferentes gérmenes estudiados.
E.coli: Se muestra un aumento de la sensibilidad, en presencia del tazobactam, de 45 y 40% en los años 1996 y 1998, respectivamente.
P. mirabilis: Se muestra un aumento de la sensibilidad, en presencia del tazobactam, del 10 y 4% en los años 1996 y 1998, respectivamente.
P. vulgaris: Se muestra un aumento de la sensibilidad, en presencia del tazobactam, del 5 y 10% en los años 1996 y 1998, respectivamente.
E. aerogenes: Se muestra un aumento de la sensibilidad, en presencia del tazobactam, del 30 y 25% en los años 1996 y 1998, respectivamente.
E. cloacae: Se muestra un aumento de la sensibilidad, en presencia del tazobactam, del 12 y 26% en los años 1996 y 1998, respectivamente.
K. pneumoniae: Se muestra un aumento de la sensibilidad, en presencia del tazobactam, del 22 y 28% en los años 1996 y 1998, respectivamente.
P. aeruginosa: Se muestra un aumento de la sensibilidad, en presencia del tazobactam, del 12 y 10% en los años 1996 y 1998, respectivamente.
Acinetobacter sp.: Se muestra un aumento de la sensibilidad, en presencia del tazobactam, del 20 y 25% en los años 1996 y 1998, respectivamente.
S. marcescens: Se muestra un aumento en la sensibilidad de aproximadamente 60% para el año 1996 y de menos de 10% en 1998, en presencia del tazobactam.
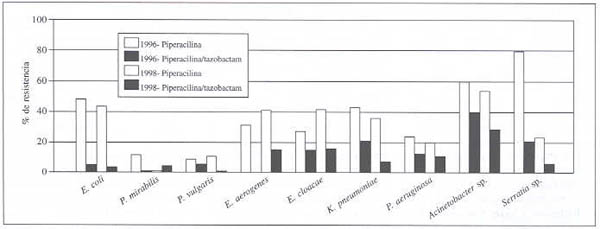
Gráfico 1. Resistencia a piperacilina y piperacilina-tazobactam en bacilos Gram-negativos aeróbicos en Hospitales de Vennezuela. 1996-1998.
Efecto del Sulbactam en la Evolución de la Resistencia de Bacilos Gram-Negativos Aeróbicos ante la Ampicilina y la Cefoperazona (1988-1998).
Comparación entre ampicilina y ampicilina-sulbactam:
Evolución de la resistencia de E. coli a la ampicilina y a la ampicilina-sulbactam (gráfico 2). En este caso hay una diferencia (D) entre las dos series para el año 1988 de 35%, para 1996 es de 40% y para 1998 es 44%. Las diferencias entre las dos series, al compararlas con 1988, para 1996 y 1998 fueron significativas (p<0,05).
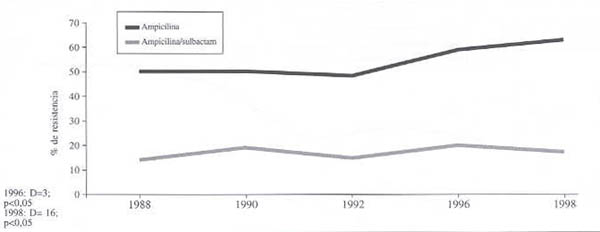
Gráfico 2. Evolución de la resistencia de E. Coli a ampicilina sola en presencia de un inhibidor de ß-lactamasas. 1988-1998
Comparación entre cefoperazona y cefoperazona-sulbactam:
Evolución de la resistencia de E. coli a cefoperazona y cefoperazona-sulbactam (gráfico 3). En este caso la D entre 1988 y 1996 es de 6%; para 1998 la diferencia es mayor y también significativa.
Evolución de la resistencia de K. pneumoniae a cefoperazona y cefoperazona-sulbactam (gráfico 4). En este caso las diferencias son significativas.
Evolución de la resistencia de P. aeruginosa a cefoperazona y cefoperazona-sulbactam (gráfico 5). En este caso la D para 1988 es de 7%, para 1996 de 15% y para 1998 de 13%. Tanto las diferencias 1988-1996 como 1988-1998 son significativas.
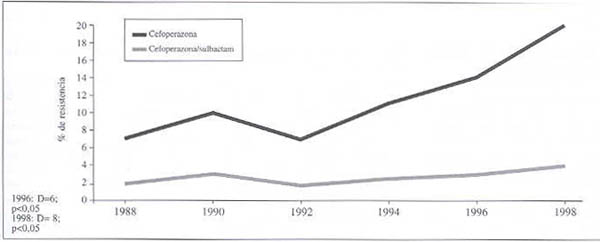
Gráfico 3. Evolución de la resistencia de E. Coli a cefoperazona sola y en presencia de un inhibidor de ß-lactamasas. 1988-1998
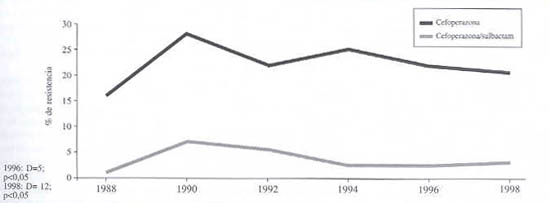
Gráfico 4. Evolución de la resistencia de K. Pneumoniae a cefoperazona sola y en presencia de un inhibidor de ß-lactamasas. 1988-1998
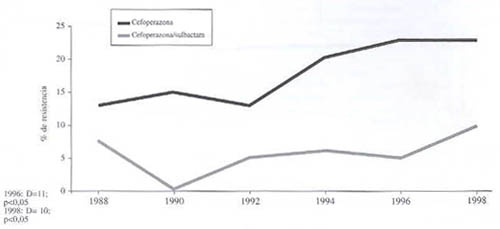
Gráfico 5. Evolución de la resistencia de P. Aeruginosa a cefoperazona sola y en presencia de un inhibidor de ß-lactamasas. 1988-1998
Conclusiones
Es de hacer notar el hecho de que los bacilos Gram-negativos han desarrollado todos los mecanismos de resistencia descritos para los ß-lactámicos ante los carbapenemos, último grupo introducido para evadir las betalactamasas hasta entonces identificadas. Estos mecanismos van desde la más sofisticada bomba de eflujo, cambios en las porinas, cambios en las PBP y la producción de diferentes grupos de ß-lactamasas (carbapenemasas). Estas enzimas son especialmente sintetizadas por diferentes bacterias para inactivar a los carbapenemos (27, 28).
Los I-ß-L probablamente no puedan inactivar las ß-lactamasas antes descritas. Sin embargo, sí pueden recuperar gran parte de la actividad de los I-ß-lactámicos por nosotros analizados.
En algunos de los casos estudiados, a la vez que se observa un aumento de la resistencia mayor ante el ß-lactámico solo (como es de esperar), se observa una diferencia entre ambas tendencias de resistencia, que va en aumento con el tiempo. La pendiente para el ß-lactámico solo es mayor que cuando está en presencia del inhibidor de ß-lactamasas. Las diferencias de las resistencias entre las dos series en el primer año (1988) es menor que las diferencias de las resistencias entre las dos series los últimos años estudiados (1996 y 1998).
Se observa claramente esta diferencia en el caso de E. coli, E. aerogenes y P. aeruginosa ante cefoperazona y cefoperazona-sulbactam; en estos casos se demuestra significancia estadística. En los otros casos se muestra similar tendencia, aunque no haya significancia.
Estos resultados son relevantes, si recordamos el efecto de inductores de lactamasas que se les ha asignado a estos inhibidores (21), por lo que esperábamos disminución de su eficacia con el tiempo. Además, trabajos recientes de Lister y col. (29, 30) establecen diferencias entre el tazobactam y el ácido clavulánico en su efecto de inductores de ß-lactamasas. Por otra parte, al observar nuestros resultados podemos afirmar que los inhibidores de ß-lactamasas siguen siendo de utilidad.
Es relevante observar la alta frecuencia de resistencia de E. coli, K. pneumoniae y Acinetobacter sp.; frecuencias que coinciden con epidemias en el mundo para los dos primeros, y con epidemia nacional para Acinetobacter sp.
Por otra parte, además del aumento en la frecuencia de resistencia en todo el mundo (31) y sus consecuencias directas sobre diferentes aspectos de la salud (32, 33, 34), existe la situación de que no hay antimicrobianos con nuevos mecanismos de acción; los que están en estudio con nuevos mecanismos de acción se encuentran en fases muy tempranas, y tomará algunos años antes de conocer sobre su posible eficacia terapéutica.
Todo lo discutido le da vigencia a los I-ß-l en la terapéutica antimicrobiana actual; y, por otra parte nos señala como mecanismos importantes a ser estudiados para nuevos fármacos aquellos que inhiben mecanismos de resistencia a los antimicrobianos en uso. (Actualmente en estudio se destacan los inhibidores de bombas de eflujo) (35).
Agradecimientos
Al Dr. F. Arias, por su colaboración en la evaluación estadística.
Referencias:
1. Medeiros, A.: Recent Increases in Resistance: Mechanisms and Organisms: Evolution and Dissemination of b-Lactamases Accelerated by Generations of ß-Lactam Antibiotics. Clin Infec Dis; 24: S19-45, 1997.. [ Links ]
2. Bush, K.: Clasification of ß-lactamases: group 2c, 2d,2e, 3, and 4. Antimicrob Agents Chemother; 33: 271-276, 1989. [ Links ]
3. Bush, K.: Classification of ß-lactamases: groups 1, 2a,2b,2e, and 2b¢. Antimicrob Agents Chemother; 33: 264-270, 1989.
4. Bush, K.: Characterization of b-Lactamases. Antimicrob Agents Chemother;33: 252-263, 1989. [ Links ]
5. Nordmann, P.: Trends in b-Lactam Resistance Among Enterobacteriaceae. Clinic Infect Dis. 1998; 27: S100-106. [ Links ]
6. Bush, K.: Metallo-ß-Lactamases: A Class Apart. Clinic Infec Dis; 27: S48-53, 1998. [ Links ]
7. Davies, J.: Inactivation of Antibiotics and the Dissemination of Resistance Genes. Science; 264: 375-382, 1994. [ Links ]
8. Tait, S.: Mobile genetic elements in antibiotic resistance. J. Med Microbiol; 38:157-159, 1993. [ Links ]
9. Martin, N. G.; Carmona, O. y Guzmán, M.: Una década en la evolución de la resistencia ante ß-lactámicos por bacilos gramnegativos aeróbicos en hospitales de Venezuela (1988-1998). Archiv Ven. Farmacol. Clin & Terap; 19:137-147, 2000. [ Links ]
10. Carmona, O. y Silva, H.: Mecanismos de Resistencia a los Antibióticos; Archiv Venez Farmac y Terap; 13: 6-18, 1994. [ Links ]
11. Martin, G.: Resistencia a ß-lactámicos. Mecanismos y Evolución. Aceptado en Archiv Venez Farmac y Terap. 2001. [ Links ]
12. Richmond, H. M. y Sykes, R. B.: The ß-lactamases of gramnegative bacilli and their possible physiologic role. Adv Microbiol Phisiol; 9: 31-88, 1973. [ Links ]
13. Bush, K.; Jacoby, G. A. and Medeiros, A.: A Functional classifications scheme for ß-lactamases and correlation with molecular structure. Antimicrob Agents Chemother; 39: 1211-1233, 1995. [ Links ]
14. Bennet, P. M. and Chopra, I.: Molecular Basis of ß-lactamases Induction in Bacteria. Antimicrob Agent Chemother; 37:153-158, 1993. [ Links ]
15. Bush, K.; Tamaky, S. H. and Sykes, R. B.: Resistance caused by decreased penetration of ß-lactam antibiotic into E cloacae. Antimicrob Agents Chemother; 25: 555-560, 1985. [ Links ]
16. Bush, K. and Macalinstal, C.: Tomos Kinetic Interactions of Tazobactam with ß-lactamases from all majors structural classes. Antimicrob Agents Chemother; 37: 851-8, 1993. [ Links ]
17. Rollinson, G. N. and Batchelor, F. R.: Bacteriological studies on a new penicillin-BRL-1241. Lancet;2: 56-570, 1965. [ Links ]
18. Howorth, T. T. and King, T. J.: Clavulanic acid, a novel b-lactam antibiotic isolated from Streptomices clavurigenus. J Chem Soc Commun; 27: 226-267, 1976. [ Links ]
19. Labia, R.; Barthelemy, M.; Peduzzi, J.; Morand, A.; Tiwari, K. and Kazmierczak, A.: Overcoming Enzymatic Resistance in Bacteria: Impact on Future Therapy. The J. Int. Med. Res; 18:48-57, 1990. [ Links ]
20. Ronald, J. N.: Impact of Changing Pathogens and Antimicrobial Susceptibility Patterns in the Treatment of Serious Infections in Hospitalised Patients The Amer j of Med; 100, 1996. [ Links ]
21. Bolívar, R.; Weaver, S. S. and Bodey, G. P.: Activity of ß-lactamase Inhibitor in Combination with New b-lactam antibiotics against Resistant gramnegative Organisms. Diagn Microbiol Infect Dis;2:255-260, 1984. [ Links ]
22. Martin, G.; Carmona, O. y Guzmán M.: Influencia de Inhibidores de betalactamasas en la evolución de la resistencia de bacilos gramnegativos ante ß-lactámicos en Venezuela (1988-1996). Bol. Infect; 7:54, 1997. [ Links ]
23. Carmona, O.; Guzmán, M.; Silva, H.; Pulido, S. y GVRB: Vigilancia de la Resistencia Bacteriana a los Antimicrobianos en Venezuela. Cuarto Informe. Bol. Soc. Ven. de Microbiol.; 12:11-22, 1992. [ Links ]
24. Stelling, J. G. and O´Brien, T. F.: Surveillance of Antimicrobial Resistance: The WHO Net Program. Clin. Infect. Dis.; 24: S157-S168, 1997.
25. Bauer, A.; Kirby, W.; Sherris, J. C. and Turk, M.: Antibiotic Susceptibility Testing by a Standardized Single Disk Method. Am. J. Clin. Pathol.; 45: 439-496, 1996. [ Links ]
26. National Committee for Clinical Laboratory Standards. Performance Standard for Antimicrobial Disk, Susceptibility Test Approved Standard M2-A4. Villanova, PA. [ Links ]
27. Rasmussen, B. A. and Bush, K.: Carbapenem-Hydrolyzing ß-lactamases. Antimicrob Agent Chemother;41:223-232, 1997. [ Links ]
28. Ito, H.; Arakawa, I. H.; Joshua, S.; Wacharotayankun, R.; Kato, N. and Ohta, M.: Plasmid mediated dissemination of metallo-ß-lactamase gene bla-imp among clinical isolated strains of Serratia marscescen. Antimicrob Agents Chemother; 39:824-829, 1995. [ Links ]
29. Lister, P.; Gardner, V. and Sanders, C.: Clavulanate Induces Expression of the Pseudomonas aeruginosa AmpC Cephalosporinase at Physiologically Relevant Concentrations and Antagonizes the Antibacterial Activity of Ticarcillin. Antimicrob Agents Chemother; 43:882-889, 1999. [ Links ]
30. Lister, P.; Prevan, A. and Sanders, C.: Importance of b-lactamase Inhibitor Pharmacokinetics in the Pharmacodynamics of Inhibitor- Drug Combinations: Studies with Piperacillin-Tazobactam and Piperacillin-Sulbactam. Antimicrob Agents Chemother; 41:721-727, 1997. [ Links ]
31. Cohen, M. L.: Epidemiology of Drug Resistance: Implications for a Post-Antimicrobial Era. Science; 257: 1050-1055, 1992. [ Links ]
32. Moelleering, R.: Antibiotic Resistance: Lessons for the Future. Clin Infec Dis; 27:S135-140, 1998. [ Links ]
33. Holbert, S. D.; Solomon, S. L. and Blake, P. A.: Health and economic impacts of antimicrobial resistance Rev. Infect. Dis. 1987; 9:1065-1078, 1998. [ Links ]
34. Acar, J. F. and Goldstein, F. W.: Consequences of Increasing Resistance to Antimicrobial Agents; 27: 125-134, 1998. [ Links ]
35. Poole, K.: Efflux Mediated Resistance to Fluoroquinolones in Gram-Negative Bacteria. Antmicro Agents Chemother; 44: 2233-2241, 2000 [ Links ]
