










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Facultad de Ciencias Veterinarias
versión impresa ISSN 0258-6576
Rev. Fac. Cienc. Vet. vol.58 no.2 Maracay dic. 2017
Efecto de la Adición de Ácidos Grasos Poliinsaturados sobre el Comportamiento Reproductivo y Productivo en Vacas Mestizas Carora en Los Llanos Centrales de Venezuela
Effect of Polyunsaturated Fatty Acids Supplementation on Reproductive and Productive Behavior in Carora Crossbred Cows under Venezuelan Central Plains conditions
Carlos Domínguez*,1, Ana Z. Ruiz**, Rafael Pérez*, Nelson Martínez***, Livia Pinto*** y Thais Díaz**
*Universidad Experimental Rómulo Gallegos, Programa de producción animal, Área Universitaria El Castrero. San Juan de los Morros 2103, estado Guárico. **Facultad de Ciencias Veterinarias y ***Facultad de Agronomía. Universidad Central de Venezuela, Maracay 2101, estado Aragua, Apartado Postal Nº 4563, Venezuela
Correo-E: cdomig@gmail.com
1 A quien debe dirigirse la correspondencia (To whom correspondence should be addressed)
Resumen
Se evaluaron los efectos de la adición de ácidos grasos poliinsaturados (AGPI) sobre los cambios en el comportamiento reproductivo y productivo en 28 vacas mestizas Carora multíparas, seleccionadas al azar y agrupadas en tres tratamientos (T): TI (n=14): pastos cultivados+4 kg/d de mezcla alimenticia de harina de sorgo (55%), harina de soya (25%) y melaza (25%) más 75 g de mineral. TII (n=7): TI+0,3 kg de grasa sobrepasante (GS), durante los últimos 20 d preparto y 60 d postparto (dpp) y TIII (n=7): TI+0,3 kg de GS, durante los primeros 60 dpp. Se evaluó: peso corporal (PC), condición corporal (CC), ciclicidad ovárica (CO), dimensión de los ovarios (DO), número de folículos por clase folicular: FOLI, II, III, cuerpos lúteos (CL), concentración sanguínea de progesterona (P4), días vacíos (DV), preñez (P), producción total de leche vendible (PTL), y grasa de la leche (% GL). Los datos fueron analizados mediante ANOVA, medidas repetidas en el tiempo, análisis de supervivencia (AS), MANOVA, pruebas de t y de Chi cuadrado (X2). No hubo diferencias (p>0,05) en CC y PC, ni en AS para CO entre tratamientos. A los 15 dpp hubo diferencias para FOLIII entre TII y TI, FOLII entre TII y TIII (p≤0,01) y FOLIII entre TII y TIII (p<0,05). CL fue diferente (p<0,05), a los 15 y 45 dpp. A los 45 dpp FOLII fue diferente (p<0,05) entre TI, TII y TIII (16,0; 18,1 y 7,7, respectivamente), igualmente FOLIII entre T. DO fue diferente (p<0,05) a los 30 dpp en el ovario izquierdo (OVI)del TII. El número de folículos y CL fueron diferentes (p<0,05) OVI durante los primeros 60 dpp. P4 fue superior (p<0,05) en TII, a los 45 dpp (4,12ng/mL±1,60) vs. TI (1,42 ng/mL±0,58) y TIII (0,85 ng/mL±0,46). DV disminuyó (p<0,01) en TII (70,43 ±10,8 d) y TIII (88,4±14,90d) vs. TI (148,7±100,71d). CC al parto influyó sobre PLT (p<0,05). PLT en TIII superó 10,5% y 15,2% a TII y TI, respectivamente (p<0,05). El % de grasa fue diferente (p<0,05) para TII, al día 30 dpp. La suplementación con AGPI tuvo efecto sobre el número de FOLII, III, CL y la dimensión del ovario izquierdo. Asimismo, favoreció la PLT, contrario a lo que ocurrió en el TI. Se demostró el beneficio de los AGPI, como estrategia para mejorar los indicadores reproductivos y productivos.
(Palabras clave: Reproducción; bovinos; producción; grasas inertes; trópico; Carora)
Abstract
The effect of addition of polyunsaturated fatty acids (PUFAs) on reproductive behavior and productive performance was evaluated in 28 crossbred Carora cows randomly grouped in three treatments (T): TI (n=14): Cultivated pastures + 4 kg/d of a mixture of sorghum flour (55%), soybean meal (25%), molasses (25%) plus 75 g of minerals; TII (n=7): TI + 0.3 kg of inert fat (IF) during the last 20 d of gestation and the first 60 d postpartum (dpp); TIII (n=7): TI + 0.3 kg IF during 60 dpp. Body weight (BW), body condition score (BCS), ovarian cyclicity (OC), ovarian dimensions (OD), follicular classes (FOLI, FOLII, FOLIII), corpora lutea (CL), blood progesterone levels (P4), days open (DO), pregnancy (% P), total milk production (TMP), and milk fat (%MF) were assessed. Data were analyzed through ANOVA, survival analysis (SA), MANOVA, t-tests, ANOVA over time, and Chi square. There were no differences (p>0.05) in BCS and BW, and SA for OC between T. On 15 dpp there were differences for FOLIII between TII and TI; FOLII between TII and TIII (p≤0.01); and FOLIII between TII and TIII (p<0.05). CL were different (p<0.05) on 15 and 45 dpp. On day 45 pp, FOLII were different (p<0.05) between TI, TII and TIII (16.0, 18.1 and 7.7, respectively); likewise, for FOL-II among T. OD was different (p <0.05) on 30 dpp in the left ovary (LO) for TII. Follicular classes and CL were different (p <0.05) in LO during the first 60 dpp. P4 was higher (p<0.05) in TII during 45 dpp (4.12 ng/mL±1.60) vs. TI (1.42 ng/mL±0.58) and TIII (0.85 ng/mL±0.46). DO decreased (P<0.01) in TII (70.43±10.8 d); TIII (88.4±14.90d) vs. TI (148.7±100,71d). BCS at calving affected TMP (p<0.05). The TMP in TIII, exceeded in 10.5% and 15.2% that of TII and TI, respectively (p<0.05). %MF was different (p<0.05) in TII, at 30 dpp. The inclusion of PUFAs positively influenced OC for FOLII, FOLIII, CL, and left ovary dimensions, favoring TMP, contrary to TI. Inclusion of PUFASs turned out to be beneficial to improve reproductive and/or productive indicators.
(Key words: Reproduction, bovine; production; inert fats; tropics; Carora)
Recibido: 12/07/17 - Aprobado: 09/01/18
Introducción
La productividad económica en los hatos de bovinos, para carne o leche, depende en gran parte de la eficiencia reproductiva. En el caso de los rebaños lecheros de doble propósito la respuesta es variable, dependiendo de factores de manejo, alimentación y salud. Algunas estrategias para minimizar el efecto de distintos factores sobre el desempeño reproductivo de las vacas se basan en el uso de sustancias nutracéuticas. Dentro de éstas, los ácidos grasos poliinsaturados (AGPI) como el docosahexaenoico, eicosahexaenoico, araquidónico y linoleico conjugado, han sido estudiados en los últimos años [1-2]. Esos AGPI de cadena larga son sintetizados en el organismo a partir del ácido a-linolénico y ácido linoleico a través de varias etapas que implican desaturación y elongación. Los AG a-linolénico y linoleico no pueden ser sintetizados por los animales [1], por lo tanto deben ser aportados en la dieta. La conversión de ácidos a-linolénico y linoleico a AGPI de cadena larga es limitada. Se han estudiado algunos AGPI, principalmente de los tipos omega-3 (n-3) y omega-6 (n-6), observándose que a través de diferentes mecanismos de acción controlan el desarrollo folicular ovárico, la producción de progesterona por el cuerpo lúteo (CL) y la secreción de PGF2a del endometrio, favoreciendo la sobrevivencia embrionaria y la tasa de preñez [3]. En función de esta premisa, se planteó realizar un ensayo cuyo objetivo fue determinar el efecto de la adición de ácidos grasos poliinsaturados (linolénico y linoleico), sobre los cambios en el comportamiento reproductivo y productivo en vacas mestizas Carora bajo condiciones de pastoreo en el eje norte llanero del estado Guárico.
Materiales y Métodos
Animales, Localización, Manejo General y Alimentación de las Vacas
En el ensayo se incluyeron 28 vacas mestizas Carora multíparas, ubicadas en la finca Pirital, municipio Monagas del estado Guárico, bajo condiciones de pastoreo, con pastos cultivados (Echynochloa polystachya, Cynodon plectostachyus y Digitaria decumbens), las cuales fueron seleccionadas al azar de acuerdo a la fecha probable de parto, peso y condición corporal. En general, el manejo llevado a cabo en la finca es de un ordeño manual con becerro, forraje producido en finca, suplementación estratégica e inseminación artificial. La producción de leche registrada previamente oscilaba entre 8-10 L por vaca.
Tratamientos y Diseño del Experimento
Las vacas fueron agrupadas en tres tratamientos (T); TI (n=14): pastoreo con pastos cultivados más 4 kg/d de una mezcla alimenticia (MA) preparada a base harina de sorgo (55%), harina de soya (25%) y melaza (25%); adicionalmente, se le administraron 75 g de una mezcla mineral completa. El tratamiento TII (n=7), recibió la misma alimentación base del TI más 0,3 kg de grasa sobrepasante (Energras®; compuesta de ácidos grasos vegetales de soya, girasol, maíz, coco, palma, algodón, hidróxido de calcio y antioxidante), durante los últimos 20 d antes del parto y los primeros 60 d postparto (dpp) y TIII (n=7), TI más 0,3 kg de grasa sobrepasante (Energras®) solamente durante los primeros 60 dpp. Los requerimientos de las vacas fueron determinados en función de peso corporal de acuerdo a las tablas del National Research Council [4]. El aporte estimado de la energía metabolizable (Mcal EM/kg MS) de la MA y de la grasa sobrepasante fue de 12,78 y 7, respectivamente. El aporte proteico estimado de la MA fue de 602 g de PC/d (22,8% de PC); mientras que el pasto aportó 1,4-1,9 Mcal EM/kg. La mezcla alimenticia y la GS fueron ofrecidas durante el ordeño dependiendo del grupo experimental.
Características y Composición Química de la Grasa Sobrepasante (Energras®)
Grasa total (min) 73%, cenizas (máx) 20%, calcio (máx) 10%. Perfil de ácidos grasos: 17% de ácido linolénico (min), 0,5%, ácido oleico (min) 30%. Relación insaturados:saturados de 1:0,9.
Mediciones Corporales
Semanalmente y desde el momento del parto se evaluó el peso corporal (PC) y la condición corporal (CC) de las vacas, usando para PC (kg) una balanza electrónica marca Roughweight (capacidad 1500 kg); la CC se midió según la metodología de NIRD (National Institute of Research in Dairying) con escala del 1 al 5 [5].
Evaluación de las Estructuras Ováricas
Las estructuras ováricas, se evaluaron desde el día 15 y hasta los 60 dpp, dos veces por semana, mediante palpación transrectal y ultrasonografía. Se realizaron exámenes ecográficos de los ovarios utilizando un escáner en modo B (marca Aloka, modelo SSD 500, Tokio, Japón), equipado con una sonda lineal transrectal de 7,5 MHz. Se registró la presencia de CL y población de folículos ováricos clasificados en clases: FolI (2-5 mm), FolII (6-9 mm), y FolIII (≥10 mm) en cada ovario [6]. Posteriormente, al finalizar la evaluación (60 dpp), las vacas fueron sometidas a un protocolo de sincronización de celos e inseminación artificial a tiempo fijo (IATF). El diagnóstico de gestación se llevó a cabo por palpación transrectal y ultrasonido.
Determinación de Progesterona
Se determinó la concentración de progesterona (P4) en sangre mediante toma de muestras dos veces /semana durante los primeros 60 dpp. Se consideró el reinicio de la actividad ovárica cíclica, cuando dos muestras consecutivas presentaron niveles de P4 en plasma ≥ 1,0 ng/mL [7]. Las determinaciones hormonales se realizaron mediante la metodología de inmunoadsorción unido a enzimas (ELISA) a través del uso de kits comerciales (Progesterone ELISA; DRG Instruments GmbH Germany®). Se utilizó como control de calidad de alta concentración de P4 suero de vacas gestantes y como control bajo, suero de becerras. Los coeficientes de variación (CV) intra e inter-ensayo, para valores altos de P4, variaron entre 7,5-8,3 % y 14,3%, respectivamente; mientras que para valores bajos de P4 fueron entre 7,4-8,3 % y 13,6%, respectivamente.
Días vacíos y Porcentaje de Preñez
Los días vacíos (DV) fueron estimados cuantificando los días transcurridos desde el parto hasta la determinación de preñez por palpación rectal y ultrasonido; igualmente, la cuantificación porcentual de preñez (PP) por tratamiento.
Producción Total de Leche y Porcentaje de Grasa
Se registró diariamente la producción total de leche vendible (PTL) en cada ordeño y consumida por el becerro (LCB), estimada por el método de doble pesaje antes y después del amamantamiento restringido, dos veces por semana hasta los 60 dpp. Para la determinación del porcentaje de grasa de la leche, se tomó una muestra individual cada 15 d de aproximadamente 300 mL de leche, previo mezclado de la leche total producida en el ordeño. Las muestras, debidamente identificadas, fueron transportadas bajo refrigeración hasta el laboratorio de la Cátedra de Industria de la Leche y de la Carne, de la Facultad de Ciencias Veterinarias de la UCV. El porcentaje de grasa se determinó mediante el método de Babcock.
Análisis Estadístico
Las variables zoométricas, CC y PC, fueron analizadas a través de análisis de varianza con medidas repetidas en el tiempo (PASW Statistic, versión 18), considerando las variables zoométricas en el tiempo, intra-sujetos y tratamiento como variable inter-sujeto y su interacción. Además de una prueba de t de comparación de medias para muestras independientes.
Los niveles de progesterona fueron analizados a través de un ANOVA y la CO durante el postparto temprano fue evaluada a través de un análisis de supervivencia de Kaplan Meier, utilizando como variable categórica de CO el valor 1, el cual corresponde al nivel de P4≥1,0 ng/mL, correspondiente a la vaca ciclando y el valor 0, a aquellos niveles de P4<1ng/mL, correspondiente a la vaca que no estaba ciclando, comparando con la presencia o no de un cuerpo lúteo funcional identificado por ultrasonido. Se empleó el estadístico Log Rank (Mantel-Cox) para evidenciar diferencias estadísticamente significativas entre las curvas de sobrevivencia.
Los valores totales obtenidos para FOLI, FOLII, FOLIII y CL fueron tabulados y analizados estadísticamente mediante un modelo lineal general (MANOVA), de medidas repetidas en el tiempo (PASW Statistic, versión 18). Las medias fueron estimadas por mínimos cuadrados y su comparación se realizó a través de la prueba de mínima diferencia significativa (DMS). El modelo incluyó efecto del tratamiento, días postparto de la evaluación y la interacción respectiva. Igualmente, la dimensión de los ovarios fue analizada durante los primeros 60 dpp a través de las pruebas de comparación de medias de t. Para comparar el número de estructuras ováricas (FOLI, II, III y CL) encontrado, entre ambos ovarios, durante los primeros 60 dpp, se utilizó un modelo lineal general (ANOVA) con medidas repetidas en el tiempo. Finalmente, se determinó los DV utilizando el análisis de supervivencia y el porcentaje de preñez por Chi cuadrado (c2).
El análisis de PLT se realizó a través de un modelo lineal general multivariado, en el cual se consideró PLT y CC al parto (CCP) y a los 15, 30, 45 y 60 d y a través de pruebas de comparación de medias entre los tratamientos, durante los primeros 60 dpp. El porcentaje de grasa en leche (corregida al 4%) se analizó y comparó entre tratamientos a través de una prueba de t para diferencia de medias en el lapso del experimento.
Resultados y Discusión
Las vacas en el experimento pastorearon durante los meses de lluvia y en época de transición lluvia-sequía en potreros de pastos introducidos, los cuales mostraron un contenido de proteína cruda dentro de valores adecuados (6,2-8,3%) durante gran parte del ensayo, con tendencias a disminuir al comienzo de la época seca. Durante esta época las vacas se mantuvieron en potreros, con adición de soca de maíz (3,8% PC) y pastos picados de guinea (Panicum maximun) con 6,9 % de PC. La MA aportó valores adecuados de proteína (11,9% PC) y minerales (0,18% P y 1,17% Ca). Con la diferencia del aporte adicional de grasa sobrepasante a los grupos experimentales TII y TIII.
Para estimar la cantidad de grasa añadida a la dieta, basada en el nivel de fibra y la fuente de grasa insaturada, se hizo de acuerdo a la ecuación formulada por Jenkins [8], en la cual el % de grasa en la dieta (MS) = (4×FDN)/AGPI; donde FDN es el % de fibra detergente neutra del forraje y AGPI la proporción de ácidos grasos poliinsaturados del total de ácidos grasos de la grasa sobrepasante empleada (C18:1 + C18:2 + C18:3). En nuestro caso, el promedio de FDN del pool de forrajes empleado estuvo alrededor de 82,3%. La proporción de AGPI en la grasa empleada fue de 64,3%. Resultando en un aporte neto de 5% de AGPI en la dieta, a partir de la mezcla alimenticia. Es decir, un aporte estimado de 1,87% de AGPI en el total de la dieta consumido por una vaca promedio al día (TII y TIII).
La adición de grasa sobrepasante incorpora mayor cantidad de AGPI en la dieta, lo que genera no sólo un aporte energético, sino también, efectos no energéticos beneficiosos relacionados con el impacto que tienen estos AG sobre el metabolismo, la respuesta hormonal e inmunológica. El efecto energético está relacionado con la mayor cantidad de energía que aportan los lípidos, lo que contribuye a disminuir el balance energético negativo (BEN) durante el periodo postparto temprano, una mayor producción de hormona luteinizante (LH) y de la hormona folículo estimulante (FSH) por la hipófisis, generando un mayor crecimiento y desarrollo folicular y favoreciendo a la ovulación [9].
Los efectos no energéticos están asociados al tipo de AG presentes en la grasa suministrada, con el incremento de los niveles de colesterol (particularmente la fracción HDL o colesterol bueno), efectos directos a nivel ovárico y uterino, incrementando los niveles de P4 y la modulación en la producción de prostaglandinas (en particular PGF2a), respectivamente, y sobre las hormonas y factores de crecimiento involucrados con la actividad reproductiva y productiva (insulina, IGF-I, entre otros). La mayoría de estos efectos no energéticos se ven favorecidos cuando se utilizan AGPI (w-6 y w-3). Los AGPI benefician el sistema inmune y el sistema reproductivo, promoviendo el reinicio temprano de la actividad ovárica postparto y la fertilidad [10]. Staples y Thatcher [11] señalaron que la reproducción del ganado está más influenciada por el tipo de lípidos consumidos, que por el consumo total de grasa, algunos AG insaturados pasan intactos por el rumen y se absorben en el intestino delgado, pudiendo mejorar la eficiencia reproductiva. Los mecanismos potenciales pueden incluir su efecto sobre el desarrollo folicular, producción de P4, supresión de las señales luteolíticas en torno al reconocimiento materno de la preñez y mejoramiento de la calidad del ovocito y del embrión.
La proporción de AGPI en la grasa empleada en nuestro experimento fue de 64,3%, de lo cual 23% correspondió a ácido linoleico (C18:2), es decir con predominancia de w-6.
Mediciones Zoométricas
Peso y condición corporal
Con respecto a la CC, no se observaron diferencias entre las medias para los tratamientos analizados. Igualmente, el análisis de varianza por medidas repetidas en el tiempo no detectó diferencias entre los tratamientos y sus interacciones (T x CC) a los 0, 7, 15, 22, 30, 37, 45, 52 y 60 dpp (Figura 1). Sin embargo, se observó que las vacas de los tratamientos TI y TII (sin suplementación con AGPI y suplementadas pre y postparto con AGPI, respectivamente) tendieron a registrar mayores pérdidas de CC a partir del día 0 y durante los primeros 15 dpp, que las suplementadas con AGPI (TIII) solamente durante el postparto, alcanzando el punto nadir para ese momento. Estas pérdidas en CC sitúan a las vacas en condiciones por debajo de 2,5 puntos en la escala de 0-5 [5], considerado como punto de inflexión, en el cual los animales tienden a entrar en un BEN durante el período de transición postparto. Su recuperación depende de la energía, en calidad y cantidad, consumida en la dieta y los requerimientos exigidos por los procesos fisiológicos del momento. A partir del día 22 pp, se revierten las tendencias en la pérdida de la CC. Mientras que las vacas en el TIII, a partir del día 30 pp mostraron una tendencia acelerada de recuperación de la CC hasta el día 45 pp, situándose por encima de las vacas TI y TII (Figura 1).

Con respecto a los resultados de PC, no se detectaron diferencias estadísticas entre las medias mediante la prueba de t. Así mismo, el análisis de varianza para medidas repetidas en el tiempo no registró diferencias entre tratamientos y sus interacciones. Sin embargo, es notable la tendencia observada entre los tratamientos TII y TIII vs. TI. Este último mostró mayores pérdidas de PC desde el día 0 hasta el día 22 pp. Así, en la Figura 2 se observa que estas pérdidas en el PC estuvieron acompañadas con pérdidas de la CC (Figura1). Mientras que en las vacas del TII, suplementadas pre- y postparto con AGPI, comenzaron a recuperar de manera progresiva el PC a partir del día 15 postparto, lo cual se reflejó en incrementos de la CC. Estos cambios en las medidas zoométricas (PC y CC) pudieran influir de manera diferencial en el desarrollo de los eventos reproductivos a nivel ovárico, uterino y en la fertilidad temprana y eventualmente en los procesos productivos de las vacas.
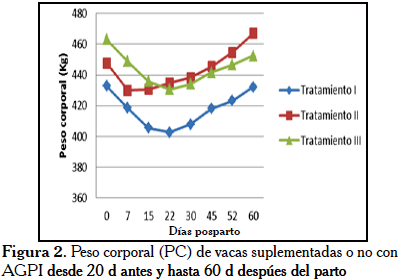
Se ha especulado que los lípidos de la dieta actúan como agentes distribuidores de nutrientes que pueden cambiar el uso de la energía de un proceso metabólico a otro, incrementando con ello el potencial de las vacas para almacenar grasa corporal y ganar o mantener su CC [12]. Los mismos autores señalaron que la utilización de grasas, como una estrategia de suplementación energética en vacas de carne, durante el periodo postparto, permite conseguir el peso y la CC adecuados a los 90 d de este periodo, cuando son comparados con animales que no tuvieron la inclusión de grasa como fuente energética. Sin embargo, la suplementación de ácidos grasos específicos en la dieta puede no compensar, de forma efectiva, el gasto energético del animal durante la lactación temprana, lo que se ve representado en los cambios que ocurren en el tejido adiposo, pues las exigencias de la lactancia y la bioquímica del tejido adiposo parece estar alterada en favor de la distribución de nutrientes hacia el tejido mamario para la síntesis de leche, a expensas de las reservas grasas de la CC [12]. Dietas que contienen un suplemento rico en ácido oleico permiten que la vaca presente un incremento en la producción de leche, mientras que si el suplemento es rico en ácido linoleico, la vaca va a mantener la CC durante el postparto. Una dieta rica en ácido linoleico, en vacas que al momento del parto presentaron una CC pobre, incrementa las reservas palpables de tejido adiposo durante la lactación temprana, aunque esta estrategia de manejo no parece alterar la partición de nutrientes. Se ha reportado que la suplementación de lípidos no influye positivamente en el peso corporal de los animales durante la lactación temprana. En contraste, estos mismos investigadores [12] mostraron un incremento en la CC, debido al aumento de las reservas de tejido adiposo en vacas que consumieron un suplemento graso.
En este experimento a pesar que no hubo diferencias significativas entre los tratamientos para la CC y PC (Figuras 1 y 2), las vacas de los grupos TI y TII, tendieron a perder mayor CC durante los primeros 15 dpp. Pero también, estas últimas (TII) a partir del día 30 pp recuperaron CC, superando a TI y TIII. Los resultados concuerdan con los trabajos realizados por otros autores [13], utilizando grasa sobrepasante en vacas cruzadas en el trópico, las cuales no presentaron diferencias en la CC. Recientemente, Roodbari et al. [14] demostraron que la suplementación con ácido linoleico conjugado durante el periodo de transición (21 d antes del parto y 21 d después del mismo) redujo las pérdidas de CC al compararse con las vacas no suplementadas con AGPI. Por otro lado, Salehi et al. [15] en un ensayo sobre suplementación preparto de vacas alimentadas con canola (alto en ácido oleico), girasol (alto en ácido linoleico) y control (sin ácidos grasos), igualmente, no encontraron diferencias en CC durante los primeros 30 dpp. Asimismo, Dirandeh et al. [16] suplementando en el postparto con dos tipos de AGPI (aceite de linasa y soya) y grasa saturada, no encontraron diferencias en CC durante los primeros 40 dpp. Palma et al. [17] no encontraron diferencias entre los tratamientos control (sin suplementación con ácidos grasos insaturados) y suplementadas con aceite de soya y/o suplementadas con sales de calcio y ácidos grasos insaturados durante el preparto (35 d) y postparto (85 d), observándose que la CC se mantuvo en promedio en 2,6; 2,5 y 2,5, respectivamente, similar a la CC encontrada en vacas de nuestro experimento entre los 37 y 60 dpp. Así mismo, Palma et al. [17] no encontraron impactos negativos de la suplementación con ácidos grasos de aceite de canola sobre el peso vivo y la CC, sugiriendo el mantenimiento de un balance energético positivo y movilización limitada de la grasa de depósito del tejido adiposo. Es posible que el balance de energía de las fuentes grasas sea menos intenso durante el periodo de transición, en comparación con la dieta control. Este efecto se asocia con la reducción de la partición de nutrientes para la grasa de la leche y la movilización de la grasa corporal. Después del parto, el balance energético de estas vacas vuelve a ser positivo, más temprano (Figura 1). Lo que evidencia que los suplementos de grasa durante la lactancia temprana pueden reducir la pérdida de peso corporal de las vacas (Figura 2).
Ciclicidad ovárica
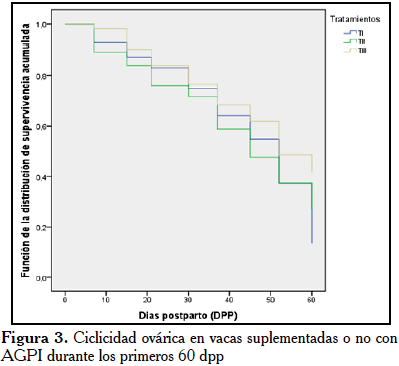
Los resultados del análisis de sobrevivencia de Kaplan Meier para la ciclicidad ovárica, incluyeron los eventos de los animales que ciclaron durante el estudio. Se observó que no hubo diferencia significativa entre las curvas para ciclicidad ovárica, de acuerdo al estadístico Log Rank (p=0,286). Sin embargo, se pudo observar que una mayor proporción de vacas suplementadas con AGPI pre- y postparto (TII), tendieron a mostrar una mayor proporción de vacas con CO durante los primeros 45 dpp, al compararse con vacas no suplementadas (TI) y suplementadas solo durante el postparto (TIII). Evidenciándose que un 50% de vacas TII presentó CO a los 45 dpp, en comparación a 40% de vacas en TI y 35% de las vacas en TIII, las cuales tendieron a ser el grupo con menor CO durante el período (Figura 3).
Debido a la importancia de los ácidos grasos específicos y su rol en los procesos reproductivos, el comportamiento reproductivo puede ser mejorado incorporando en la dieta ácidos grasos de cadena larga específicos, más que alimentación con grasa como un todo [18]. Los mecanismos involucrados en el mejoramiento del comportamiento reproductivo dependen de la composición del ácido graso en el suplemento y la permanencia del mismo en la sangre. Por lo tanto, es determinante aumentar la cantidad de estos ácidos grasos que escapan de la biodegradación ruminal, para mejorar su absorción en el intestino delgado.
En cuanto a la evaluación de la ciclicidad ovárica usando una curva de sobrevivienvia (Figura 3), solo mostró una tendencia positiva en el TII, posiblemente esto pudiera obedecer a una mayor actividad endocrina en este grupo de vacas suplementadas. La adición de grasa a la dieta ha mostrado un consistente incremento del colesterol a nivel plasmático, fluido folicular y en cuerpo lúteo [18], ya que constituye el precursor o sustrato en la síntesis de progesterona por las células foliculares; igualmente, las concentraciones de lipoproteínas de alta y baja densidad mueven el colesterol hacia el tejido para la esteroidogénesis.
Población folicular y cuerpo lúteo
El análisis de correlación de Spearman (no paramétricas) se realizó entre los tratamientos y las variables de población folicular (FOLI, II y III) y CL a los 15, 30, 45, y 60 dpp. Solo resultando significativo y negativo (p<0,05) la correlación entre tratamiento y FOLII, a los 60 dpp. El análisis de varianza multivariada (MANOVA) incluyó como variables dependientes las diferentes poblaciones de estructuras ováricas representadas: número de folículos I, II, III y CL para los días 15, 30, 45 y 60 dpp. Como variable independiente se incluyó el tratamiento (T) (I, II y III), y la variable CC al parto (CCP) como covariable.
En el Cuadro 1, se muestran las diferencias comparativas para la población folicular y CL obtenidas.
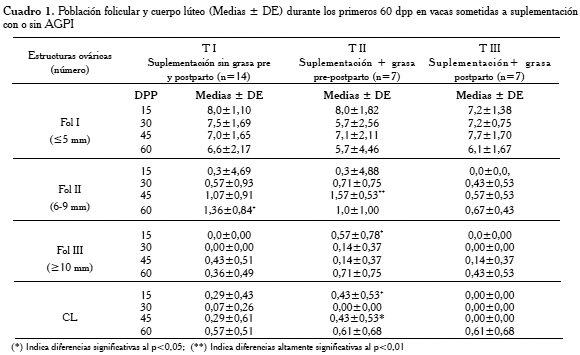
La prueba t de diferencias de medias, para las mismas variables, indicó diferencias significativas (p≤0,01) para FOLIII a los 15 dpp entre TI y TII, siendo los valores de las medias 0,00 vs. 0,57, respectivamente. Estas diferencias se mantuvieron hasta el día 60 pp. Se observó igualmente diferencias significativas (p≤0,01) entre TII y TIII para FolII a los 45 dpp, con medias de 1,57 vs. 0,57, respectivamente y una diferencia (p<0,05) entre TII y TIII para FolIII a los 15 dpp (0,57 vs. 0,0), respectivamente. Adicionalmente, se detectó diferencia significativa (p≤0,01) en la población de CL a los 15 y 45 dpp (0,43 vs. 0,0) entre TII y TIII, respectivamente. Un análisis de varianza no paramétrico (Kruskal-Wallis) determinó diferencias (p<0,05) en los promedios totales de FOLII entre los tratamientos I, II y III (16,0; 18,1 y 7,7, respectivamente); especialmente durante los 45 dpp cuando la diferencia fue superior (14,2; 19,7 y 9,71, respectivamente). De igual manera, se pudo detectar diferencia significativa (p<0,01) en la población FOLIII total, entre los tratamientos I, II y III (13,19 y 13, respectivamente). La misma se puede visualizar el número acumulado de la población de FOLIII, durante los primeros 30 dpp, la cual confirmó que fue superior en las vacas del TII, como se muestra en la Figura 4.
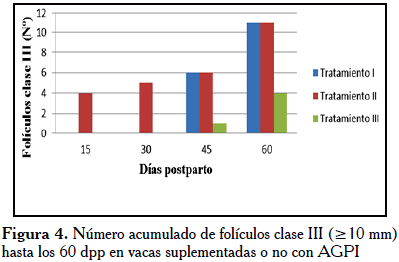
Con respecto a la población folicular y CL, pudimos observar un efecto significativo (p<0,05) de la suplementación con AGPI pre- y postparto (TII) sobre la población folicular clase II (6-9 mm) a los 45 dpp y clase III (≥10 mm) a los 15dpp; igualmente, para CL a los 15 y 45 dpp, respetivamente, al compararse con los otros tratamientos (TI y TIII) (Cuadro 1). Nuestros resultados evidenciaron que el número acumulado de folículos clase III, fue mayor en el TII que en TI y TIII, desde los primeros dpp, como se observa en la Figura 4. Estas tendencias coinciden con los reportado por Hernández y Díaz [1] en vacas Brahman puras de primera lactancia, suplementadas con AGPI en el pre- y postparto.
Evans et al. [19] señalaron que un mayor número de folículos clase III indicaría mayores posibilidades de tener mayor número de CL, mayor número de células luteales produciendo progesterona, lo cual parcialmente suprimiría la producción de prostaglandina por el útero al momento de la concepción, impidiendo la regresión del cuerpo lúteo, en consecuencia mejorando la fertilidad. Al respecto, Hernández y Díaz [1] concluyeron que un mayor número de folículos clase III podría ser un indicador de una mayor probabilidad de tener folículos preovulatorios, por lo que habría mayor oportunidad de ovulación en vacas que consumen grasa durante el pre y postparto.
Lucy et al. [20] utilizaron ácidos grasos de cadena larga protegidos con sales de calcio al inicio del postparto, encontrando un incremento en el número de folículos de tamaño medio (6-9 mm) durante los primeros 25 dpp y folículos mayores >15 mm en un ciclo estral sincronizado; igualmente, encontraron que el diámetro del folículo mayor fue superior en estas vacas, con esta dieta, al compararse con las vacas control (alimentadas con maíz). Robinson et al. [21] reportaron un incremento en el número de folículos de tamaño mediano (5-10 mm) en vacas lecheras en lactación temprana suplementadas con AGPI (linoleico y linolénico), específicamente determinaron en el primer folículo dominante, mayores concentraciones del IGF-I al momento del estro y de colesterol en vacas suplementadas con ácido linoleico (LA, n-6). Staples y Thatcher [11] encontraron que el diámetro del folículo dominante superó en 3,2 mm el diámetro del folículo dominante en vacas del grupo control, el cual representó un 23% de incremento en el diámetro del folículo en las vacas suplementadas con grasa. Santos et al. [18] señalaron que existe una relación entre el incremento del número y tamaño de los folículos más grandes, producto de la suplementación con ácidos grasos y la reducción en el intervalo parto y primera ovulación postparto.
Por otro lado, Badiei et al. [22] experimentaron con diferente duración de la suplementación con grasa (n-3) durante el período peri-parto, sobre el rendimiento lácteo y reproducción de vacas lecheras. En este caso, los animales fueron suplementados con ácidos grasos de cadena larga (AGCL; n-3) en diferentes momentos del periodo seco (últimas 8, 5 y 3 sem) vs. no suplementadas con AGCL en los mismos períodos. Encontrándose que las vacas suplementadas con AGCL tenían folículos ovulatorios de mayor diámetro en comparación con aquellas alimentadas con la dieta control.
Dimensión ovárica
Al comparar las dimensiones de los ovarios (derecho e izquierdo) durante los primeros 60 dpp, se encontró que los principales cambios de los ovarios fueron observados en las vacas suplementadas con AGPI en pre- y postparto (TII). Esto se observó en ambos ovarios durante el periodo de evaluación; sin embargo, las diferencias fueron detectadas en los primeros 30 dpp en el ovario izquierdo, ya que a los 15 dpp hubo una tendencia (p=0,08) a favor del TII y a los 30 dpp la diferencia fue estadísticamente significativa (p≤0,05), sugiriendo una mayor actividad de crecimiento de estructuras ováricas durante ese período. A los 45 dpp, el ovario derecho mostró una tendencia de mayor dimensión, pero no resultó significativamente diferente. En la Figura 5, se observan las diferencias en la dimensión de los ovarios izquierdo y derecho según el tipo de suplementación empleada.

El análisis de varianza (GLM) con medidas repetidas en el tiempo de un factor, sobre las estructuras ováricas (FOLI, II, III y CL) en ambos ovarios (izquierdo y derecho) durante los primeros 60 dpp encontró diferencias significativas (p<0,05) en el ovario izquierdo, y una tendencia (p=0,10) en el ovario derecho. En la estimación de los parámetros (Prueba t), considerando dpp (15, 30, 45 y 60) como variable dependiente y tratamiento (T) como variable independiente, el número promedio de estructuras en el ovario izquierdo fue significativamente superior (p<0,05) en el TII, al comparar con el número promedio en el TI y TIII a los 15 y 60 dpp, y en el ovario derecho a los 45 dpp. La media global igualmente fue significativamente (p<0,01) superior en el ovario izquierdo, al compararla con la media en el ovario derecho (Cuadro 2).
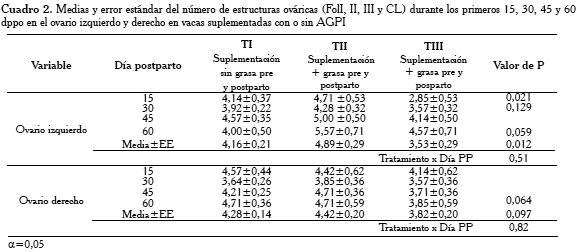
Los cambios en la población folicular (FOLII y III) y del número de CL encontrados, reflejaron una mayor actividad ovárica en las vacas TII que en las vacas TI y TIII, influyendo en una mayor dimensión promedio del ovario izquierdo en el TII, desde el día 15 al 30 pp (p≤0,05). Estas diferencias estadísticas en los promedios de la dimensión ovárica fueron validadas estadísticamente en un modelo lineal general con medidas repetidas en el tiempo, resultando igualmente significativa (p≤0,05), para el ovario izquierdo. Una posible razón de estas diferencias fue la presencia de mayor número de estructuras ováricas de la serie folicular (FOLI, II y III) y CL, cuyos números fueron superiores en promedio durante todo el experimento (15, 30, 45 y 60 dpp), siendo las diferencias significativas (p<0,012) más notables durante los días 15 y 60 pp (Cuadro 2).
Un estudio de Mossa et al. [23] reportó alta correlación (p<0,001) entre el área o dimensión del ovario y el contaje de folículos antrales (CFA), este contaje incrementó en la medida que aumentaba la dimensión del ovario. Es decir, que las vacas con un CFA bajo tienen ovarios más pequeños que aquellas con un número mayor. Previamente, otros estudios encontraron que las vacas que fueron alimentadas con aceite de pescado aumentaron el número de folículos medianos (5-10 mm de diámetro) [24]; sin embargo, hay incoherencia entre los hallazgos sobre los efectos de los AGPI en la dieta sobre la foliculogénesis, y esto puede atribuirse a las diferencias entre los estudios en las cantidades de suplementos, la etapa de lactancia y el estado metabólico de las vacas. Mossa et al. [23] indicaron que las vacas lecheras con un bajo número de folículos ováricos (≤15) tienen menor rendimiento reproductivo en comparación con las vacas con mayor número de folículos que crecen durante las ondas foliculares, lo cual fue confirmado por Evans et al. [19], demostrando que las vacas con un número relativamente alto de folículos antrales, tenían tasas de preñez más altas, intervalos parto-concepción más cortos y menor número de servicios de IA/concepción, en comparación con vacas con menor número de folículos antrales.
Niveles de progesterona
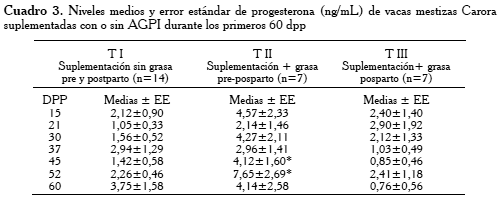
Los niveles medios y error estándar de progesterona plasmática durante los días 15, 21, 30, 37, 45, 52 y 60 pp se observan en el Cuadro 3. El análisis de varianza encontró valores de P4 con diferencias significativas (p<0,05) entre TII durante los días 45 y 52 (4,12±1,60 ng/mL y 7,65±2,69 ng/mL, respectivamente) vs. TI (1,42 ±0,58 ng/mL y 2,26±0,46 ng/mL, respectivamente) y TIII (0,85±0,46 ng/mL y 2,41±1,18 ng/mL, respectivamente).
La literatura en el contexto de la progesterona y su relación con los AGPI es inconsistente. En un estudio se observó un incremento de la producción de P4 (in vitro) cuando se cultivaron células de la granulosa de cabras en un medio enriquecido con ácido linoleico, pero no con ácido a-linolénico [25]. Igualmente, estudios in vitro indicaron que el metabolismo de la progesterona puede ser inhibido por concentraciones altas de ácido a-linolénico (300 µM) en el medio de cultivo [26]. Si esto ocurre in vivo, tal vez se relaciona con que ciertos AGPI pueden incrementar la concentración sérica de insulina y ésta a su vez, reduce la expresión hepática de algunas enzimas del complejo citocromo P450 que catabolizan la progesterona [27].
Por el contrario, Dirandeh et al. [16] no encontraron diferencias en la concentración plasmática de progesterona entre dietas suplementadas al parto con ácido linoleico (aceite de soya) y ácido a-linolénico (aceite de linasa), pero si con respecto al control, durante los primeros 20 dpp.
En nuestro experimento, las concentraciones de P4 (ng/mL) fueron superiores en el tratamiento II desde el día 15 hasta los 60 dpp, comparadas con las vacas de los TI y TIII (Figura 3 y Cuadro 3), siendo las diferencias significativas (p<0,05) durante los días 45 y 52 pp. Estos niveles promedios de concentración plasmática de P4 (4,12±1,60 ng/mL y 7,65±2,69 ng/mL para los días 45 y 52 pp, respectivamente), coinciden con el mayor promedio en el número de CL (Cuadro 1) en las vacas TII a los 45 dpp, con respecto a los grupos TI y TIII.
Los valores de P4 observados en TII a los 45 dpp, se asemejan a los niveles encontrados en vacas suplementadas con AGPI protegidos, reportado por Cardona et al. [28] a los 30 d del período de suplementación y en el momento de la transferencia de embriones, siendo los niveles de P4 de 5,41±1,84 ng/mL, superior a 3,68±1,17 ng/mL (control) y 3,65 ng/mL (tratamiento alimentado con maíz).
Días vacíos y preñez
Se observó que el promedio de DV fue menor en los grupos suplementados con AGPI, siendo menor en las vacas del TII (70,43±10,8 d), seguido de TIII (88,4±14,90 d) y TI (148,7±100,71 d). Se pudo observar que el ANOVA, y la prueba estadística (Log Rank) de la prueba de sobrevivencia de Kaplan-Meier para DV, basado en las proporciones (Chi-cuadrado) fueron diferentes estadísticamente (p<0,01). Por lo que se pudo demostrar el impacto de la suplementación pre y postparto con AGPI, sobre la respuesta reproductiva (Figura 6).
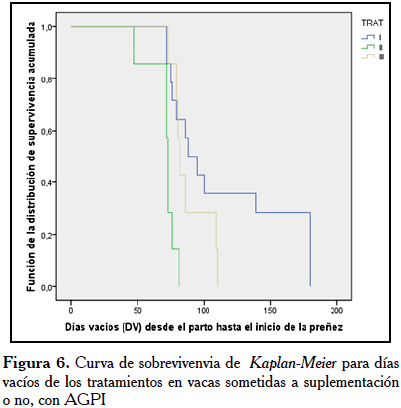
El efecto de la suplementación con AGPI influyó en menor número de DV en las vacas del grupo TII, en comparación con las vacas de los grupos TIII y TI, como se observa en la Figura 5, demostrándose el impacto de la suplementación pre- y postparto con AGPI, sobre la fertilidad y la respuesta reproductiva. Hubo 100% de preñez en las vacas suplementadas con AGPI pre- y postparto (TII) y suplementadas postparto (TIII), contra 74,1% de las vacas control, resaltando lo exitoso de la suplementación durante la aplicación de este protocolo, basado en una alimentación estratégica previa con AGPI.
Es posible que la recuperación más rápida en el peso, la CC y la mejora de la funcionalidad ovárica desde el pre-parto, influyó determinantemente en mejorar el proceso de involución uterina, la foliculogénesis, la calidad y el número de los folículos pre-ovulatorios clase III [19] y CL en las vacas TII, lo cual mostró una fertilidad más temprana (menos DV), como también lo han señalado otros autores [18]. Recientemente, Branko et al. [29] comprobaron que el incremento en los AGPI en el suero sanguíneo, puede causar un aumento de los mismos en el fluido folicular, el cual podría influir en la dinámica folicular y en la fertilidad. Los mismos autores reportaron que alrededor de un tercio del fluido folicular correspondió a ácido linoleico (C18:2; n-6) en las vacas suplementadas con AGPI enriquecidas con ácido linoleico, lo cual indica la posible existencia de mecanismos selectivos en el folículo que regulan la concentración de C18:2; n-6 en el fluido folicular, siendo particularmente importante para el desarrollo de la competencia del ovocito.
Juchem et al. [30] evaluaron el efecto de la suplementación de vacas antes y después del parto con sales de calcio de AG de cadena larga (Ca-AGCL) derivados del aceite de palma o una mezcla de ácidos linoleico y trans-octadecenoico. Las vacas alimentadas con AG insaturados tuvieron mayores tasas de preñez comparadas con las vacas suplementadas con aceite de palma a los 27 (37,9 vs. 28,6%) y 41 d (35,5 vs. 25,8%) después de la inseminación artificial, respectivamente.
Producción de leche total
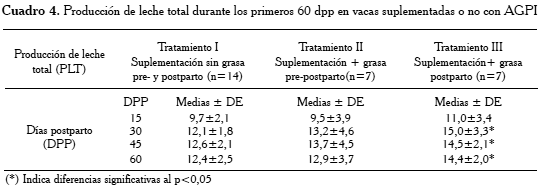
Los resultados a partir de un modelo lineal general multivariado para PLT durante el período evaluado (15, 30, 45 y 60 dpp), demostraron que la CCP influyó sobre PLT a los 30 y a los 60 dpp (p<0,05). Siendo el grupo de vacas correspondiente a TIII las que alcanzaron un mayor volumen en PLT (Cuadro 4). En términos porcentuales, la producción promedio de leche total en el TIII, superó en 10,5% y 15,2% la producción de leche total en las vacas correspondientes a TII y TI, respectivamente.
La prueba de t comparativa de medias entre tratamientos para el porcentaje de grasa resultó en 2,129±0,93% para TI, 2,957±0,49% para TII y 1,657±0,37% para TIII, observándose diferencias estadísticamente significativas (p<0,05) para el TII al día 30 pp. Mientras que a los 60 dpp no se detectaron diferencias estadísticas.
Las vacas pertenecientes al TIII, retrasaron el reinicio de la ciclicidad ovárica durante el postparto temprano, destinando los mayores substratos energéticos recibidos hacia la producción de leche, posiblemente apoyada por mayor movilización y aportes adicionales de grasa corporal durante el periodo de transición.
En consecuencia, es posible que la distribución de la energía consumida en las vacas suplementadas solo durante el postparto con AGPI (TIII) fue derivada hacia la producción de leche. Mientras que en el grupo TII, las vacas distribuyeron mejor la energía aportada por la suplementación pre- y postparto hacia la actividad reproductiva; adicionalmente, estas vacas tuvieron una producción superior al grupo no suplementado (TI) y produjeron mayor contenido de grasa en la leche total, comparadas con el resto de las vacas (TI y TIII).
El hecho de haber una mayor producción de leche de las vacas TIII (suplementadas con AGPI postparto) podría indicar según Bernal-Santos et al. [31], que la suplementación con ácido linoleico conjugado en vacas lecheras durante la lactancia temprana, disminuyó la síntesis de grasa de la leche y pareciera responder a la partición de más nutrientes hacia la síntesis de leche (Cuadro 4), en lugar de mejorar el balance energético neto. De hecho, Santos et al. [18] señalan que la síntesis y secreción de hormonas, la ovulación de un folículo y el mantenimiento de un embrión en desarrollo, presentan costos energéticos mínimos en comparación con los costos de la lactancia. En nuestro experimento pudimos encontrar diferencias (p<0,05) para el promedio de porcentaje de grasa de la leche entre TII (2,957%) vs. TIII (1,657%) a los 30 dpp; sin embargo, hubo una diferencia de 2 kg de leche/d para el mismo período a favor de las vacas TIII, coincidiendo con la tendencia señalada por Santos et al.[26] durante el postparto temprano y con vacas Carora [1] y con vacas Holstein [14], concluyendo que la suplementación con AGPI y ácido linoleico conjugado (ALC) durante el período de transición (pre y/o postparto), redujo el porcentaje de grasa de la leche, las pérdidas de CC, pero incrementó el rendimiento de leche sobre el grupo control (suplementadas con aceite de palma), lo cual coincide con nuestros resultados en las vacas TII y TIII, las cuales produjeron más leche que las vacas control (TI). Estas diferencias se observan en la Figura 1 y en el Cuadro 4, respectivamente. Es importante resaltar en nuestro experimento la tendencia que las vacas del TIII, con mayores niveles de PLT promedio, mostraron menor dimensión ovárica y número de estructuras ováricas totales durante los primeros 15, 30 y 45 dpp. Contrariamente, las vacas TII, con menor PLT, alcanzaron una mayor dimensión ovárica y número de estructuras ováricas en el mismo periodo evaluado. Posiblemente, esto podría indicar una mejor adaptación metabólica y distribución energética en TII. En vacas Holstein de alta producción de leche, Dos Santos et al. [32] señalaron, una baja frecuencia de ciclicidad ovárica (17,6%) y menor tamaño de folículos dominantes en los primeros días postparto. Sin embargo, Audron et al. [33] encontraron que el tamaño ovárico y su dinámica dependió de la productividad de la vaca durante el periodo postparto, cuando fue mejor expresada la dinámica del sistema reproductivo a través de la mayor productividad.
Las vacas lecheras movilizan los tejidos corporales para apoyar la producción de leche y, debido a que el suministro de glucosa es limitado, los lípidos se utilizan preferentemente para la producción de energía. Durante el pico de la lactancia más del 80% de la glucosa disponible en el cuerpo del animal es fraccionada y dirigida hacia la glándula mamaria para la síntesis de leche. La actividad lipogénica se desactiva y los mecanismos lipolíticos en el tejido adiposo aumentan, a través de cambios en la expresión de varias enzimas clave. Esto da como resultado una pérdida de la CC, junto con altas concentraciones circulantes de ácidos grasos no esterificados. Los cambios en las vías de síntesis, secreción y señalización de las hormonas somatotróficas (insulina, hormona del crecimiento e IGF-I) y adipocinas (leptina) son claves para la regulación de estos procesos. Sin embargo, una alta dependencia de los ácidos grasos como fuente de energía durante el período peri-parto causa daño oxidativo a las mitocondrias en los tejidos metabólicamente activos, incluyendo el hígado y el tracto reproductivo. Es por ello que el exceso de acumulación de lípidos en los ovocitos y el endometrio en regeneración, reduce la fertilidad afectando la supervivencia del embrión y el aumento de los cambios inflamatorios, respectivamente [34]. Es por ello que los mecanismos de partición de energía son vitales en el equilibrio y balance energético del animal. En el caso de animales mestizos Carora, como en nuestro experimento, pareciera que la partición de la energía sigue otro tipo de patrón de distribución energética, sobre todo cuando el animal es suplementado durante el peri-parto, siendo este tipo de animal capaz de mantener tanto la función de reproducción como la producción láctea.
Conclusiones
La suplementación con AGPI durante el preparto-postparto favoreció las reservas energéticas del animal (condición corporal), impactando la actividad ovárica a nivel del crecimiento y desarrollo de las poblaciones foliculares (FOLII y FOLIII), como también el número de cuerpos lúteos (CL) en el grupo TII. Igualmente, esto se reflejó en un aumento de la dimensión del ovario izquierdo. La suplementación con AGPI durante el postparto (TIII) favoreció la PLT, pero no así la proporción de la grasa (%) en la leche. Sin embargo, en este grupo, aún cuando en los inicios del experimento (30 dpp), mostró retraso en la ciclicidad ovárica y en los DV, logró quedar gestante el 100%, al igual que las vacas TII. Las vacas no suplementadas con AGPI (TI) resultaron tener menos respuestas en CO, DV y PLT, lo cual demostró el beneficio de los AGPI, cuando se usa estratégicamente para mejorar los indicadores reproductivos y/o productivos.
Agradecimiento
Al MPCYT-FONACIT por el apoyo en el financiamiento a través del proyecto PEI2012-360.
Declaración de Conflicto de Intereses
Los autores declaran expresamente que no hubo conflicto de intereses durante el desarrollo de este trabajo.
Aporte de los Autores al Trabajo
CD: Redacción del manuscrito y análisis y discusión de resultados. AR: redaccion del manuscrito, aporte metodológico en el manejo de variables reproductivas. RP: Analisis de variables relacionadas con involucion uterina y actividad ovárica, aportes bibliograficos para la discusión. NM: Análisis e interpretacion de resultados en las corridas de progesterona. Aspectos estadisticos. LP: Interpretacion de resultados de producción de leche asociados con grasa de la leche. Aspectos estadisticos. TD: ayuda en la redacción del manuscrito y análisis global del trabajo.
Referencias
1. Hernández R, Díaz T. Las grasas sobrepasantes y su efecto sobre la actividad productiva y reproductiva en rumiantes. Innovacion & Tecnología en la Ganadería Doble Propósito, Fundación GIRARZ, C. González-Stagnaro, N. Madrid-Bury y E. Soto Belloso (eds). Ediciones Astro Data S.A. Capitulo XXXIII. 2011; 333-343. [ Links ]
2. Otto JR, Nish RO, Balogun MJ, Freeman BS, Malau-Aduli PA, Lane O, et al. Effect of dietary supplementation of pasture-based primiparous Holstein-Friesian cows with degummed crude canola oil on body condition score, liveweight, milk yield and composition. J App Ani Res. 2016; 44(1):194-200. [ Links ]
3. Pérez OR, Espinoza VJL, Palacios EA, Arjona LO, Palacios ME. Los ácidos grasos de la dieta afectan la fisiología reproductiva en la hembra bovina. Una revisión. Trop Subtrop Agro, 2012; 15:153-163. [ Links ]
4. National Research Council. 1989. Nutrient Requirements of Dairy Cattle. 6th. Rev Ed Natl Acad Sci, Washington, D.C. [ Links ]
5. Fattet IM, Jaurena G. El Estado Corporal de las Vacas Lecheras. Hemisferio Sur. Buenos Aires, Argentina. 1998; 8 p. [ Links ]
6. Diaz T, Schmitt EJP, de la Sota RL, Thatcher MJ, Thatcher WW. Human chorionic gonadotropin-induced alterations in ovarian follicular dynamics during the estrous cycle of heifers. J Anim Sci. 1998; 76:1929-1936. [ Links ]
7. Peters AR, Lamming GE. Regulation of ovarian function in the postpartum cow: An endocrine model. Vet Rec. 1986; 118: 236-239. [ Links ]
8. Jenkins T. Success of fat in dairy rations depends on the amount. Feedstuffs. 1997; 11-12. [ Links ]
9. Díaz T, Betancourt R, Hernández R, Gallo J. El efecto de las grasas omega 3 y omega 6 sobre la reproducción de vacas de primera lactancia pastoreando en los llanos venezolanos. En: E. Chacón y F. Espinoza (Eds). Memorias, III Simposium sobre Recursos y Tecnologías Alimentarias apropiadas para la Producción Bovina a Pastoreo en Condiciones Tropicales. Pasteurizadora Táchira C.A. (PASTCA). San Cristóbal. 2009; 1-28 CD-ROM. [ Links ]
10. Thatcher WW, Santos JEP, Silvestre FT, Kim IH, Staples CR. Perspective on physiological/endocrine and nutritional factors influencing fertility in post-partum dairy cows. Reprod Dom Anim. 2010; 45:2-14. [ Links ]
11. Staples CR, Thatcher WW. Effects of fatty acids on reproduction of dairy cows. In: Garnsworthy PC, Wiseman, J. (eds.). Recent Advances in Animal Nutrition. Nottingham University Press, UK. 2005; 229-256. [ Links ]
12. Arana GD, Uribe VLF. Estrategia para mejorar la CC en vacas de carne. Biosalud. 2012; 11: 71- 89. [ Links ]
13. Tyagi N, Thakur S, Shelke S. Effect of bypass fat supplementation on productive and reproductive performance in crossbred cows. Trop Anim Health Prod. 2010; 42:1749-1755. [ Links ]
14. Roodbari AR, Towhidi A, Zhand M, Rezayazdi K, Rahimi Mianji G, Dirandeh E, et al. Effect of conjugated linoleic acid supplementation during the transition period on plasma metabolites and productive and reproductive performances in dairy cows. Anim Feed Sci and Techn. 2016; 219:294-303. [ Links ]
15. Salehi R, Colazo MG, Oba M, Ambrose DJ. A prepartum diet supplemented with oilseeds high in oleic or linoleic acid reduced GnRH-induced LH release in dairy cows during second week postpartum. Reprod Bio Endocrin. 2015; 13:69. [ Links ]
16. Dirandeh EA, Towhidi S, Zeinoaldini M, Ganjkhanlou Z, Pirsarae A, Fouladi-Nashta A. Effects of different polyunsaturated fatty acid supplementations during the postpartum period of early lactating dairy cows on milk yield, metabolic responses, and reproductive performance. J Anim Sci. 2014; 91(2):713-21. [ Links ]
17. Palma RF, de Freitas Júnior JE, Rodrigues Gandra J, Maturana Filho M, Camargo Verdurico L, Navajás Rennó L, et al. Effect of unsaturated fatty acid supplementation on digestion, metabolism and nutrient balance in dairy cows during the transition period and early lactation. R Bras Zootec. 2014; 43(4):212-223. [ Links ]
18. Santos JEP, Bilby TR, Thatcher WW, Staples CR, Silvestre FT. Long chain fatty acids of diet as factors influencing reproduction in cattle. Reprod Dom Anim. 2008; 43:23-30. [ Links ]
19. Evans ACO, Mossa F, Walsh SW, Scheetz D, Jimenez-Krassel F, Ireland JLH, et al. Effects of maternal environment during gestation on ovarian folliculogenesis and consequences for fertility in bovine offspring. Reprod Dom Anim. 2012; 47 (Suppl. 4): 31-37. [ Links ]
20. Lucy MC, Staples CR, Michel FM, Thatcher WW, Bold TJ. Effect of fedding calcium soap to early postpartum dairy cow on plasma prostaglandin F2α, luteinizing hormone and follicular growth. J Dairy Sci 1991; 74: 483-489. [ Links ]
21. Robinson RS, Pushpakumara PGA, Cheng Z, Peters AR, Abayasekara DRE, Wathes DC. Effects of dietary polyunsaturated fatty acids on ovarian and uterine function in lactating dairy cows. Reproduction. 2002; 124:119-131. [ Links ]
22. Badiei A, Aliverdilou A, Amanlou H, Beheshti M, Dinanheh E, Moosakhani F, et al. Postpartum responses of dairy cows supplemented with n-3 fatty acids for different durations during the peripartal period. J Dairy Sci. 2014; 97(10):6391-6399. [ Links ]
23. Mossa F, Walsh SW, Butler ST, Berry DP, Carter F, Lonergan P, et al. Low numbers of ovarian follicles ≥3 mm in diameter are associated with low fertility in dairy cows. J Dairy Sci. 2012; 95:2355-2361. [ Links ]
24. Heravi Moussavi AR, Gilbert RO, Overton TR, Bauman DE, Butler WR. Effects of feeding fish meal and n-3 fatty acids on ovarian and uterine responses in early lactating dairy cows. J Dairy Sci. 2007; 90:145–154. [ Links ]
25. Coyral-Castel S, Ramé C, Fatet A, Dupont J. Effects of unsaturated fatty acids on progesterone secretion and selected protein kinases in goat granulose cells. Dom Anim Endocrin. 2010; 38: 272-283. [ Links ]
26. Piccinato CA, Sartori R, Sangsritavong S, Souza AH, Grummer RR, Luchini D, et al. In vitro and in vivo analysis of fatty acid effects on metabolism of 17b-estradiol and progesterone in dairy cows. J Dairy Sci. 2010; 93: 1934-1943. [ Links ]
27. Lemley CO, Butler ST, Butler WR, Wilson ME. Short communication: Insulin alters hepatic progesterone catabolic enzymes cytochrome P450 2C and 3A in dairy cows. J Dairy Sci. 2008; 91:641-645. [ Links ]
28. Cardona JCA, Ospina PH, Ramirez HMM, Pereira CH, Swanson K. Effects of supplementation with protected polyunsaturated fatty acids on productive and hormonal parameters of embryo recipient heifers. Bol Ind Anim. Nova Odessa. 2016; 73(2):150-158. [ Links ]
29. Atanasov B, Hostens M, Hajrulai-Musliu Z, Uzunov R, Adamov N, Davkov F, et al. Comparison of PUFA profiles in the blood and in follicular fluid and its association with follicular dynamics after PGF2a induced luteolysis in dairy cows. Mac Vet Rev. 2016; 39 (2): 175-183. Available online at www.macvetrev.mk. [ Links ]
30. Juchem SO, Cerri RLA, Villaseñor M, Galvão KN, Bruno RGS, Rutigliano HM, et al. Supplementation with calcium salts of linoleic and trans-octadecenoic acids improves fertility of lactating dairy cows. Reprod Dom Anim. 2010; 45: 55-62. [ Links ]
31. Bernal-Santos G, Perfield JW, Barbano DM, Bauman DE, Overton TR. Production responses of dairy cows to dietary supplementation with conjugated linoleic acid (CLA) during the transition period and early lactation. J Anim Sci. 2003; 86(10):3218-3228. [ Links ]
32. Dos Santos BJC, Kozicki LE, Grebogi AM, Weiss RR, Formighieri BMA, Saporski SM, et al. Productive and reproductive parameters in dairy cows and the relationship with ovarian follicular dynamics in the puerperium. Asian J Agr Food Sci 2014; 2:83-88. [ Links ]
33. Rekeit A, Tarvainis K, Račkauskait A, Juozaitien V, ilinskas H, ilaitis V. Milk production and age impact on ovarian activity during postpartum period. Veterinarija ir Zootechnika. 2016; 74-96. [ Links ]
34. Wathes DC, Clempson AM, Pollott GE. Associations between lipid metabolism and fertility in the dairy cow. Reprod Fertil Develop. 2013; 21(25):48-61. http://dx.doi.org/10.1071/RD12272. [ Links ]

