











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.27 n.9 Caracas set. 2002
IS CYCASIN IN Eumaeus minyas (LEPIDOPTERA: LYCAENIDAE) A PREDATOR DETERRENT?
Citlalli Castillo-Guevara and VÍctor Rico-Gray
Citlalli Castillo-Guevara. Ph.D. in Ecology and Management of Natural Resources. Address: Departamento de Ecología Vegetal, Instituto de Ecología, A.C., Apdo. 63, Xalapa, Veracruz 91070, México. e-mail: citlalli@ecologia.edu.mx
Víctor Rico-Gray. Ph.D., Tulane University. Senior Research Scientist, Departamento de Ecología Vegetal, Instituto de Ecología, A.C. e-mail: ricogray@ecologia.edu.mx
Summary
Sequestration of defense compounds from host plants may require particular physiological adaptations by larvae to ingest, accumulate, and store them. Defense compounds are used particularly against predators, where chemical defense reduces the risk of predation. Cycasin is a secondary metabolite present in cycads (Cycadales), which are host to butterflies of the genus Eumaeus (Lycaenidae). Cycasin is sequestered by the aposematic butterfly E. minyas from its host plant Zamia loddigesii (Zamiaceae) and used as a chemical defense. The presence of cycasin in all stadia of E. minyas was assessed in laboratory and field experiments. Through palatability experiments in the laboratory, it was established that pure cycasin deterred a potential predator (Solenopsis geminata, Formicidae) of E. minyas, and that cycasin in egg, larva and adult extracts deterred this predator. To assess the efficiency of this chemical defense for eggs and larvae in natural conditions, a predator exclusion experiment was carried out. Using survival analysis it is shown that the number of surviving eggs and larvae were significantly higher in the exclusions. This suggests that the protective function of cycasin in natural conditions is not totally effective against natural predators. This is the first study that tests experimentally the protective function of cycasin in natural conditions. We suggest that cycasin in the aposematic butterfly E. minyas has a defensive function against predators; however, its populations may be regulated by the negative effect of its predators, which apparently circumvent the chemical defensive barrier, and by cannibalism.
Resumen
El secuestro de compuestos de defensa de las plantas hospederas puede requerir adaptaciones fisiológicas particulares para que las larvas los ingieran, acumulen y almacenen. Los compuestos de defensa son usados para varios propósitos, particularmente contra depredadores, donde la defensa química reduce el riesgo de depredación. La cicasina es un metabolito secundario presente en las cícadas (Cycadales), que son hospederas de mariposas del género Eumaeus (Lycaenidae). La cicasina es secuestrada por la mariposa aposemática E. minyas de su planta hospedera Zamia loddigesii (Zamiaceae) y es usada como defensa química. En experimentos de campo y laboratorio se evaluó la presencia de cicasina en todos los estadios de E. minyas. Mediante experimentos de palatabilidad en el laboratorio, se estableció que la cicasina pura repele a Solenopsis geminata (Hymenoptera: Formicidae), un depredador potencial de E. minyas y, al utilizar extractos de huevos, larvas y adultos de esta mariposa se obtiene el mismo efecto de manera experimental. Para evaluar la eficiencia de esta defensa química en huevos y larvas en condiciones naturales, se realizó un experimento de exclusión de depredadores. Usando un análisis de sobrevivencia se demostró que el número de huevos y larvas sobrevivientes fue significativamente más alto en las exclusiones, sugiriendo que la función protectiva de la cicasina en condiciones naturales no es totalmente efectiva contra los depredadores naturales. Este es el primer estudio que incluye manipulación experimental de la función protectiva de la cicasina en condiciones naturales. Sugerimos que la cicasina en la mariposa aposemática E. minyas tiene una función defensiva contra los depredadores; sin embargo, sus poblaciones pueden estar reguladas por el efecto negativo de sus depredadores, los cuales aparentemente saltan la barrera química defensiva y por el canibalismo.
Resumo
O sequestro de compostos na defesa das plantas hospedeiras, pode exigir adaptaçoes fisiológicas particulares para que as larvas os comam, acumulem e armazenem. Os compostos de defesa são usados para vários propósitos, particularmente contra depredadores, onde a defesa química reduz o risco de depredação. A cicasina é um composto metabólico secundário presente nas cícadas (Cycadales), as quais são hospedeiras de mariposas do género Eumaeus (Lycaenidae). A cicasina é sequestrada pela mariposa aposemática Eumaeus minyas de sua planta hospedeira Zamia loddigesii (Zamiaceae) e é utilizada como defesa química. Utilizando-se experimento de campo e laboratório, avaliou-se a presença de cicasina em todos os estágios de E. minyas. Mediante experimentos de palatabilidade em laboratório, estabeleceu-se que a cicasina pura repele a Solenopsis geminata (Hymenoptera: Formicidae), que é um depredador potencial de E. minyas e, ao utilizar extratos de ovos, larvas e indivíduos adultos desta mariposa, se obtém o mesmo efeito. Para avaliar a eficiência desta defesa química em ovos e larvas realizou-se um experimento de exclusão de depredadores em condições naturais. Usando-se uma análise de sobrevivência, demonstrou-se que o número de ovos e larvas sobreviventes foi significativamente mais alto nas excluções, sugerindo que a função de proteção da cicasina em codições naturais nao é totalmente efetiva contra os depredadores. Este é o primeiro estudo realizado que inclui manipulação experimental da função protetora da cicasina em condição natural. Sugerimos que a cicasina presente na mariposa aposemática E. minyas, tem uma função de defesa contra os depredadores, mas as suas populaçoes podem estar reguladas pelo efeito negativo de seus depredadores, os quais, aparentemente saltam a barreira química defensiva, e pelo canibalismo.
KEYWORDS / Aposematic Butterfly / Cycads / Deterrent Effect of Cycasin / Lycaenidae / Predation /
Received: 03/01/2002. Modified: 07/30/2002. Accepted: 08/08/2002
Larvae exhibit a wide range of defensive strategies to avoid being eaten, e.g., mimetic coloration, shelter construction, unpalatability due to urticating hairs, spines and defensive glands, regurgitation, chemicals sequestered from host plants, secretion of volatiles, and noise production (see Brower, 1984; Bowers, 1993). Not only do they use their bad taste or unpleasant odor as a defense, they also announce it to potential predators by means of a conspicuous coloration, and gregarious and sedentary behavior (Bowers, 1993). Sequestration of defense compounds from larval host plants may require particular physiological adaptations by larvae to ingest, accumulate, and store those compounds (Brattsten, 1986; Bowers, 1992). Defense compounds are used for various purposes, particularly against predators, (Bowers, 1990; Duffey, 1980; Blum, 1983; Brower, 1984). Research on acquisition of chemical defenses by insects, particularly in Lepidoptera, has been done using adult individuals. However, it is usually during the larval stage that chemical defenses, sequestered from host plants, are ingested, processed and stored (Bowers, 1993). Examples of chemical defense of Lepidoptera larvae are well known (Brower, 1984; Bowers, 1990; Witz, 1990).
In aposematic species unpalatability is coupled with a warning coloration which can have many consequences for the biology and ecology of these species. Many "chemically defended" Lepidoptera are aposematic, and store plant compounds that are known vertebrate toxins, such as cardenolides (Brower, 1984), alkaloids (Rothschild et al., 1979; Boppre, 1990; Montllor et al., 1990), and cyanogens (Jones et al., 1962). Chemical defense of aposematic insects has also been shown to be effective against invertebrate predators, which can learn to subsequently avoid similar prey (Montllor and Bernays, 1993).
Cycasin is a secondary metabolite present in cycads (Cycadales), belonging to the azoxyglycosid group (Matsumoto and Strong, 1963; Whiting, 1963; Kobayashi and Matsumoto, 1965; Morgan and Hoffman, 1983; Norstog and Nicholls, 1997; Jones, 2000). Species of the American genera Zamia, Ceratozamia and Dioon (Cycadales: Zamiaceae) are hosts to aposematic butterflies of the genus Eumaeus (Lepidoptera: Lycaenidae). It has been demonstrated that E. atala sequesters cycasin from Z. floridana, which is later used as a defense against both vertebrate (Bowers and Farley, 1990) and invertebrate (Rothschild et al., 1986; Bowers and Larin, 1989) predators. These results were based on laboratory experiments, but no field research has confirmed them. It has also been suggested that cycasin works in E. minyas as a defense mechanism, providing chemical protection (Clark and Clark, 1991; Nash et al., 1992; DeVries, 1994). However, this has not been tested and the life cycle of E. minyas remains undescribed. We present the results of laboratory and field experiments to evaluate the protective function of cycasin in the aposematic butterfly E. minyas. In particular, the following questions are addressed: In which stages of its life cycle does E. minyas contain cycasin? Does pure cycasin repel potential predators of E. minyas? Does cycasin turn eggs, larvae and adults unpalatable? Is cycasin efficient in protecting eggs and larvae from their natural predators under natural conditions? This research is part of a wider study on the interactions between E. minyas and E. debora and their host plants Z. loddigesii and D. edule.
Materials and Methods
Study site
Field work was accomplished in an oak forest near Chavarrillo, in central Veracruz, México (19º24N, 96º48W; 1000m altitude), characterized by a calcareous-derived soil and abundant rock outcrops. The climate is temperate-humid, mean annual temperature is 24.5C°, total annual precipitation is ca. 1110mm, with a rainy season between June and September and an extended six-month dry season (García, 1964; Soto et al., 1996). The vegetation is characterized (Flores, 1995) by a mixture of oak forest [Quercus oleoides, Q. laurina, Q. peduncularis (Fagaceae), Nectandra sanguinea (Lauraceae), Bursera simaruba (Burseraceae)] and palm groves of Brahea dulcis (Arecaceae). Epiphytes are present in the Araceae, Bromeliaceae, Orchidaceae and Cactaceae (Castillo, 1985). The main shurbs and herbs are (Flores, 1995) Acacia cornigera, A. pennatula (Mimosaceae), Zamia loddigesii, and Dioon edule (Zamiaceae).
Species
Zamia loddigesii (Zamiaceae, Miquel 1843) (voucher, L. M. Whitelock 11/26/1963, XAL) is a small plant (up to 1m tall) with 1 to 6 fronds, inhabiting tropical dry and deciduous forests and secondary vegetation (Vovides et al., 1983). It is distributed along the coast of the Gulf of Mexico up to Guatemala, from 0 to 1000m in elevation (Vovides et al., 1983; Jones, 2000). Z. loddigesii is protected under the status of threatened species both internationally (CITES Appendix II) and nationally (Annonymous, 1994).
Eumaeus minyas (Lycaenidae, Hübner 1809) is distributed from México to Costa Rica, where the larvae have been reported to consume fronds and female reproductive cones from Z. furfuracea, Z. skinnery and Z. loddigesii (DeVries, 1976; 1983a; Clark and Clark, 1991). E. minyas is considered aposematic, exhibiting a flashy warning coloration (DeVries, 1977; Clark and Clark, 1991; Nash et al., 1992).
Chemical analyses
Chromatography on silica-gel plates (Whatman PE SIL G/UV) was used to determine presence or absence of cycasin in all stages of E. minyas. From individuals collected in November 1999, we used 36 eggs (0.0293g), 16 larvae of the four instars (2.045g), 1 pupa (0.1039g), and 5 adults (0.5351g). The fresh samples were processed using the technique described by Yagi et al. (1980) and Bowers and Larin (1989). For comparison we used standard pure cycasin (Biochemical 66950), and computed the reference factor (Rf) for each sample. Vouchers are deposited in a personal collection at the Departamento de Ecología Vegetal, Instituto de Ecología, A.C.
Field experiments
Palatability experiments, modified from Bowers and Larin (1989) were conducted using individuals of Solenopsis geminata (Hymenoptera: Formicidae, Fabricius 1804) as predators. This ant inhabits the study site and was observed on Z. loddigesii. Before the experiments, ants were collected from a colony in Francisco Javier Clavijero Botanical Garden (Xalapa, Veracruz, México; 19º30N, 96º57W; 1280m altitude). The sample included workers, soldiers and larvae in order to ensure that ant behaviour was the least affected by sampling, (Jorge Valenzuela-González, personal communication). Ants were placed in 25 x 13cm plastic containers covered with mesh, and were deprived of food 48h before the experiments. The containers were then uncovered and placed within a square (0.26m2) of Tanglefoot® (Tanglefoot Co., Grand Rapids, MI, USA), where different food solutions were offered to the ants (see below).
To test if pure cycasin deterred S. geminata individuals, ants were offered two solutions, 1) control (0.75g sucrose + 0.30ml water), and 2) experimental (0.75 g sucrose + 0.30ml of 1mg/ml cycasin in water). Cycasin was the same as that used for chemical analysis. The number of ant visits to each solution was recorded during 90 min. When an ant lowered its head and touched a solution, it was considered a visit. The experiment was replicated eight times with different ants in the same conditions mentioned above.
To test whether cycasin in eggs, larvae and adults rendered them unpalatable and deterred S. geminata individuals, individuals of E. minyas were collected in the study site: 33 eggs (0.02g), 15 larvae of the four instars (0.70g), and 2 adults (0.20g). Extracts were prepared with EtOH 70%, separately for each stage. These were used to prepare three experimental solutions (0.75g sucrose + 0.30ml of egg, larval or adult extract). Ants collected previously were offered four solutions, a control one (0.75g sucrose + 0.30ml water), and three experimental ones (with egg, larva or adult extract). This experiment was also replicated eight times, with different ants in the same conditions and the number of ant visits to each solution was recorded during 90 min trial periods.
To test whether the presence of 70% ethanol used in the extraction procedure affected predator behavior, or if it masked the presence or cancelled out the effect of cycasin, we counted visits of individuals of S. geminata to a control (0.75g sucrose + 0.30ml water) and to an experimental solution (0.75g sucrose + 0.30ml 70% EtOH). The number of visits were transformed using the square root of X + 0.5, no significant differences (ANOVA, F1,14= 0.075, p= 0.7877) were found in the number of visits between solutions (control, X= 84.375 +5.123, N= 675; experimental, X= 86.875 +6.465, N= 695).
To test the defensive function of cycasin under natural conditions, a predator exclusion experiment was conducted at the study site. Egg clusters of E. minyas were located on individuals of Z. loddigesii and three predator exclusion treatments were applied: 1) fronds covered with mesh, 2) fronds with a band of Tanglefoot® at the base, and 3) fronds with Tanglefoot® and mesh; the control were fronds with egg clusters under natural conditions. Different treatments were used to determine differential effects of the different groups of predators (mainly for birds and ants, but other flying and crawling insects as well). Eighty-three eggs (total for the study site at the time) grouped in egg clusters located on 15 Z. loddigesii individuals were used, 32 on control plants and 51 on treatment plants (16 with mesh, 19 with Tanglefoot®, 16 with Tanglefoot® and mesh). Over 15 days (the time for egg hatching), the plants were visited every day and the number of eggs preyed upon, hatched and unhatched was registered. The exclusion experiment was continued applying the same treatments to the recently hatched larvae, plus 15 more first-instar larvae collected elsewhere in the study site. A total of 70 larvae, 26 on control plants and 44 on treatment plants (19 with mesh, 15 with Tanglefoot®, 10 with Tanglefoot® and mesh) were observed. The number of larvae that had disappeared, died or still survived was counted daily. To establish the larval instar, larval length every third day until pupation (ca. 16 days) was measured. Observations were extended up to 32 days because different-aged cohorts were observed. The number of resulting pupae and of larvae that reached adulthood was also counted.
Statistical analyses
The total number of visits to a particular solution was analyzed with a heterogeneity x2-test (Zar, 1999) to determine whether pure cycasin deterred potential predators. The number of visits to a particular solution were transformed (square root of X + 0.5) and then analyzed with a one-way ANOVA (Abacus Concepts, Inc. 1996; Zar, 1999) to determine whether the presence of cycasin in butterflies offered protection.
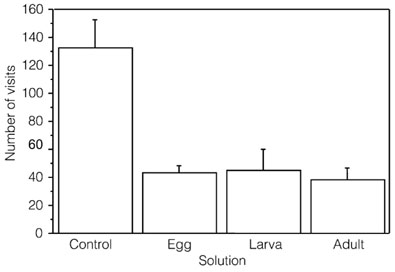
Figure 1. Mean (+SE) number of visits by experimental ant predator, Solenopsis geminata, to control (0.75g sucrose + 0.30ml water) and experimental (0.75 g sucrose + 0.30ml of egg, larva and adult extracts of E. minyas) solutions.
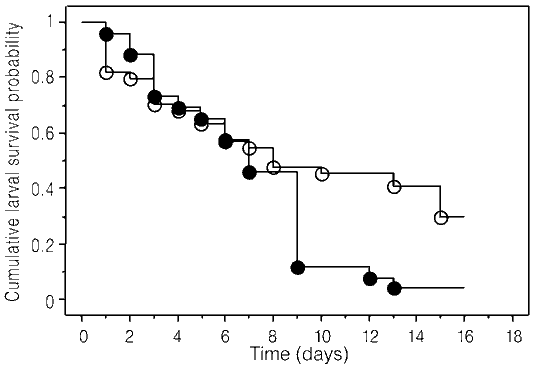
Figure 2. Survival curves for E. minyas larvae with and without exclusion of predators. Treatments of exclusion: mesh, tanglefoot, tanglefoot and mesh. Without exclusion: control. Time is period of larval state (16 days). Predation events with exclusion, Predation events without exclusion.
To test for the efficiency of cycasin as a chemical defense of eggs under natural conditions, the percentage of surviving eggs was computed. To test cycasin as a chemical defense of larvae in natural conditions, we used a survival or failure time analysis (Muenchow, 1986; Pyke and Thompson, 1986). The analysis calculates the probability of the occurrence of predation events during one observation period. The start of the observations was designated as time zero, and the predation events, disappearance of larvae, were subsequently counted over 16 days. If a given larva was preyed upon, it was considered an uncensored data point, and, if predation never occurred, i.e., the larvae survived, it was considered a censored data point. To compute the functions among variables the product-limit Kaplan-Meier nonparamethric method (Abacus Concepts, Inc. 1996) was used, and the log-rank statistic (Mantel-Cox) was used to test for differences between the control and a given treatment (treatments were grouped). An among-treatments comparison showed that >20% of the events were censored and, as suggested by Pyke and Thompson (1986), instead of the survival analysis we used a 3x2 contingency table (Zar, 1999). All results are shown as mean + standard error.
Results
Life history
The only herbivore observed foraging on the fronds of Z. loddigesii was E. minyas. During the rainy season Z. loddigesii is frequently visited by E. minyas. Similar to other cycads (Norstog and Nicholls, 1997), Z. loddigesii grows slowly and irregularly (only a few centimeters per year) and produces one to two fronds per year during the rainy season.
Adult butterflies of E. minyas oviposit on new fronds, of which larvae may consume up to 90% during their development. Larvae may also forage on the raquis and, while uncommon, butterflies may also oviposit on female reproductive cones, which are also harmed by larvae.
Eggs are deposited in small clusters (mean of egg by cluster= 5.238 +0.547, N= 42), they are pale pink, turning white after one day, and hatch after approximately 11 days. Larvae are bright red with seven transversal yellow stripes and undergo four instar changes in 16 days. Pupae are orange with black marks, pupation lasting approximately 19 days. Larvae and pupae are gregarious (DeVries, 1976; 1977; 1983b). The distinctive characteristics of adults are an orange-red abdomen, and black wings with metallic marks and white margins. It takes approximately 46 days from oviposition to adulthood.
Cycasin detection and palatability experiments
Cromatographic comparison indicated that cycasin was detected in all stages of the life cycle of E. minyas: standard Rf= 71.6; eggs= 71.0; larvae= 63.1; pupa= 53.7; and adults= 63.3. Variation in larvae and pupae were attributed to differences in the molecular structure of cycasin due to metabolic changes within a stage. There are some differences in pattern due to the metabolic conjugation in the insect of some of the plant components (Harborne, 1988). These variations in Rf were also observed in evaluations in the variations of cardenolide content of adult monarch butterflies by comparing the cromatographic separation of extracts from insects and from food plants (Harborne, 1988).
Visits of S. geminata were significantly higher to the water/sucrose solution than to the water/pure cycasin solution (x2= 80.7249, p<0.001). Similarly, significant differences (ANOVA, F3,28= 11.591, p<0.0001) were found (Figure 1) among ant visits to the different solutions prepared with E. minyas extracts and the control solution: sucrose/water (X= 132.37 +19.77, N= 1059), egg extract (X= 43.50 +4.89, N= 348), larva extract (X= 45.25 +14.57, N= 362), and adult extract (X= 38 +8.79, N= 304). There were significantly more visits to the water/sucrose solution (Fishers, p<0.001), and there were no significant differences among the other treatments (Fishers, p>0.05).
In situ predator exclusion experiment
No predation was found in the exclusions, as all of the 51 eggs in exclusion survived. However, out of 32 eggs in control plants 22 (68.75%) survived and 10 (31.25%) were predated. Three eggs fell from the frond due to heavy rain, 15 did not hatch, and 55 hatched (i.e., 66% passed from egg to 1st instar larvae). Hatching time was 11 days (X= 11.28 +0.39, N= 18), and it took from one to four days for all eggs in a cluster to hatch (X= 1.85 +0.27, N= 13).
Fifty six of the initial 70 larvae were killed. In 26 larvae of control plants, 25 (96.15%) were predated, and in 44 larvae of exclusion plants, 31 (70.45%) were predated. The number of surviving larvae (Figure 2) was significantly higher (x21= 4.72, p= 0.029) when predators were excluded (N= 13) than when they were not (N= 1). We found no significant differences (x22= 1.31, p>0.05) in predation events among exclusion treatments (mesh= 14, Tanglefoot®= 9, Tanglefoot®+ mesh= 8). Only 14 (20%) of the original 70 larvae pupated, and all but one pupa reached adulthood, a process lasting 19 days (X= 19.44 +0.24, N= 16).
Cannibalism was an important factor in the survival of E. minyas. In one predator exclusion treatment (Tanglefoot® + mesh), 80% mortality was attributed to cannibalism.
Discussion
The results show for the first time that all stages of E. minyas contain cycasin that is sequestered from its host plant Z. loddigesii. Moreover, it is shown that pure cycasin and cycasin in E. minyas (eggs, larvae and adult) can deter a potential predator (S. geminata) under laboratory conditions. This is similar to findings by Bowers and Larin (1989) using E. atala and the ant Camponotus abdominalis floridanus (Hymenoptera: Formicidae) which, however, were not obtained under natural conditions. Our results show that predation in natural conditions had a negative effect on the survival of E. minyas, suggesting that some predators can avoid the protective chemical barrier provided by cycasin.
Our results suggest a pattern. When the Rf of a sample (egg, larva, pupa or adult) approaches that of the standard, the defense attributed to cycasin increased, whereas when the Rf is different the defense decreased. In the predation experiment under natural conditions it was evident that the egg stage is the most protected and its Rf was similar to the standard, whereas the Rf of the larval stage was lower than the standard, and predation of larvae was higher. Similar experiments using pupae and adult butterflies, are needed in order to evaluate predation on these stages.
The results of the predation experiment suggest that the impact of predation was different for the different stages. While predation on eggs (control) amounted to 31.25%, predation on larvae was 96.15%. Furthermore, 21% of the eggs did not hatch, which is comparatively low considering the reported 53% non-hatched eggs for E. minyas in Costa Rica (Clark and Clark, 1991). Furthermore, eggs are subject to predation but also to environmental factors. Some eggs slid off from the fronds due to heavy rain, eggs may dry up during days of extreme high temperature (>30ºC), and eggs and larvae were subject to attack by fungi like Beauveria bassiana (Bals.) Vuill., Deuteromycota. Egg mortality can also be attributed to factors intrinsic to the adult. For example, feeding quality and quantity while larvae or adult can significantly influence egg production and maturation (Scott, 1986; Braby and Jones, 1995). The egg stage is critical for the future survival of an E. minyas individual, the more eggs that hatch into larvae the higher the probability of surviving the larval stage, which, no doubt, is the most vulnerable and critical stage for the survival of an E. minyas individual (96.15% predation in the control).
The mortality rate of adults is important as every additional day of survival increases reproductive potential (Brower, 1984). Predation was significant on 3rd-and 4th-instar larvae (between 8 and 16 days of development). Under natural conditions, predation was significantly higher on non-protected larvae. However, predation in 1st– and 2nd-instar larvae did not differ between protected and non-protected conditions, and death of these larvae should be attributed to factors other than predation. Younger larvae are smaller and probably less conspicuous than older, larger, larvae (e.g., length of 3rd instar, X= 15.89mm +0.64, N= 42; 4th instar, X=21.97mm +0.64, N= 32; C. Castillo-Guevara, unpublished). No significant differences were obtained among exclusion treatments that would enable us to suggest the types of predators involved, although a number of ant species (Pheidole sp., Camponotus sericeiventris, Solenopsis geminata, and Ectatomma tuberculatum, Hymenoptera: Formicidae), were collected in the study site. S. geminata and E. tuberculatum are carnivorous and potential predators (Valenzuela-González et al., 1995).
Each stage in the life cycle of lepidopterans has a specific guild of predators, and the mechanisms used to avoid them seem to be mostly related to visually-hunting vertebrate predators (Rausher, 1980; 1981). However, the first instars of many lepidopterans are predated by a wide range of invertebrates, mostly night feeders (Dempster, 1984), and thus camouflage and warning coloration may not be a good defense mechanism.
Cannibalism was important to the survival of E. minyas larvae, especially when they were fully confined (Tanglefoot® + mesh treatment). Ingestion of conspecifics can result in a direct and crucial metabolic gain when lipid accumulation is not enough to support stress due to food shortage, but also in an indirect gain, the reduction of competitors (Polis, 1981; Wagner and Wise, 1996). Cannibalism in groups of E. minyas was also observed in Costa Rica and Panama; those slowest to molt were eaten by the fast-molting members (Phil DeVries, personal communication). We suggest that cannibalism among E. minyas larvae is a direct, easier and less energy-costly way to sequester cycasin.
As toxin levels vary considerably over short distances and through time, palatability to predators can also vary spatiotemporally (Harborne, 1999). Even though predators have developed behaviors and physiological responses to recognize unpalatable prey and thus decrease the effect of defenses (Calvert et al., 1979; Fink and Brower, 1981), a decrease in availability of palatable prey may induce predators to forage on less palatable prey (Alcock, 1970; Boyden, 1976). Moreover, young predator individuals may sporadically forage on unpalatable prey, or predators may forget a learned behavior (González et al., 1967).
It is not clear how cycasin works protecting lepidopterans against predators. The responses in predators include those caused by cardiac glycosides (vomiting) or pyrrolizidine alkaloids (bitter taste), which mask the real taste of cycasin (Nash et al., 1992). In addition, the presence of pyrazines (in E. atala) can enhance the protection offered by cycasin (Rothschild, 1984; Rothschild et al., 1986). Certain insects (e.g., Seiractia echo, Lepidoptera: Arctiidae) can circumvent the protective action of the cycasin produced by Zamia floridana (Teas, 1967), supporting the idea that no chemical defense against predators is absolute (Steward and Keeler, 1988; Agrawal, 1998). However, in the Eumaeus-Zamia interaction the level of cycasin is higher in Eumaeus than in the host plant (Rothschild et al., 1986). Production of secondary metabolites is not related only to defense against vertebrate or invertebrate herbivores, the system is even more complex and evidence for these substances under natural conditions is currently lacking and difficult to obtain. Furthermore, even though not tested, there seems to be a close association between warning coloration and unpalatability in E. minyas. First instar larvae are pale orange, and after three days of feeding on Z. loddigesii fronds and sequestering cycasin their color changes to bright red; consequently, they are more conspicuous.
Specialization on the Zamiaceae and the warning attributes in Eumaeus larvae may suggest a long association through evolutionary time. However, protection by cycasin in eggs and larvae could have evolved associated to predators that are currently extinct and thus extant predators are not so easily warned. Finally, we suggest that cycasin in the aposematic butterfly E. minyas has a defensive function against predators, however, its populations may be regulated by the negative effect of its predators, which apparently avoid the chemical defensive barrier.
ACKNOWLEDGEMENTS
The authors thank J. López-Portillo, J.G. García-Franco, V. Parra-Tabla and J.F. Ornelas for their suggestions to earlier drafts of the manuscript; S. Cervantes and V. Vázquez for their help with field and laboratory work, G. Carrión for fungi identification; and J. Valenzuela-González for ant identification and suggestions for experiments. We specially thank C. Lara for his help in the field and for his suggestions for statistical analyses. This research was supported by IdeaWild, Instituto de Ecología, A.C. (902-16) and a CONACYT (112677) scholarship to CCG.
REFERENCES
1. Abacus CI (1996) Abacus Concepts, Stat View Reference. Berkeley, California. 451 pp. [ Links ]
2. Agrawal AA (1998) Induced responses to herbivory and increased plant performance. Science 279: 1201-1202. [ Links ]
3. Alcock J (1970) Punishment levels and the response of black-capped chickadees (Parus atricapillus) to three kinds of artificial seeds. Anim. Behav. 18: 592-599. [ Links ]
4. Annonymous (1994) Norma Oficial Mexicana NOM-059-ECOL-1994, que determina las especies y subespecies de flora y fauna silvestres terrestres y acuáticas en peligro de extinción, amenazadas, raras y las sujetas a protección especial, y que esTablece especificaciones para su protección. Diario Oficial de la Federación 488: 2-60. [ Links ]
5. Blum MS (1983) Detoxication, deactivation, and utilization of plant compounds by insects. In Hedin PA (Eds.) Plant resistence to insects. ACS Symposium Series 208. American Chemical Society. Washington, D.C. pp. 265-275. [ Links ]
6. Boppre M (1990) Lepidoptera and pyrrolizidine alkaloids. J. Chem. Ecol. 16: 165-186. [ Links ]
7. Bowers MD (1990) Recycling plant natural products for insects defense. In Evans DL, Schmidt JO (Eds.) Insects defenses, adaptative mechanisms and strategies of prey and predators. State University of New York. Albany, NY. pp. 353-386. [ Links ]
8. Bowers MD (1992) Unpalatability and the cost of chemical defense in insects. In Roitberg B, Isman MB (Eds.) Chemical ecology of insects: an evolutionary approach. Chapman and Hall. New York. pp. 216-244. [ Links ]
9. Bowers MD (1993) Aposematic caterpillars: life-styles of the warnigly colored and unpalaTable. In Stamp N, Casey TM (Eds.) Caterpillars. Ecological and Evolutionary Constraints on Foraging. Chapman and Hall. London. pp. 331-371. [ Links ]
10. Bowers MD, Larin Z (1989) Acquired chemical defense in the lycaenid butterfly, Eumaeus atala. J. Chem. Ecol. 15: 1133-1146. [ Links ]
11. Bowers MD, Farley S (1990) The behaviour of grey jays, Perisoreus canadensis, towards palaTable and unpalaTable Lepidoptera. Anim. Behav. 39: 699-705. [ Links ]
12. Boyden TC (1976) Butterfly palatability and mimicry: experiments with Ameiva lizards. Evolution 30: 73-81. [ Links ]
13. Braby MF, Jones RE (1995) Reproductive patterns and resource allocation in tropical butterflies influence of adult diet and seasonal phenotype on fecundity, longevity and egg size. Oikos 72: 189-204. [ Links ]
14. Brattsten LB (1986) Fate of ingested plant allelochemicals in herbivorous insects. In Brattsten LB, Ahmad S (Eds.) Molecular aspects of insects plant associations. Plenum. New York. pp. 211-255. [ Links ]
15. Brower LP (1984) Chemical defense in butterflies. In Vane-Wright R, Ackery P (Eds.) The Biology of Butterflies. Academic Press. London. pp. 109-134. [ Links ]
16. Calvert WH, Hedrick LE, Brower LP (1979) Mortality of the monarch butterfly due to avian predation at five overwintering sites in Mexico. Science 204: 847-851. [ Links ]
17. Castillo G (1985) Integración de paisajes en la región de Jalcomulco, Ver. Thesis, Universidad Veracruzana-Xalapa. México. 110 pp. [ Links ]
18. Clark DB, Clark DA (1991) Herbivores, herbivory, and plant phenology: patterns and consequences in a tropical rain-forest cycad. In Price PW, Lewinsohn TM, Fernandes GW, Benson WW (Eds.) Plant-Animal Interactions: Evolutionary Ecology in Tropical and Temperate Regions. John Wiley. New York. pp. 209-225. [ Links ]
19. Dempster JP (1984) The natural enemies of butterflies. In Vane-Wright R, Ackery P (Eds.) The Biology of Butterflies. Academic Press. New York. pp. 97-104. [ Links ]
20. DeVries PJ (1976) Notes on the behavior of Eumaeus minyas (Hübn.) (Lepidoptera: Lycaenidae) in Costa Rica. Brenesia 8: 103. [ Links ]
21. DeVries PJ (1977) Eumaeus minyas Hübner: an aposematic lycaenid butterfly. Brenesia 12/13: 269-270. [ Links ]
22. DeVries PJ (1983a) Zamia skinneri and Zamia fairchildiana (zamia, palmera siempre verde, cycad). In Janzen DH (Ed.) Costa Rican Natural History. University Chicago Press. Chicago. pp. 349-350. [ Links ]
23. DeVries PJ (1983b) Checklist of butterflies. In: Janzen DH (Ed.) Costa Rican Natural History. University Chicago Press. Chicago. pp. 654-678. [ Links ]
24. DeVries PJ (1994) Patterns of butterfly diversity and promising topics in natural history and ecology. In McDade LA, Bawa KS, Hespenheide HA, Hartshorn GS (Eds.) La Selva. Ecology and Natural History of a Neotropical Rain Forest. University Chicago Press. Chicago. pp. 187-194. [ Links ]
25. Duffey SS (1980) Sequestration of plant natural products by insects. Ann. Rev. Entomol. 22: 447-477. [ Links ]
26. Fink LS, Brower LP (1981) Birds can overcome the cardenolide defense of monarch butterflies in Mexico. Nature 291: 67-70. [ Links ]
27. Flores PA (1995) Biología reproductiva de Ryncholaelia glauca (Lindl.) Schltr. (Orchidaceae: Laeliinae) en un encinar del centro de Veracruz. Thesis. Universidad Veracruzana-Xalapa. México. 50 pp. [ Links ]
28. García E (1964) Modificaciones al Sistema de Clasificación Climática de Koeppen (para adaptarlo a las condiciones de la República Mexicana). Universidad Nacional Autónoma de México, México. 246 pp. [ Links ]
29. González RC, Behrend ER, Bitterman ME (1967) Reversal learning and forgetting in birds and fish. Science 158: 519-521. [ Links ]
30. Harborne JB (1988) Introduction to ecological biochemistry. Academic Press. San Diego. pp. 356. [ Links ]
31. Harborne JB (1999) Recent advances in chemical ecology. Nat. Prod. Rep. 16: 509-523. [ Links ]
32. Jones DA, Parsons J, Rothschild M (1962) Released hydrocyanic acid from crushed tissues of all stages of the life cycle of species of Zygaeninae (Lepidoptera). Nature 193: 52-53. [ Links ]
33. Jones DL (2000) Cycads of the World. Smithsonian Institution. Washington, DC. pp 312. [ Links ]
34. Kobayashi A, Matsumoto H (1965) Studies on methylazoxymethanol, the aglycone of cycasin. Isolation, biological, and chemical properties. Arch. Biochem. Biophys. 110: 373-380. [ Links ]
35. Matsumoto H, Strong FM (1963) The occurrence of methylazoxymethanol in Cycas circinalis L. Arch. Biochem. Biophys. 101: 229-310. [ Links ]
36. Montllor CB, Bernays EA (1993) Invertebrate predators and caterpillar foraging. In Stamp N, Casey TM (Eds.) Ecological and Evolutionary Constraints on Foraging. Chapman and Hall. London. pp. 170-202. [ Links ]
37. Montllor CB, Bernays EA, Barbehem RV (1990) Importance of quinolizidine alkaloids in the relationships between larvae of Uresiphita reversalis (Lepidoptera: Pyralidae) and a host plant, Genesia monspessulana. J. Chem. Ecol. 16: 1853-1865. [ Links ]
38. Morgan RW, Hoffmann GR (1983) Cycasin and its mutagenic metabolites. Mutation Res. 114: 19-58. [ Links ]
39. Muenchow G (1986) Ecological use of failure time analysis. Ecology 67: 246-250. [ Links ]
40. Nash RJ, Bell EA, Ackery PR (1992) The protective role of cycasin in cycad-feeding lepidoptera. Phytochemistry 31: 1955-1957. [ Links ]
41. Norstog KJ, Nicholls TJ (1997) The Biology of the Cycads. Cornell University. New York. pp 363. [ Links ]
42. Polis GA (1981) The evolution and dynamics of intraespecific predation. Ann. Rev. Ecol. Syst. 12: 225-251. [ Links ]
43. Pyke DA, Thompson JN (1986) Statistical analysis of survival and removal rate experiments. Ecology 67: 240-245. [ Links ]
44. Rausher MD (1980) Host abundance, juvenil survival, and oviposition preference in Battus philenor. Evolution 34: 342-355. [ Links ]
45. Rausher MD (1981) Host plant selection by Battus philenor butterflies: the roles of predation, nutrition, and plant chemistry. Ecol. Mon. 51: 1-20. [ Links ]
46. Rothschild M (1984) Pyrazines as warning components in the Monarch butterfly, Danaus plexippus, and in moths of the genera Zygaena and Amata (Lepidoptera). Biol. J. Linn. Soc. 23: 375-380. [ Links ]
47. Rothschild M, Aplin RT, Cockrum PA, Edgard JA, Fairweather P, Lees R (1979) Pyrrolizidine alkaloids in arctiid moths (Lep.) with a discussion on host plant relationships and the role of these secondary plant substances in the Arctiidae. Biol. J. Linn. Soc. 12: 305-326. [ Links ]
48. Rothschild M, Nash RJ, Bell EA (1986) Cycasin in the endangered butterfly Eumaeus atala florida. Phytochemistry 25: 1853-1854. [ Links ]
49. Scott JA (1986) The Butterflies of North America. Stanford University. California. 583 pp. [ Links ]
50. Soto M, Giddings L, Gómez M (1996) Algunos usos de bioclimas: un sistema especializado de información geográfica. In Gormsen E, Jáuregui EO, Massey DB, Metcalfe SE, Santos M (Eds.) Investigaciones geográficas. Boletín del Instituto de Geografía. Número especial 4. Universidad Nacional Autónoma de México, México. pp. 63-83. [ Links ]
51. Steward JL, Keeler HK (1988) Are there trade-offs among antiherbivore defenses in Ipomoea (Convolvulaceae)? Oikos 53: 79-86. [ Links ]
52. Teas HJ (1967) Cycasin synthesis in Seirarctia echo (Lepidoptera) larvae fed methylazoxymethanol. Biochem. Biophys. Res. Comm. 26: 686-690. [ Links ]
53. Valenzuela-González J, López Méndez A, Lachaud JP (1995) Activity patterns and foraging activity in nests of Ectatomma tuberculatum (Hymenoptera: Formicidae) in cacao plantations. South. Entomol. 20: 505-515. [ Links ]
54. Vovides AP, Rees JD, Vázquez-Torres M (1983) Zamiaceae. In Gómez-Pompa A, Sosa V (Eds.) Flora de Veracruz, Fascículo 26. Instituto Nacional de Investigaciones sobre Recursos Bióticos. México. pp. 25-28. [ Links ]
55. Wagner JD, Wise DH (1996) Cannibalism regulates densities of young wolf spiders: evidence from field and laboratory experiments. Ecology 77: 639-652. [ Links ]
56. Whiting GM (1963) Toxicity of cycads. Econ. Bot. 17: 270-302. [ Links ]
57. Witz B (1990) Antidepredator mechanisms in arthropods: a twenty year literature survey. Fl. Entomol. 73: 71-79. [ Links ]
58. Yagi F, Tadera K, Kobayashi A (1980) Simultaneous determination of cycasin, methylazoxymethanol and formaldehyde by high performance liquid chromatography. Agr. Biol. Chem. 44: 1423-1425. [ Links ]
59. Zar JH (1999) Biostatistical Analysis. Prentice-Hall. New Jersey. 123 pp. [ Links ]
