











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.28 n.6 Caracas jun. 2003
ADAPTACIÓN AL MEDIO NATURAL DE BABAS (Caiman crocodilus) CRIADAS EN CAUTIVERIO
Andrés E. Seijas, Hilda Cuevas L. y Néstor González
Andrés E. Seijas. Licenciado en Biología, Universidad Central de Venezuela. M.Sc. y Ph.D., Universidad de Florida, EEUU. Profesor Asociado, Universidad Nacional Experimental de los Llanos Occidentales Ezequiel Zamora (UNELLEZ) Dirección: Vice-rectorado de Producción Agrícola. Antiguo Convento de San Francisco, carrera 3, Guanare, Portuguesa, Venezuela.e-mail: aeseijas@cantv.net
Hilda Cuevas L. Ingeniero en Recursos Naturales Renovables y M.Sc., UNELLEZ.
Néstor González. Ingeniero en Recursos Naturales Renovables, UNELLEZ.
Resumen
Se evaluó la adaptación al medio natural de babas (Caiman crocodilus) criadas en cautiverio liberadas antes del comienzo de la época seca (noviembre-diciembre 1992, 676 individuos) y a comienzos del período de lluvias (abril-julio 1993, 496 ejemplares) en cuerpos de agua del hato El Cedral, estado Apure, Venezuela. La recaptura de 210 individuos, entre 26 y 523 días después de su liberación, permitió determinar cambios en la gordura relativa, incidencia de heridas y tasas de crecimiento en su hábitat natural. Ciento quince (115) babas silvestres capturadas en el mismo hato se usaron como grupo control. Un análisis con un índice de gordura relativa (IGR) mostró que tanto las babas silvestres como las liberadas perdieron peso y crecieron a una tasa muy baja durante la época seca, comparado con la de aquellas liberadas al comienzo del periodo de lluvias. De 78 babas liberadas con cola completa (más de 16 crestas caudales sencillas), nueve (11,5%) perdieron parte de ésta, presumiblemente por interacciones intra e interespecíficas entre los 3 y 17 meses después de la liberación; esta cifra de individuos con cola rota es significativamente menor que la encontrada en babas silvestres de tallas similares, entre las cuales 22,8% tenían la cola rota. Los resultados indican una alta adaptabilidad al medio natural de las babas criadas en cautiverio y apoyan dicha actividad como estrategia para la recuperación de poblaciones sobre-explotadas o localmente extintas.
Summary
The adaptation to the natural environment of the spectacled caiman (Caiman crocodilus) raised in captivity and released before the dry season (november-december 1992, 676 individuals) and at the beginning of the rainy season (April-July 1993, 496 individuals) in water bodies of El Cedral ranch, Apure state, Venezuela, was evaluated. The recapture of 210 individuals, between 26 and 523 days after their release, was used to calculate changes in relative fatness, growth rates in their natural habitat, and prevalence of injuries. One hundred and fifteen (115) wild caimans captured in the study area were used as control group. Analyses with a relative fatness index showed that both wild and captive raised caimans lost weight during the dry season, and grew at a very low rate during the dry season, as compared to the growth rate of individuals released at the beginning of the rainy season. Nine out of 78 (11.5%) caimans released with complete tails (16 or more rows in the single crested caudal whorl) lost part of them, presumably due to inter and intraspecific interaction, between three and 17 months after their release. This percentage is lower than the one found in wild caimans of similar sizes, for which 22.8% had lost part of their tail. The results indicated a high adaptability of caimans raised in captivity to the natural environment, and support this procedure as a strategy for the recovery of overexploited or locally extinct populations.
Resumo
Avaliou-se a adaptação ao meio natural de filhotes de jacaré (Caiman crocodilus) criados em cativeiro liberados antes do começo da época seca (novembro-dezembro 1992, 676 indivíduos) e no início do período de chuvas (abril - julho 1993, 496 exemplares) em corpos de água do sitio El Cedral, estado Apure, Venezuela. A recaptura de 210 indivíduos, entre 26 y 523 dias depois de sua liberação, permitiu determinar mudanças na gordura relativa, incidência de feridas e taxas de crescimento em seu hábitat natural. Cento e quinze (115) filhotes de jacaré silvestres, capturados no mesmo sitio, usaram-se como grupo de controle. Uma análise com um índice de gordura relativa (IGR) mostrou que, tanto os filhotes de jacaré silvestres como os liberados, perderam peso e cresceram com uma taxa muito baixa durante a época seca comparado com a daqueles liberados no começo do período de chuvas. De 78 filhotes de jacaré liberados com cauda completa (mais de 16 crestas caudais simples), nove (11,5%) perderam parte desta, presumivelmente por interações intra e interespecíficas entre os 3 e 17 meses depois da liberação; esta cifra de indivíduos com cauda rompida é significativamente menor que a encontrada em filhotes de jacaré silvestres de tamanhos similares, entre as quais 22,8% tinham a cauda rompida. Os resultados indicam uma alta adaptabilidade ao meio natural dos filhotes de jacaré criados em cativeiro e apóiam dita atividade como estratégias para a recuperação de populações sobre-exploradas ou localmente extintas.
PALABRAS CLAVES / Caiman crocodilus / Manejo de Fauna / Zoocría /
Recibido: 14/03/2003. Modificado: 26/05/2003. Aceptado: 28/05/2003
Introducción
El levante y cría en cautiverio de crocodílidos (babas y caimanes) para su posterior liberación en el medio natural ha sido una estrategia de conservación varias veces propuesta y puesta en práctica en Venezuela (Blohm, 1973; Rivero-Blanco, 1974; Espinoza, 1988; Ramo et al., 1992; Arteaga, 1993; Lugo, 1998; Chávez, 2000; Muñoz y Thorbjarnarson, 2000; Seijas et al., 1990). Esta estrategia se fundamenta en el hecho de que en condiciones naturales todos los crocodílidos presentan una tasa de mortalidad mayor de 95% en los primeros meses de vida, etapa en que los recién nacidos son presas fáciles de un gran número de depredadores. El levante en condiciones controladas de individuos nacidos en cautiverio, o colectados en el medio natural en las primeras horas después de la eclosión, aumenta la probabilidad de supervivencia de éstos. Los ejemplares son mantenidos en los centros de levante durante un tiempo, generalmente un año, lo que les permite alcanzar una talla y peso adecuados para su liberación con tamaño suficiente para hacerlos poco vulnerables a gran parte de sus depredadores habituales.
El programa de aprovechamiento comercial de babas (Caiman crocodilus) iniciado en Venezuela en 1982, planteó la necesidad de establecer estrategias para mitigar el impacto que la extracción de individuos pudiera causar sobre las poblaciones naturales. En 1987 se inició un programa experimental de cría de babas (Gutiérrez, 1991) con los objetivos básicos de repoblar áreas naturales sujetas a explotación comercial y determinar la factibilidad económica de la cría de este crocodílido. Al implementarse la zoocría de babas con fines comerciales, se estableció en la normativa legal que cada zoocriadero debería liberar a la vida silvestre una cuota anual (2 al 10%) de los animales levantados en sus instalaciones, provenientes de neonatos o huevos colectados en el medio natural (Baquero y Quero, 1996). El presente artículo analiza y discute la adaptación al medio natural de babas criadas en cautiverio, en términos de crecimiento, condición física y desplazamiento, dentro del contexto de la normativa legal vigente para la fecha.
Área de Estudio
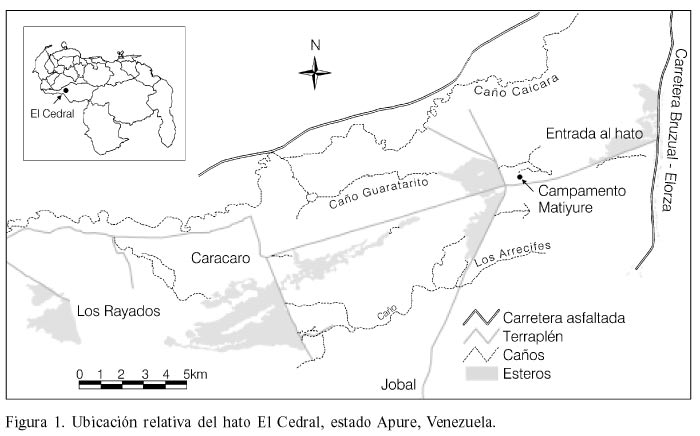
El estudio se realizó en el Hato El Cedral, estado Apure, Venezuela, ubicado entre los ríos Caicara y Matiyure, en la jurisdicción del Municipio Autónomo Muñoz, con una superficie de 53000ha (Figura 1). El área de estudio presenta la topografía plana típica del Alto Apure, con las formas fisiográficas denominadas por Ramia (1978) como bancos, bajíos y esteros.
El clima del área se caracteriza por presentar dos períodos definidos: uno de lluvias escasas, entre diciembre y marzo, y uno lluvioso, desde mayo hasta octubre. Los meses de abril y noviembre son de transición entre los periodos definidos. La precipitación anual es de unos 1600mm. La temperatura del área oscila entre 26,5 y 27,0ºC, con variaciones entre las medias mensuales inferiores a los 4ºC.
La investigación se realizó en las inmediaciones del área de administración del hato y campamento turístico Matiyure, y en las inmediaciones de la fundación Los Rayados, a unos 22km del punto anterior. En el hato existen diferentes tipos de cuerpos de agua tales como préstamos, caños y módulos. Los préstamos son excavaciones de profundidad y forma variable, generalmente rectangulares, construidos intencionalmente para retener agua durante el período seco o son subproductos del proceso de extracción de tierra para construcción de carreteras o diques que luego se llenan con el agua de lluvia o la escorrentía. Los caños son cauces estrechos, de bajo caudal, que pueden extenderse y ramificarse por la sabana y tienden a interrumpir el flujo de agua durante la sequía extrema. Los módulos, que son en esencia esteros artificiales, son extensiones de sabana que permanecen inundadas gran parte del año debido a la construcción de diques y terraplenes que interrumpen el flujo natural de las aguas durante el período de lluvias. Los módulos se caracterizan por poseer aguas relativamente profundas con abundancia de vegetación acuática, mientras que los préstamos son de muy variada forma y tamaño. Esta diferenciación de hábitat representa una simplificación y en cada una de esas categorías existe una amplia variedad de condiciones en términos de superficie, profundidad, vegetación acuática y permanencia de agua durante la época seca, lo cual seguramente incide en la disponibilidad de recursos alimentarios para las babas en cada uno de ellos.
Los cuerpos de agua donde se liberaron las babas fueron los caños Arrecife y Guaratarito; los módulos y préstamos a lo largo de los terraplenes vía El Jobal, Caicara y Caracaro, los cuales parten desde el Campamento Matiyure en direcciones sur, suroeste y noroeste, respectivamente; y los módulos y préstamos a lo largo del terraplén vía Los Rayados (Figura 1).
Materiales y Métodos
En noviembre 1992 existían en el zoocriadero de babas del Hato El Cedral unos 2000 ejemplares de aproximadamente 2 años de edad. De esta población se liberaron 676 individuos al medio natural al finalizar el periodo lluvioso de 1992 (noviembre-diciembre; Grupo 1) y 496 a comienzos del periodo de lluvias de 1993 (abril-julio; Grupo 2). Durante el desarrollo del estudio se capturaron 115 babas silvestres de tallas similares a las del zoocriadero, para tener un patrón de referencia al analizar la condición física de los individuos liberados recapturados (Tabla I).

Las hembras representaron 76,5% de las babas liberadas, no obstante para la mayoría de los análisis no se discriminó a los ejemplares por sexo. Todos los individuos fueron marcados con placas metálicas numeradas o se les practicó un marcaje por mutilación de escamas en la cresta caudal sencilla (CCS). A todas ellas se les midió la longitud total (LT), medida ventralmente desde la punta del hocico hasta el extremo de la cola; la longitud cabeza-cuerpo (LCC; Tabla II) desde la punta del hocico hasta el extremo posterior de la abertura cloacal; la longitud de la cabeza (Lcab) desde la punta del hocico hasta el borde posterior de la Tabla craneana; y el peso (P; Tabla II). Se registró además el número de filas en la CCS y el sexo. Las babas con menos de 16 CCS fueron consideradas como con cola rota.
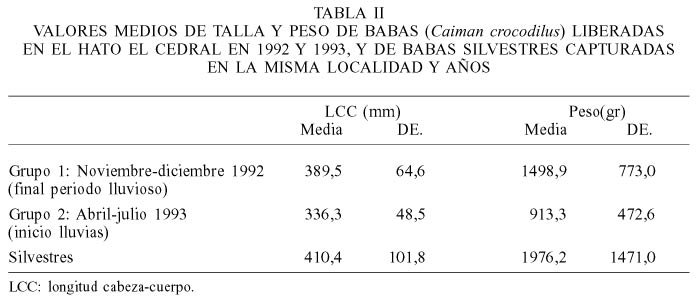
La comparación de las medidas o condición física de las babas al momento de la liberación con la que presentaban las babas silvestres o los mismos ejemplares cuando fueron recapturadas permitió evaluar sus adaptaciones al medio natural en términos de crecimiento (en LCC), gordura relativa, e incidencia de heridas. Tasas de crecimiento y gordura relativa relativamente altas, así como baja incidencia de heridas fueron consideradas como indicadores de adaptación al medio.
Para determinar si la probabilidad de supervivencia estaba relacionada con el tamaño de las babas al momento de la liberación, se comparó la estructura por clases de tamaño inicial (categorías de LCC) con la presentada por los animales recapturados después de 2 y 4 meses de su liberación. Las categorías de tamaño definidas para este propósito fueron: pequeñas (200-299mm LCC), medianas (300-399mm LCC) y grandes (>400mm LCC).
Para evaluar la condición física de los animales, se usó un índice de gordura relativa (IGR) expresado como
IGR = PR · PE-1 (1)
donde PR es peso real del individuo y PE es peso que se esperaría de una baba de talla (T) determinada.
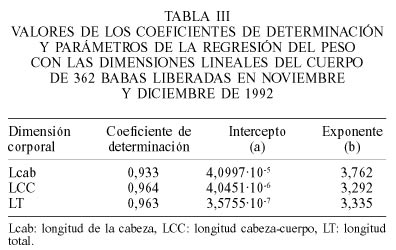
Para establecer el PE de cada individuo, se seleccionaron entre las babas liberadas en 1992, todas aquellas sin signos evidentes de enfermedad y sin mutilaciones. Con los datos de estos individuos se realizó un análisis de regresión lineal entre el logaritmo del peso y el logaritmo de cada una de las medidas corporales tomadas como indicadoras del tamaño (LT, LCC y Lcab). Dicho análisis permitió obtener la relación
PE = a · Tb (2)
donde PE es peso esperado y T es alguna de las medidas de tamaño corporal ya señaladas. Sustituyendo PE en la Ec. 1, se obtiene
IGR = PR (a · Tb)-1 (3)
donde IGR, PR y T son variables ya definidas; a y b son constantes obtenidas a partir del análisis de regresión, cuyos valores dependerán de las unidades de medida y de las dimensiones corporales utilizadas. En cualquier caso, el IGR está centrado en 1. Babas con IGR<1 son consideradas flacas y con IGR>1 son consideras gordas.
Para este estudio se consideraron como días de déficit hídrico (época seca) desde el lº de enero y hasta el 31 de mayo de cada año. Durante ese lapso los cuerpos de agua se reducen paulatinamente, llegando algunos a secarse por completo, lo que significa una reducción del hábitat y de los recursos en general para la baba. Los días restantes fueron considerados como de época húmeda, con abundancia de recursos para esta especie.
Se calcularon dos tipos de tasas de crecimiento: la tasa de crecimiento neta (TCN) y la tasa de crecimiento por época (seca o húmeda). La TCN se calculó dividiendo el incremento en LCC (cm) entre el tiempo (en meses) transcurrido entre la liberación y la recaptura. Los lapsos considerados para calcular la TCN incluyeron tanto días de época seca como de época húmeda.
Estudios previos han demostrado que las babas tienen un crecimiento muy lento durante la época seca (Gorzula, 1978; Thorbjarnarson, 1990). Para el cálculo del crecimiento en la época seca (Cs) se usaron solamente aquellos animales del Grupo 1 recapturados antes del comienzo del período lluvioso de 1993.
Debido a dificultades para la captura de babas durante los meses de lluvia, la tasa de crecimiento para la época húmeda (Ch) se determinó usando el modelo elaborado por Messel y Vorlicek (1989)
D
LCC = Ch · Dh + Cs · Ds (4)donde DLCC es el incremento en la longitud cabeza-cuerpo de la baba (LCC) en el lapso liberación-recaptura; Ch es la tasa de crecimiento durante la época húmeda; Cs es la tasa de crecimiento durante la época seca, calculada a partir de las recapturas de las babas liberadas temprano en la estación seca. Dh y Ds son los días de época húmeda y seca experimentados por el individuo en el lapso considerado.
De la Ec. 4, se despeja la tasa de crecimiento durante la época húmeda:
Ch = (DLCC - Cs · Ds) Dh-1 (5)
Para contrastar la hipótesis que las babas que ya hubieran experimentado la época seca previa en el medio natural crecerían más rápido durante la época húmeda subsiguiente que aquellas liberadas directamente al comienzo de la época húmeda, se usó un análisis de covarianza (ANCOVA) usando como covariable el lapso transcurrido entre liberación y recaptura.
Resultados
Para la determinación del índice de gordura relativa (IGR) se utilizó una muestra de 362 babas aparentemente sanas y sin mutilaciones de ningún tipo. La relación entre el peso y la LCC fue la que mostró el mejor coeficiente de determinación y la que produjo el exponente más cercano a 3 (valor para isometría perfecta) para la Ec. 2 (Tabla III, Figura 2). Por esta razón se usó sólo esta variable para los análisis del IGR.
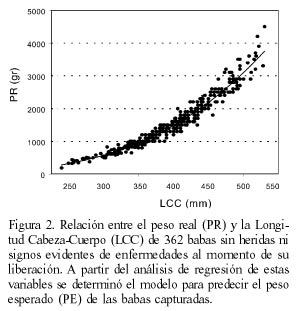
La ecuación final del IGR quedó como
IGR=PR(4,051·10-6·LCC3,292)-1 (6)
El IGR es independiente de la talla de los animales (Figura 3) y permite, por lo tanto comparar la gordura relativa de animales de distintos tamaños.
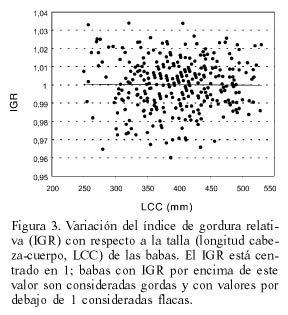
Las babas silvestres capturadas en noviembre y diciembre de 1992, en fechas cercanas a la de las babas liberadas del Grupo 1, presentaron un IGR mayor que las babas del criadero (IGR= 1,02; t= 6,89; P <0,0001).
Durante el transcurso del estudio se recapturaron 210 babas (17,9%) en períodos que van desde los 26 a los 523 días después de la fecha de liberación. Para algunos análisis se descartaron los datos de babas capturadas a menos de 60 días de su liberación o con cualquier dato que pusiera en duda la validez de la información vertida en las planillas de campo. Las 171 babas recapturadas después de los 60 días de la liberación representaron 14,6% de las liberadas.
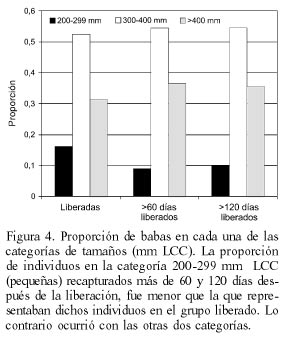
Al dividir las babas liberadas en tres categorías de tamaño (pequeñas, medianas y grandes), la proporción de individuos pequeños recapturados entre dos y cuatro meses después de la liberación, fue menor que la que ellos representaban entre las babas liberadas inicialmente. Las babas grandes, por el contrario, estuvieron representadas entre las babas recapturadas (luego de más de cuatro meses) en mayor proporción que el que presentaban al momento de la liberación (Figura 4). Las diferencias, sin embargo, no fueron significativas (Pearson X2= 8,304; P= 0,0811).
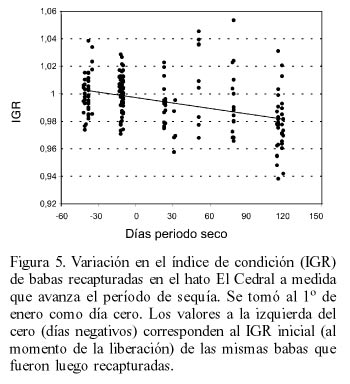
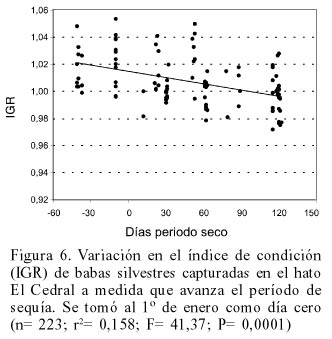
Las babas del Grupo 1 perdieron peso a medida que avanzaba la temporada de sequía (Figura 5). Esta pérdida de gordura relativa es similar a la experimentada por babas silvestres durante el mismo período (Figura 6) aunque el modelo de regresión explica una mayor proporción de la varianza en el caso de las babas silvestres (21%) con respecto a las babas liberadas (16%).
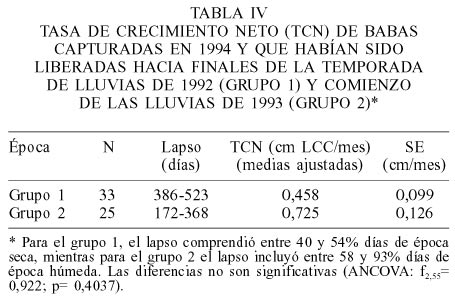
La tasa de crecimiento neto (TCN) de las babas del Grupo 1 capturadas en 1994, y que por lo tanto habían permanecido durante toda la época seca de 1993 en el medio natural, fue menor que las del Grupo 2, liberadas directamente al comienzo de la época húmeda. La diferencia, no obstante, no fue estadísticamente significativa (Tabla IV).
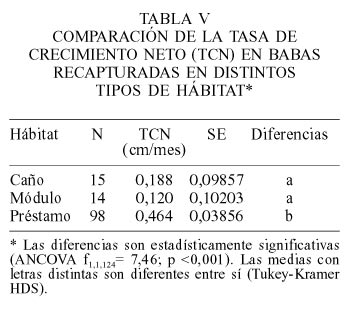
Las babas recapturadas en préstamos mostraron una TCN mayor que la exhibida por aquellas recapturadas en caños y módulos (Tabla V). Esas diferencias deben ser tomadas solamente como un indicio para profundizar en investigaciones posteriores, ya que no es posible saber cuanto tiempo, del lapso transcurrido entre liberación y recaptura, pasó cada individuo en alguna de las categorías de hábitat mencionadas.
El crecimiento de las babas durante el período seco (Cs) fue de 0,257 ±0,04cm/mes. Una alta proporción (17,7%) de ellas mostró incluso "crecimiento negativo". Este valor Cs se usó luego para estimar el crecimiento durante la época húmeda de acuerdo al modelo de Messel y Vorlicek (1989).
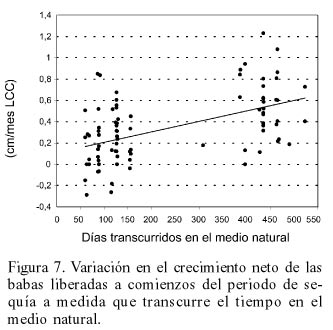
No obstante lo reducido del crecimiento durante la época seca, particularmente en los días inmediatos después de la liberación, la mayoría de las babas mostraron un incremento neto en sus tallas al transcurrir más de un año, cuando ya habían experimentado un periodo de lluvias en el medio natural (Figura 7).
La tasa de crecimiento promedio durante la época húmeda (Ch) de las babas que ya habían experimentado un período seco en el medio natural (0,752cm/mes) fue similar al de babas liberadas directamente al comienzo de dicho período (0,739cm/mes). Esto sugiere que los meses de adaptación previos no constituyeron una ventaja apreciable en la adaptación de las babas a su medio natural.
Los machos mostraron un Ch= 1,052cm/mes, mayor que el de las hembras (0,724cm/mes) pero debido a la amplia variación de los valores las diferencias no fueron estadísticamente significativas (t= 1,571; P= 0,1221). Basado en este análisis y en el hecho de que la muestra estuvo muy sesgada hacia las hembras, en los análisis siguientes no se discriminó a las babas por sexo.
De un total de 78 babas que fueron liberadas con la cola completa (>15 CCS), nueve (11,5%) perdieron parte de ésta entre los tres y 17 meses después de la liberación. Dichas mutilaciones ocurrieron probablemente como consecuencia de interacciones intra o interespecíficas. Dicha cifra es significativamente menor (X2= 3,9; P <0,05) que la encontrada en babas silvestres de tallas similares, entre las cuales 22,8% tenían la cola rota (menos de 16 filas en la CCS).
Discusión
Aún cuando la liberación de babas criadas en zoocriaderos en Venezuela se llevó a cabo en oportunidades previas a la reseñada en el presente estudio (Espinoza, 1989; Baquero y Quero, 1996) este representa el primer esfuerzo de evaluar el éxito de esta estrategia de manejo.
El porcentaje de babas recapturadas durante el estudio (14,6%) representa una estimación mínima de la supervivencia en el medio natural de los individuos liberados. Muchas de las babas se soltaron en localidades donde la recaptura era difícil. Otras pudieron haberse movilizado hacia lugares no muestreados y por lo tanto, fuera del alcance de los investigadores. En todo caso, la proporción de recapturas es una subestimación de la supervivencia de las babas liberadas y excede con mucho la tasa de supervivencia de crías de crocodílidos en el medio natural, que probablemente no supere el 5%.
La talla inicial de las babas liberadas pudiera jugar un papel importante en la probabilidad de supervivencia en el medio natural. Los resultados indican que las babas más grandes (mayores de 400mm LCC) aparecieron en las recapturas en mayor proporción que la esperada de acuerdo a su representación entre los animales liberados. Aunque la prueba estadística no apoyó de manera contundente esta hipótesis, la probabilidad asociada al valor de X2 (P= 0,0811) es lo suficiente bajo como para recomendar que se liberen animales de mayores tallas en ensayos futuros.
La pérdida de peso de las babas liberadas durante los días de la época seca se asemeja a la experimentada por babas silvestres en la misma localidad, lo que indica que se trata de un fenómeno natural consecuencia del estrés provocado por la escasez de agua y, probablemente, de alimento durante dicha estación. La mayor dispersión en la caída del IGR de las babas recapturadas puede ser interpretada como una consecuencia de las dificultades de estos animales en ubicarse, durante el tiempo transcurrido desde la liberación, en las condiciones de hábitat más favorables, cosa que las babas silvestres, con mucho más tiempo de aprendizaje, debieron haber logrado.
El estrés anteriormente señalado es lo que provocaría la baja tasa de crecimiento durante la época seca. Las tasas de crecimiento calculadas en este estudio durante la estación seca son superiores a las encontradas por Thorbjarnarson (1990) en otra localidad de los llanos de Venezuela. Dicho autor reportó tasas negativas para babas en las categorías de 30-50cm LCC, es decir, similares a las de este estudio. Thorbjarnarson (1990) explica que las tasas de crecimiento negativas probablemente se deban a una compactación de los elementos esqueléticos en la columna vertebral.
Por otra parte, las tasas de crecimiento durante la época húmeda calculadas en este estudio son inferiores a los reportados por Thorbjarnarson (1990), quien reportó valores entre 0,032 y 0,039cm/día (equivalentes a 0,973 y 1,186cm/mes). Una posible explicación a esta discrepancia es que las babas liberadas en el Hato El Cedral todavía no habrían alcanzado una adaptación plena a las nuevas condiciones y algunas todavía no habrían logrado seleccionar las condiciones de hábitat óptimas para su pleno desarrollo. Esto es apoyado por las diferencias que se encontraron en las tasas de crecimiento de babas capturadas en los distintos cuerpos de agua, donde aquellas localizadas en los préstamos fueron las que obtuvieron las tasas de crecimiento mayores. Esta conclusión debe ser tomada con cuidado pues no hubo realmente un control sobre el tipo de hábitat en que las babas se mantuvieron durante la realización del estudio.
Debido a la dificultad de recapturar animales durante la época húmeda, las tasas de crecimiento durante ese lapso se estimaron usando el modelo de Messel y Vorlicek (1989). Se esperaba que las babas liberadas al comienzo de la época seca, y que ya habían experimentado un largo período (más de cinco meses) en el medio natural, crecieran más rápido durante la época húmeda que babas liberadas directamente al comienzo de dicho periodo. El caso es que ese factor no parece ejercer ningún efecto sobre el crecimiento de las babas liberadas. Es recomendable, por lo tanto, liberar los animales directamente durante el periodo lluvioso, evitándole, en sus etapas iniciales el estrés que significa la época seca.
Otro elemento con el que se puede medir la adaptación de los animales liberados, es la incidencia de heridas. Las babas introducidas al medio natural mostraron una incidencia de mutilaciones de la cola menor que las silvestres de tallas similares. No obstante, hay que tener en cuenta que las babas silvestres con que se compararon las babas liberadas seguramente habían permanecido en su medio ambiente por un lapso mucho mayor que el máximo de 17 meses que las babas provenientes del zoocriadero. Otra posible explicación de esta diferencia es que las babas silvestres son más propensas a heridas y mutilaciones en las primeras etapas de su vida, etapa que no experimentaron las babas del zoocriadero, que fueron liberadas con tallas promedio superiores a los 30cm LCC, talla que las babas silvestres alcanzan, tomando en cuenta los datos disponibles en la literatura (Gorzula, 1978; Ayarzagüena, 1983; Thorbjarnarson, 1990) al menos luego de los dos años de vida.
En términos generales se puede concluir que las babas criadas en cautiverio mostraron signos de adaptación al medio natural, en términos de supervivencia, crecimiento e incidencia de heridas. Esto permite recomendar a la cría en cautiverio como estrategia para restaurar poblaciones que hayan sido explotadas en extremo. Este no era el caso del Hato El Cedral para el momento de realizar el estudio. Aunque los zoocriaderos de babas de Venezuela dejaron de funcionar a mediados de los años 90 del siglo pasado, si programas de aprovechamiento comercial similares (con esa especie u otras especies de crocodílidos) llegaran a desarrollarse en el futuro, sería conveniente mantener la provisión de reservar una pequeña cuota de los animales del zoocriadero para programas de restauración dentro o fuera del área donde se encuentre el zoocriadero. La utilización o no de los animales reservados para dichos fines dependería de las condiciones del momento. Si los programas de aprovechamiento de poblaciones silvestres de babas mantienen tasas de extracción adecuadas, a niveles determinados en estudios técnicos bien conducidos, programas de restauración poblacional como el discutido en este trabajo quizás no serían necesarios de implementar.
Agradecimientos
Este estudio contó con el apoyo económico de la Asociación de Criadores de Baba (ASOBABA, Manuel Denís), PROFAUNA (MARN, Mirna Quero) y la UNELLEZ. El hato El Cedral prestó facilidades de alojamiento durante el trabajo de campo. Los autores agradecen en particular a Tulio Aguilera y Saúl Gutiérrez (hato El Cedral). Carlos Chávez, Manuel González, Gloria Duno, Carlos Erazo, María Muñoz, Jesús Rivas y John Thorbjarnarson, entre otros, colaboraron en la recolección de información de campo. Juan E. García asesoró en el proceso de interpretación de los datos en una fase inicial de este estudio.
Referencias
1. Arteaga A (1993) Repoblamiento del embalse de Tacarigua (Edo Falcón, Venezuela) con caimanes de la costa (Crocodylus acutus) criados de cautiverio. En Zoocría de los Crocodylia. Memorias I Reunión Regional CSG/UICN/World Conservation Union. Gland, Suiza. pp. 263-269. [ Links ]
2. Ayarzagüena J (1983) Ecología del caimán de anteojos o baba (Caiman crocodilus L.) en los llanos de Apure (Venezuela). Doñana. Acta Vertebrata 10: 45-117. [ Links ]
3. Baquero B, Quero M (1996) Manejo del programa de zoocriaderos de la especie baba (Caimán crocodilus) en Venezuela. Zoocriaderos 1: 1-6. [ Links ]
4. Blohm T (1973) Conveniencia de criar cocodrilos en Venezuela con fines económicos y para prevenir su extinción. 1er Simposio Internacional sobre la Fauna Silvestre y Lacustre Amazónica. Manaus, Brasil. pp. 1-30. [ Links ]
5. Chávez C (2000) Conservación de las poblaciones del caimán del Orinoco (Crocodylus intermedius) en Venezuela. Informe Profauna-Corpoven. MARN, Caracas, Venezuela. 110 pp. [ Links ]
6. Espinoza E (1988) Desarrollo de criaderos de babas Caiman crocodilus en los llanos venezolanos. II Simposio Nacional sobre Biotecnología. Maracaibo, Venezuela. pp. 127-134. [ Links ]
7. Espinoza E (1989) La cría de la baba Caiman crocodilus en Venezuela. Estado actual y futuro. II Taller de Conservación y Manejo de la Baba Caiman crocodilus en Venezuela. Guanare, Venezuela. 13 pp. [ Links ]
8. Gorzula S (1978) An ecological study of Caiman crocodilus crocodilus inhabiting savannah lagoons in the Venezuelan Guayana. Oecologia 35: 21-34. [ Links ]
9. Gutiérrez, S (1991) Situación actual de los zoocriaderos de baba en Venezuela. III Taller sobre Conservación y Manejo de la Especie baba (Caiman crocodilus). Caracas, Venezuela. 10 pp. [ Links ]
10. Lugo M (1998) Evaluación del programa de liberación del caimán del Orinoco (Crocodylus intermedius) en el Refugio de Fauna Silvestre "Caño Guaritico" y alrededores. Estado Apure. Tesis. UNELLEZ. Guanare, Venezuela. 104 pp. [ Links ]
11. Messel H, Vorlicek GC (1989) Status and conservation of Crocodylus porosus in Australia. En Crocodiles. Their Ecology, Management, and Conservation. IUCN. Gland, Suiza. pp. 110-137. [ Links ]
12. Muñoz M, Thorbjarnarson J (2000) Movement of captive-released Orinoco crocodiles (Crocodylus intermedius) in the Capanaparo River, Venezuela. J. Herpetol. 34: 397-403. [ Links ]
13. Ramia M (1978) Tipos de sabana de los llanos de Venezuela. Bol. Soc. Venez. Cienc. Nat. 112: 264-287. [ Links ]
14. Ramo C, Busto B, Utrera A (1992) Breeding and Rearing Orinoco crocodile (Crocodylus intermedius) in Venezuela. Biol. Cons. 60: 101-108. [ Links ]
15. Rivero-Blanco C (1974) Hábitos reproductivos de la baba en los llanos venezolanos. Natura 52: 24-29. [ Links ]
16. Seijas AE, Chang A, Cordero DG (1990) Cría de Caimanes de la costa (Crocodylus acutus) con fines de repoblamiento. Biollania 7: 13-26. [ Links ]
17. Thorbjarnarson J (1990) Ecology and behavior of the spectacled caiman (Caiman crocodilus) in the central Venezuelan llanos. Dissertation. University of Florida, Gainesville. EEUU. 268 pp. [ Links ]
