











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.30 n.9 Caracas sep. 2005
COMPARACIÓN DE LA TOLERANCIA A LA SEQUÍA DE CUATRO VARIEDADES DE MAÍZ (Zea mays L.) Y SU RELACIÓN CON LA ACUMULACIÓN DE PROLINA
Carlos Hugo Avendaño Arrazate, Carlos Trejo López, Cándido López Castañeda, José Domingo Molina Galán, Amalio Santacruz Varela y Fernando Castillo González
Carlos Hugo Avendaño Arrazate. Ingeniero Agrónomo, Universidad Autónoma Chapingo (UACh), México. Maestría y Estudiante de Doctorado en Genética, Colegio de Postgraduados (COLPOST), México. Dirección: IREGEP, Colegio de Postgraduados. Km. 36.5 Carretera México-Texcoco, CP 56180, Montecillo, Edo. de México. México.e-mail: acarlos@colpos.mx
Carlos Trejo López. Biólogo, Universidad Nacional Autónoma de México. Maestría, COLPOST, México. Ph.D., University of Lancaster, Inglaterra. Profesor Investigador, COLPOST, México.
Cándido López Castañeda. Ingeniero Agrónomo, UACh, México. Maestría, COLPOST, México. Ph.D., Australian National University. Profesor Investigador, COLPOST, México.
José Domingo Molina Galán. Ingeniero Agrónomo, UACh, México. Maestría, COLPOST, México. Ph.D., North Carolina State University, EEUU. Profesor Investigador, COLPOST, México.
Amalio Santacruz Varela. Ingeniero Agrónomo, UACh, México. Maestría, COLPOST, México. Ph.D., Iowa State University, EEUU. Profesor Investigador, COLPOST, México.
Fernando Castillo González. Ingeniero Agrónomo, UACh, México. Maestría, COLPOST, México. Ph.D., North Carolina State University, EEUU. Profesor Investigador, Colegio de COLPOST, México.
Resumen
Se comparó la tolerancia a la sequía con base en los potenciales hídricos, acumulación de prolina y biomasa, en las variedades de maíz Zacatecas 58 original (Zo), Cafime original (Co) y sus respectivas variedades mejoradas para resistencia a la sequía, Zacatecas 58 Selección Masal 19 (Z19) y Cafime Selección Masal 16 (C16) en condiciones de sequía y riego en invernadero. En la quinta hoja de las cuatro variedades los potenciales hídrico (Yw) y osmótico (Yp) disminuyeron conforme se prolongó la sequía; las variedades mejoradas presentaron menores Yw y Yp con respecto a las originales. La acumulación de prolina aumentó conforme se prolongó el periodo de sequía en las cuatro variedades, en diferente proporción. Las variedades mejoradas presentaron mayor acumulación de prolina que las originales, con excepción de la Z19, que a los 38 días de sequía mostró menor acumulación que Zo y mayor acumulación de biomasa. Los resultados permiten establecer que las variedades mejoradas por selección masal en condiciones de humedad restringida en el suelo, han desarrollado un mecanismo de resistencia a sequía semejante al propuesto inicialmente por otros autores, al cual han llamado latencia o resurrección, que les permite sobrevivir en condiciones extremas de sequía, recuperarse completamente y reiniciar su metabolismo normal y crecimiento una vez que hay humedad, donde la prolina juega un papel preponderante.
Summary
Drought tolerance was evaluated in relation to proline accumulation, hydric relations and biomass of the original varieties Zacatecas 58 (Zo) and Cafime (Co), as well as of their drought resistant improved versions, Zacatecas 58 SM19 (Z19) and Cafime SM16 (C16), under drought and irrigation conditions in a greenhouse. On the fifth leaf of all four varieties the water potential (Yw) and osmotic potential (Yp) diminished as the drought lengthened; the improved varieties showed lower Yw and Yp than the original ones. Proline accumulation increased in different proportions in the four varieties as the drought period lengthened. The improved varieties showed more proline accumulation than the originals, except for Z19 at 38 days of drought, when they showed less proline accumulation and more biomass yield than Zo. The results indicate that the improved varieties through mass selection under soil water restriction have developed a drought resistant mechanism, similar to the one previously proposed, known as latency or resurrection, which helps plants survival in extreme drought conditions, reviving entirely and recuperating their normal metabolism and growth after watering. Proline might be playing an important role in this mechanism.
Resumo
Comparou-se a tolerância à seca com base nos potenciais hídricos, acumulação de prolina e biomassa, nas variedades de milho "Zacatecas 58" original (Zo), "Cafime" original (Co) e suas respectivas variedades melhoradas para resistência a seca, "Zacatecas 58" Seleção Massal 19 (Z19) e "Cafime" Seleção Massal 16 (C16) em condições de seca e irrigado em estufa. Na quinta folha das quatro variedades os potenciais hídrico (Yw) e osmótico (Yp) diminuíram conforme se prolongou a seca; as variedades melhoradas apresentaram menores Yw y Yp em relação às originais. A acumulação de prolina aumentou conforme se prolongou o período de seca nas quatro variedades, em diferente proporção. As variedades melhoradas apresentaram maior acumulação de prolina que as originais, com exceção da Z19, que aos 38 dias de seca mostrou menor acumulação que Zo e maior acumulação de biomassa. Os resultados permitem estabelecer que as variedades melhoradas por seleção massal em condições de umidade restringida no solo, tem desenvolvido um mecanismo de resistência à seca semelhante ao proposto inicialmente por outros autores, ao qual tem chamado latência ou ressurreição, que lhes permite sobreviver em condições extremas de seca, recuperar-se completamente e reiniciar seu metabolismo normal e crescimento uma vez que existe umidade, onde a prolina tem um papel preponderante.
PALABRAS CLAVE / Potencial Hídrico / Prolina / Sequía / Zea mays L. /
Recibido: 20/08/2004. Modificado: 22/06/2005. Aceptado: 20/07/2005.
Introducción
En México el maíz es el cultivo más importante en términos de consumo humano; sin embargo, aproximadamente el 50% de la producción se lleva a cabo en zonas de temporal, donde la lluvia es escasa y su distribución aleatoria (Muñoz, 1980), lo que provoca una reducción importante en el rendimiento. La mayoría de los trabajos de mejoramiento genético realizados en maíz en condiciones de sequía han sido enfocados principalmente al estudio de los componentes del rendimiento, bajo el supuesto de que la selección masal visual estratificada en condiciones de humedad restringida del suelo, incrementa la resistencia a la misma (Molina, 1983). En estos estudios se ha podido observar que la sequía provoca el aborto de los sacos embrionarios e induce una asincronía entre la antesis y la floración femenina (Desai y Singh, 2001). El déficit hídrico en las plantas provoca toda una serie de respuestas morfológicas, fisiológicas y fenológicas, las cuales pueden tener algún valor adaptativo y por lo tanto conferir un cierto grado de tolerancia a esta condición de estrés (Dreesmann et al., 1994; Attipalli et al., 2004). En maíz, la sequía reduce el rendimiento de grano como consecuencia del menor número de granos por mazorca y peso del mismo (Stone et al., 2001).
Durante el estrés hídrico en los tejidos de las plantas se genera una acumulación activa de solutos como respuesta de sobrevivencia (Attipalli et al., 2004). Uno de los principales solutos registrados durante este estrés es la prolina, cuya función ha sido asociada a sostener la turgencia de los tejidos para mantener la función celular (Ramanjulu y Sudhakar, 2000). Se ha sugerido que la acumulación de prolina en hojas estresadas por sequía, actúa como agente osmótico durante el estrés, protegiendo a la planta contra la desecación (Harsh, 2003) y actuando como almacén de nitrógeno que será utilizado durante la rehidratación (Stewart y Hanson, 1980; Ramanjulu y Sudhakar, 2000). Se ha observado que el aumento de la concentración de prolina en maíz se lleva a cabo en gran medida en la raíz primaria, sobre todo cuando crece en substratos con bajos potenciales hídricos, incrementando su elongación y favoreciendo una mayor exploración (Petcu y Terbea, 1996; Bajji et al., 2000); por ello, altas concentraciones de prolina en plantas bajo estrés hídrico han sido consideradas como un criterio de selección de materiales tolerantes a la sequía (Ober y Sharp, 1994; Verslues y Sharp, 1999). En los últimos 20 años, en el Colegio de Postgraduados, México; se ha desarrollado un programa de mejoramiento genético en maíz (Molina, 1980), con el fin de obtener genotipos tolerantes a la sequía mediante la selección masal. Este programa ha generado las variedades Zacatecas 58 y Cafime, las cuales durante 19 y 16 ciclos de selección masal respectivamente, han mostrado cierta tolerancia a la sequía y adaptación a las bajas precipitaciones de las áreas productoras de maíz ubicadas en valles altos de México; lo cual representa una valiosa alternativa para atenuar los bajos rendimientos de maíz en dichas áreas. El objetivo del presente trabajo, fue comparar la tolerancia a la sequía con base en potenciales hídricos, acumulación de prolina y rendimiento de biomasa en cuatro variedades de maíz.
Materiales y Métodos
El trabajo fue realizado en 2003, en condiciones de invernadero en el Colegio de Postgraduados, México; ubicado en valles altos entre 19º29'N y 98º53'O y a 2250msnm. El clima es del tipo Cb(wo)(w)(i')g, correspondiente a templado con verano fresco largo, temperatura media anual entre 12 y 18ºC. El mes más frío oscila entre -3 y 18ºC, y el más cálido entre 6,5 y 22ºC. La precipitación media anual es de 637mm (García, 1988).
Como material biológico se utilizaron las variedades de maíz originales Zacatecas 58 (Zo) y Cafime (Co), y sus respectivas variedades mejoradas, denominadas Zacatecas 58 selección masal 19 (Z19) y Cafime selección masal 16 (C16), las cuales fueron obtenidas bajo el método de selección masal visual estratificada en suelos con muy baja humedad (Molina, 1983).
Se utilizó un diseño de bloques completamente al azar con tres repeticiones en un arreglo factorial 5×4, correspondiendo a cuatro tratamientos de sequía y uno de riego en cada una de las cuatro variedades. Los tratamientos fueron: riego (R) o sequía (S), pudiendo esta última mantenerse por 4, 10, 20 ó 38 días (S1, S2, S3 y S4, respectivamente). Los tratamientos de sequía se iniciaron a partir de que el suelo llegó a punto de marchitez permanente (PMP; S0), y el tratamiento de riego consistió en mantener el suelo por arriba del 60% de humedad aprovechable durante todo el ciclo, equivalente a 140 días. A todos los tratamientos de sequía se les aplicó un riego de recuperación (RR) una vez alcanzado el periodo determinado de días de sequía a partir del PMP, para continuar con riego normal durante el resto del ciclo. La unidad experimental en cada tratamiento estuvo constituida por dos macetas, cada una con dos plantas haciendo un total de 192 macetas (96 en sequía y 96 en riego). Se utilizaron como macetas, tubos de PVC de 50cm de altura y 10cm de diámetro con tapa y perforaciones en el fondo. Cada maceta contuvo 4,25kg de suelo de textura migajón-arcillo-arenoso con un contenido hídrico a capacidad de campo (CC) de 24,3% y PMP de 12,2% de la curva de retención de humedad. Las plantas crecieron en el invernadero 140 días a temperatura máxima promedio de 42ºC y mínima promedio de 9ºC. Cada maceta recibió una fertilización 15 días después de la siembra con la fórmula NPK 40-30-20, usando 27g de sulfato de amonio al 20,5%, 21g de superfosfato de Ca simple al 20% y 4,5g de cloruro de potasio al 60%.
Las variables evaluadas fueron los potenciales hídrico (Yw) y osmótico (Yp), y acumulación de prolina en la quinta hoja completamente expandida de las dos plantas de cada maceta. El Yw fue determinado con una bomba de presión Scholander a las 11:00am, y el Yp con un osmómetro de presión de vapor (VAPRO/WESCOR mod. 5200); ambas variables se registraron por primera vez cuando el suelo alcanzó el PMP (29 días después del último riego y 41 días acumulados desde la siembra). Posteriormente las determinaciones se realizaron de acuerdo al periodo de sequía para cada tratamiento, de tal forma que para S1 correspondió a 0 y 4 días, para S2 a 6, 8 y 10 días, mientras que para S3 y S4, debido a que el grado de marchitamiento fue tan severo no fue posible hacer mediciones. La acumulación de prolina fue determinada por el método colorimétrico desarrollado por Bates et al. (1973). Las determinaciones fueron hechas tomando 4cm2 de las hojas de cada tratamiento. El primer muestreo se hizo cuando el suelo alcanzó el PMP y después, de acuerdo al periodo de sequía para cada tratamiento. Para S1 correspondió a los 4 días después de PMP y 24h después del riego de recuperación (RR1). Para S2, a los 10 días después de PMP y 24h después de RR (RR2). Para S3 y S4 a los 20 y 38 días después de PMP y 24h después del RR (RR3 y RR4). Al final del ciclo de cada tratamiento se estimó la biomasa aérea total. El efecto de tratamiento fue determinado mediante el análisis de varianza de acuerdo con el procedimiento del modelo lineal general. La comparación entre medias de variedades se hizo mediante el método de diferencia mínima significativa (DMS, p £0,05).
Resultados y Discusión
Potenciales hídricos
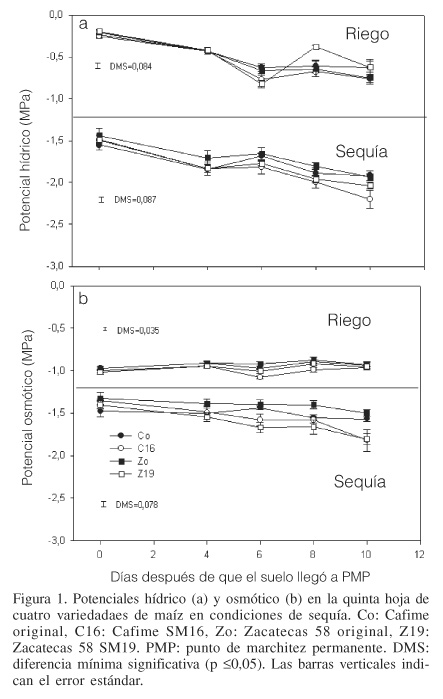
El potencial de agua (Yw) no mostró diferencias significativas en ninguna de las variedades para el tratamiento de riego (R). Sin embargo, en los tratamientos de sequía se observó una ligera tendencia a la disminución del Yw conforme el periodo de sequía S1 y S2 y la edad fenológica de los materiales avanzó. La diferencia mínima significativa (DMS p £0,05) para los tratamientos de sequía entre variedades fue de 0,087 (Figura 1a), lo que confirma la diferencia entre las variedades originales y las mejoradas. La disminución real del Yw fue en promedio -1,5MPa en las cuatro variedades cuando el suelo alcanzó el PMP, y posteriormente, a los 4, 6, 8 y 10 días la disminución fue gradual hasta alcanzar su valor promedio más bajo de -2,04MPa.
Los valores de potencial osmótico (Yp) registrados durante los periodos de sequía S1 y S2 presentaron poca variación significativa entre variedades, lo cual se reflejó en su DMS p £0,05 de 0,078 (Figura 1b). Para los tratamientos S3 y S4, debido a que el grado de marchitamiento fue muy severo no fue posible obtener valores. El Yp no presentó mucha variación durante los primeros cuatro días (S1) para las cuatro variedades. Sin embargo, a partir del día seis, las variedades mejoradas disminuyeron ligeramente su Yp, alcanzando su valor más bajo a los diez días. Es probable que a pesar de ser muy ligera, la variación del Yp haya sido suficiente para que los tejidos perdieran la turgencia. Lo anterior podría explicar porqué en las plantas bajo los tratamientos S3 y S4 no fue posible obtener valores de Yw y Yp, debido a extrema marchitez. Aún cuando la variación de los potenciales registrados en las cuatro variedades no fue muy significativa, sugiere que las variedades mejoradas responden con mayor rapidez que las originales a cambios ligeros en el gradiente del potencial hídrico del suelo y el potencial de agua de la raíz.
Acumulación de prolina
La acumulación de prolina en el tratamiento de riego mantuvo un valor promedio de 1,48nmol en las cuatro variedades sin diferencia estadística entre ellas (Figura 2). Por el contrario, en los tratamientos de sequía se observó una acumulación de aproximadamente cuatro veces, en comparación con el tratamiento de riego. En la variedad mejorada C16, la acumulación de prolina entre las plantas bajo riego y sequía se incrementó hasta 20 veces conforme el periodo de sequía alcanzó los 38 días (S4; Figura 2e). La acumulación de este aminoácido y de otros como valina, isoleucina, leucina, ácido glutámico, ácido aspártico y treonina, es considerada como una respuesta de las plantas al estrés hídrico (Girousse et al., 1996; Van Heerden y Kruger, 2002).
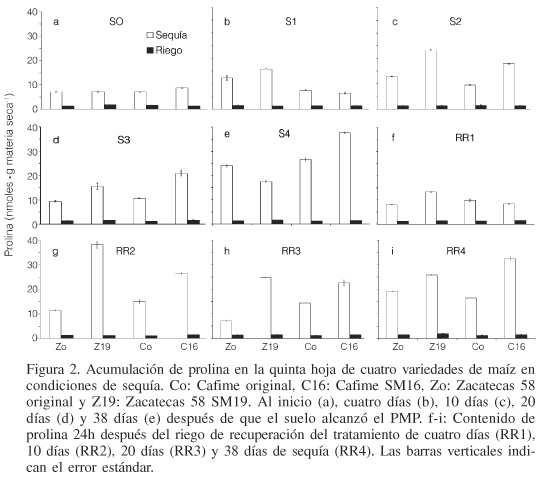
La acumulación de prolina por gramo de materia seca, entre las variedades y tratamientos de riego y sequía fueron altamente significativas. En riego la acumulación promedio fue de 1,48nmol·g-1, mientras que para sequía fue de 16,43nmol·g-1 (Figura 2). En Zacatecas 58, la acumulación de prolina fue mayor en la variedad mejorada Z19 que en la original Zo a medida que la sequía fue más severa (Figuras 2a-d), con excepción del tratamiento S4 donde Zo fue mayor que Z19 (Figura 2e). En Cafime, la acumulación de prolina fue mayor en la variedad mejorada C16 que en la original Co, conforme se prolongó la sequía (Figuras 2a-e). Una mayor acumulación de prolina en las variedades mejoradas de maíz para resistencia a sequía, con respecto a las no mejoradas, también fue observada por Carceller et al. (1999). Los casos en que la variedad original acumuló más prolina que la variedad mejorada, como ocurrió en el tratamiento S1 para Cafime (Figura 2b) y en el S4 para Zacatecas 58 (Figura 2e), también fueron observadas en maíz por Ilahi y Dorffling (1982), y en la propia variedad Cafime con siete ciclos de selección para resistencia a sequía por Ibarra-Caballero et al. (1988). Similar respuesta ha sido observada en maíz y otras especies por Ober y Sharp (1994), Bogoslavsky y Neumann (1998), Verslues y Sharp (1999), y Wassom et al. (2000).
La acumulación de prolina en las cuatro variedades durante la sequía no mostró un incremento constante, lo cual podría atribuirse al hecho de que durante el periodo de estrés hídrico la prolina es degradada para inhibir su oxidación (Ramanjulu y Sudhakar, 2000), lo cual habría impactado en las concentraciones registradas (Figura 2).
Después del riego de recuperación (RR) la acumulación de prolina fue mayor en las variedades mejoradas Z19 y C16 que en sus respectivas variedades originales Zo y Co (Figura 2f-i). Al comparar en las cuatro variedades, la cantidad de prolina en la condición de sequía con la retenida un día después del RR, se observa que Zo acumuló más prolina durante la sequía que después del RR y lo contrario sucedió con Z19, con excepción del tratamiento S1. Este comportamiento probablemente se debió a que el mecanismo de síntesis de prolina ya estaba activado y aun después del RR las plantas siguieron biosintetizando prolina para tener una mayor recuperación. En este contexto, Wassom et al. (2000) encontraron que la acumulación de prolina en hojas estresadas está asociada positivamente con la capacidad de recuperación y señalan que, posiblemente, la prolina sirve como fuente de energía respiratoria para que la planta pueda recuperarse. De la misma forma, Neale et al. (2000) encontraron que la recuperación de las plantas bajo severo estrés hídrico está asociada a la expresión de genes específicos 24h después del riego de recuperación. Estos autores han denominado a este fenómeno como resurrección.
Biomasa total
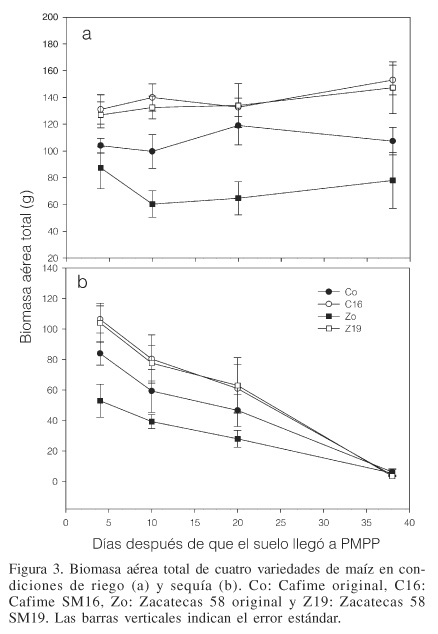
Los resultados obtenidos para biomasa aérea de las cuatro variedades mostraron que ésta se redujo gradualmente conforme el periodo de sequía fue más prolongado (Figura 3), lo cual podría ser atribuido al estrés hídrico. Las variedades mejoradas Z19 y C16 disminuyeron en menor grado su biomasa que sus respectivas variedades originales Zo y Co. Si se relaciona la reducción de biomasa con la acumulación de prolina, es posible sugerir que a mayor acumulación de prolina en los tejidos, menor reducción de biomasa. En la Figura 4 se observa que las variedades mejoradas presentaron una relación lineal menos negativa que las variedades mejoradas. Estos resultados pueden tomarse en cuenta para hacer planteamientos más específicos y probar la hipótesis, al menos para el caso de estas variedades, de que una mayor concentración de prolina en los tejidos corresponde a menor abatimiento de la biomasa.
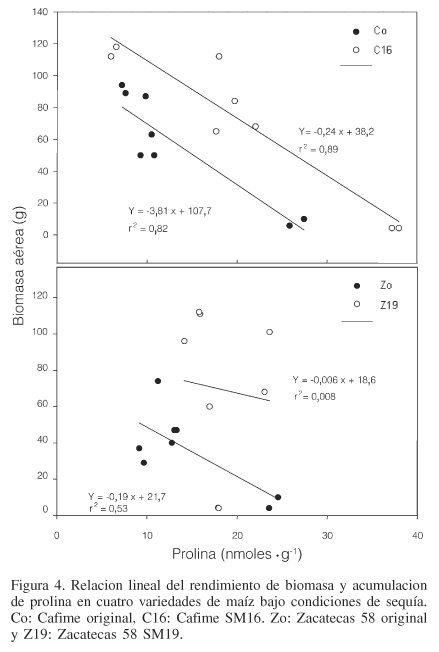
Los resultados obtenidos con la prolina sugieren que posiblemente este aminoácido esta relacionado con el mantenimiento de las funciones fisiológicas en el citoplasma de las células aun cuando éstas hayan sido deshidratadas severamente, lo cual le confiere un mecanismo de sobrevivencia como el mencionado por Palacios de la Rosa (Muñoz et al., 1983) llamado "latencia". Este mecanismo es de particular importancia, ya que se ha observado que bajo condiciones extremas de estrés hídrico, las plantas detienen completamente su crecimiento y aparentemente entran en un estado de latencia, y en el momento en que hay nuevamente humedad en el suelo, reinician su crecimiento hasta completarlo. Lo anterior está estrechamente relacionado con el fenómeno denominado "resurrección", ya que las plantas reviven completamente y reinician su metabolismo normal y crecimiento después de 24h del riego de recuperación (Graff, 1989; Neale et al., 2000). En este trabajo se pudo observar que en los tratamientos de sequía, las variedades detuvieron su crecimiento cuando el suelo alcanzó el PMP. La altura promedio en este punto fue de 17,3 ±0,56; 17,4 ±0,66; 18,2 ±0,71 y 15,5 ±0,94cm para Zo, Z19, Co y C16, respectivamente. Una vez que se aplicó el RR, las variedades se recuperaron y reiniciaron su crecimiento. La recuperación fue observada con mayor rapidez en las variedades mejoradas, alcanzando un crecimiento exponencial a los 14, 10 y 5 días para los tratamientos S1, S2 y S3 respectivamente, mientras que para S4 las plantas no alcanzaron la recuperación. La altura final para el tratamiento S3 fue de 74,1 ±5,3; 139 ±10,8; 108,5 ±6,9 y 137,7 ±7,3cm, para Zo, Z19, Co y C16, respectivamente. Las diferencias en altura entre las variedades no presentan diferencia estadística, debido a que cada una son de diferente porte.
Conclusiones
Las cuatro variedades mostraron diferente comportamiento a los tratamientos de sequía. Sin embargo, las variedades mejoradas registraron mayor biomasa y acumulación de prolina, lo cual sugiere mayor tolerancia a la sequía. El incremento del potencial osmótico en los tejidos como un reflejo de la falta de agua, fue detectado con mayor rapidez en las variedades mejoradas y puede ser considerado como parte de un mecanismo de respuesta rápida ante la falta de agua. En las cuatro variedades se observó una recuperación después del RR, semejante a lo registrado como latencia o resurrección, siendo más evidente en las mejoradas. Se considera que las variedades mejoradas Z19 y C16 pueden ser incluidas en programas de producción de maíz en áreas de valles altos de México con regímenes de baja precipitación.
REFERENCIAS
1.Attipalli RR, Kolluru VC, Munusamy V (2004) Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 161: 1189-1202. [ Links ]
2.Bajji M, Lutts S, Kinet JM (2000) Physiological changes after exposure to and recovery from polyethylene glycol-induced water deficit in roots and leaves of durum wheat (Triticum durum Desf.) cultivars differing in drought resistance. J. Plant Physiol. 157: 100-108. [ Links ]
3.Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39: 205-207. [ Links ]
4.Bogoslavsky L, Neumann PM (1998) Rapid regulation by acid pH of cell wall adjustment and leaf growth in maize plants responding to reversal of water stress. Plant Physiol. 118: 701-709. [ Links ]
5.Carceller M, Prystupa P, Lemcoff H (1999) Remobilization of proline and other nitrogen compounds from senescing leaves of maize under water stress. J. Agron. Crop Sci. 183: 61-66. [ Links ]
6.Desai SA, Singh RD (2001) Combining ability studies for some morphophysiological and biochemical traits related to drought tolerance in maize (Zea mays L.). Indian J. Gen. Plant Breed. 61: 34-36. [ Links ]
7.Dreesmann DC, Harn C, Daie J (1994) Expression of genes encoding Rubisco in sugarbeet (Beta vulgaris L.) plants subjected to gradual desiccation. Plant Cell Physiol. 35: 645-653. [ Links ]
8.García E (1988) Modificaciones al sistema de clasificación climatica de Koppen (Para adaptarlo a las condiciones de la República Mexicana). Editorial Inst. de Geografía. UNAM. México, D.F. 217 pp. [ Links ]
9.Girousse C, Bournoville R, Bonnemain JL (1996) Water deficit-induced changes in concentrations in proline and some other amino acids in the phloem sap of alfalfa. Plant Physiol. 111: 109-113. [ Links ]
10.Graff DF (1989) Responses of desiccation tolerant "resurrection" plants to water stress. En Krebb KH, Richter H, Hinkley TM (Eds.) Structural and Functional Responses to Environmental Stresses. SPB. The Hague, Holanda. pp. 255-268. [ Links ]
11.Harsh N (2003) Accumulation of osmolytes and osmotic adjustment in water-stressed wheat (Triticum aestivum) and maize (Zea mays) as affected by calcium and its antagonists. Envir. Exp. Bot. 50: 253-264. [ Links ]
12.Ibarra-Caballero J, Villanueva-Verduzco C, Molina-Galán JD, Sánchez-de-Jiménez E (1988) Proline accumulation as a symptom of drought stress in maize: A tissue differentiation requirement. J. Exp. Bot. 39: 889-897. [ Links ]
13.Ilahi I, Dorffling K (1982) Changes in abscisic acid and proline levels in maize varieties of different drought resistance. Physiol. Plant. 55: 129-135. [ Links ]
14.Molina GJD (1980) Selección masal para resistencia a sequía en maíz. Agrociencia 42: 69-77. [ Links ]
15.Molina GJD (1983) Selección masal visual estratificada. Centro de Genética. Colegio de Postgraduados. Chapingo, México. 35 pp. [ Links ]
16.Muñoz, OA (1980) Resistencia a la sequía y mejoramiento genético. Ciencia y Desarrollo 33: 26-35. [ Links ]
17.Muñoz O, Stevenson AKR, Ortiz CJ, Thurtell GW, Carballo CA (1983) Transpiración, fotosíntesis, eficiencia en uso de agua y potencial hídrico en maíces resistentes a sequía y a heladas. Agrociencia 51: 11-153. [ Links ]
18.Neale AD, Blomstedt CK, Bronson P, Le TN, Guthridge K, Evans J, Gaff DF, Hamill JD (2000) The isolation of genes from the resurrection grass Sporobolus stapfianus wich are induced during severe drought stress. Plant Cell Envir. 23: 265-277. [ Links ]
19.Ober ES, Sharp RE (1994) Proline accumulation in maize (Zea mays L.) primary roots at low water potentials. I. Requirement for increased levels of abscisic acid. Plant Physiol. 105: 981-987. [ Links ]
20.Petcu E, Terbea M (1996) Dynamics of the proline content in maize plants under drought conditions. Anal. Institut. Cercetari pentru Cereale Plante Tehnice, Fundulea. pp. 263-272. [ Links ]
21.Ramanjulu S, Sudhakar C (2000) Proline metabolism during dehydration in two mulberry genotYpes with contrasting drought tolerance. J. Plant Physiol. 157: 81-85. [ Links ]
22.Stewart RC, Hanson AD (1980) Proline accumulation as a metabolic response to water stress. En Turner NC, Kramer PJ (Eds.) Adaptation of plants to water and high temperature stress. Wiley. New York, EEUU. pp: 173-189. [ Links ]
23.Stone PJ, Wilson DR, Reid JB, Gillespie RN (2001) Water deficit effects on sweet corn. I. Water use, radiation use efficiency, growth, and yield. Aust. J. Agric. Res. 52: 103-113. [ Links ]
24.Van Heerden PDR, Kruger GHJ (2002) Separately and simultaneously induced dark chilling and drought stress effects on photosynthesis, proline accumulation and antioxidant metabolism in soybean. J. Plant Physiol. 159: 1077-1086. [ Links ]
25.Verslues PE, Sharp RE (1999) Proline accumulation in maize (Zea mays L.) primary roots at low water potentials. II. Metabolic source of increased proline deposition in the elongation zone. Plant Physiol. 119: 1349-1360. [ Links ]
26.Wassom JJ, Reese RN, Schumacher TE, Wicks ZW (2000) Proline accumulation in response to dehydration and diurnal hydration cycles varies among maize genotypes. Maydica 45: 335-343. [ Links ]
