











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.33 n.6 Caracas jun. 2008
Theoretical assessment of the impact of seeding on the exploitation of some economically viable marine populations
Ramón Lacayo 1 and Rodrigo Sfeir 2
1 M.Sc. in Physics and Mathematics, Peoples Friendship University, Moscow, USSR. M.Sc. in Statistics, Rutgers University, New Brunswick, NJ, USA. Doctor in Economics, Universidad de Barcelona, Barcelona, Spain. Associate Professor, Escuela de Ingeniería Comercial, Universidad Católica del Norte, Coquimbo, Chile. e-mail: rlacayo@ucn.cl
2 B.Sc. in Business Administration, Universidad Católica del Norte, Antofagasta, Chile. Doctor in Economic Development, Universidad Autónoma de Madrid, Madrid, Spain. Assistant Professor, Universidad Católica del Norte, Larrondo 1281, Coquimbo, Chile. e-mail: rsfeir@ucn.cl
SUMMARY
Many rural communities in the developing world practice small-scale harvesting of marine species of various kinds (fish, crustaceans, mollusks) as their main economic activity. Lack of regulation or enforcement or both has led in many cases to overexploitation of resources and consequent decline of the populations of organisms on which the communities depend. This situation, common along the north-central Chilean coast, served as the starting point of the present work. Recent advances in biotechnology and aquaculture, however, have made plausible halting or even reversing the trend towards resource depletion, favorably affecting the economic wellbeing of the communities. In this paper the theoretical implications of seeding the habitual harvesting grounds of a hypothetical community with individuals produced in the laboratory is explored. In particular, we examine the effect of periodic seeding of hatchlings or juveniles on the population of an economically important species harvested under either one of two modes of extraction, constant harvest or harvest proportional to the stock level. The population dynamics is modeled by a first order ordinary differential equation containing a series of delta functions, and which in words reads as follows: the population growth rate is equal to the natural growth rate minus the harvest rate plus the seeding rate. It was found that, at least from a theoretical standpoint, seeding could potentially lead to a notable increase in the productivity and well-being of these sea-based economies.
KEYWORDS / Aquaculture / Bioeconomics / Delta Function / Laplace Transform / Padé Approximant /
Evaluación teórica del impacto de siembra en la explotación de algunas poblaciones marinas económicamente importantes
RESUMEN
Muchas comunidades rurales en el mundo en desarrollo practican la recolección de especies marinas de varios tipos (peces, crustáceos, moluscos) como actividad económica principal. La falta de regulación o de aplicación de leyes ha conllevado en muchos casos a la sobreexplotación de los recursos y disminución de poblaciones de organismos de los que dependen estas comunidades. Esta situación, común en las zonas norte y centro del litoral chileno, sirvió como punto de partida para este trabajo. Avances recientes en biotecnología y acuicultura han hecho creíble la posibilidad de detener e incluso revertir la tendencia hacia el agotamiento del recurso y así afectar favorablemente el bienestar de la comunidad. En este trabajo se explora las implicaciones teóricas de la siembra con individuos producidos en el laboratorio en sectores usuales de captura de una comunidad hipotética; en particular, el efecto de la siembra periódica de crías o juveniles sobre la población de una especie económicamente importante cuya recolección se hace bajo uno de dos modos de extracción, constante o proporcional a la población. La dinámica poblacional se modela por una ecuación diferencial ordinaria de primer orden que contiene una serie de funciones delta y que, en palabras, dice que la tasa de crecimiento de la población es igual a la tasa natural de crecimiento menos la tasa de extracción más la tasa de siembra. Se encontró que, al menos desde el punto de vista teórico, la siembra podría potencialmente llevar a un incremento notable en la productividad y bienestar de estas comunidades marino-dependientes.
Avaliação teórica do impacto do plantio na exploração de algumas populações marinhas economicamente importantes
RESUMO
Muitas comunidades rurais no mundo em desenvolvimento praticam a colheita de espécies marinhas de vários tipos (peixes, crustáceos, moluscos) como atividade econômica principal. A falta de regulamentação ou de aplicação de leis tem conduzido em muitos casos à superexploração dos recursos e diminuição de populações de organismos dos quais dependem estas comunidades. Esta situação, comum nas zonas norte e centro do litoral chileno, serviu como ponto de partida para este trabalho. Avanços recentes em biotecnologia e aqüicultura têm tornado aceitável a possibilidade de deter e inclusive reverter a tendência para o esgotamento do recurso e assim afetar favoravelmente o bem-estar da comunidade. Neste trabalho se explora as implicações teóricas do plantio com indivíduos produzidos no laboratório em setores usuais de captura de uma comunidade hipotética; em particular, o efeito do plantio periódico de filhotes ou juvenis sobre a população de uma espécie economicamente importante cuja colheita se faz sob um de dois modos de extração, constante ou proporcional à população. A dinâmica populacional se modela por uma equação diferencial ordinária de primeira ordem que se representa por uma série de funções delta e que, em palavras, diz que a taxa de crescimento da população é igual à taxa natural de crescimento menos taxa de extração mais taxa de semeadura. Encontrou-se que, pelo menos desde o ponto de vista teórico, o plantio poderia potencialmente levar a um incremento notável na produtividade e bem-estar destas comunidades marinho-dependentes.
Received: 12/14/2006. Modified: 04/30/2008. Accepted: 05/02/2008.
Introduction
When analyzing traditional fishing practices of coastal, rural communities in the Coquimbo Region of Chile, it becomes apparent that these mostly economically disadvantaged people are highly dependent on the harvesting of benthic organisms such as the sea urchin and a variety of shellfish –Conchalepas conchalepas ("loco"), Fisurella latimarginata ("lapa") and Mesodesma donacium ("macha") among them– which are key to their survival. This overdependence on benthic resources can be explained as much by tradition as by the economic situation of these communities (lack of fishing equipment, vessels, storage facilities and transportation to either catch or commercialize other marine resources). The principal reason, however, is the depletion of commercially important species in the so-called historic fishing zones (SERNAPESCA, 2005).
In order to give some degree of rationality to the extraction of marine resources and ostensibly to make it a sustainable economic activity in the long run, in 1996 the Chilean government declared certain marine areas as protected (AMERB, Spanish acronym for Areas for the Management and Exploitation of Benthic Resources) and provided them with the needed regulation. Additionally, towards the end of 2004, the government approved legislation allowing aquacultural activities in these protected areas (Diario Oficial (2005).
Since everything points towards an ever increasing pressure on the benthic resources of these special zones (Palma and Chávez, 2006), the need to increase their productivity is evident. One solution to this conundrum may very well lie in their periodic repopulation (seeding) with individuals produced inland. There are some promising experiences with loco and macha in Chile (Pereira, 2004; Illanes, 2006) and this experience could be replicated anywhere. The situation is similar for many communities around the world.
Repopulation models are more or less often encountered in the field of botany, especially with regard to logging of forests and reforestation efforts. Marine stock enhancement projects of various sorts exist in many places throughout the world and have been evaluated mainly from an economic perspective (Hilborn, 1988; Moksness, 1999; Caddy and Defeo, 2003; Nakagawa et al., 2007). From a purely mathematical standpoint, however, we believe that our model represents a leap forward in the quest to understand the dynamics of repopulation of marine ecosystems in their natural setting. Aside from this Introduction, the paper contains four short sections. The first deals with a particular population growth model which we call "natural", that is, a population without human interference through either seeding or harvesting. In the next, periodic seeding is allowed and the (approximate) solution of the corresponding differential equation is found via a Laplace transform technique. The two complete and realistic models which are the main contribution of this paper are treated thereafter, after introduction of harvesting; in the first model harvesting occurs at a constant rate, while in the second it takes place at a rate that is proportional to the stock level. The final section is devoted to a mostly graphical and qualitative analysis of the results and contains a few conclusions. Finally, some brief conclusions are presented.
The Natural Population Growth Model
Let the size of the resource population at time t be p(t). We assume that the intrinsic growth rate of this population is r and that the saturation level of the environment (otherwise known as carrying capacity) is K. In our population model, the natural growth rate is based on the differential equation
p'(t) = r(1-p(t)/K) (1)
This choice of growth rate as a linear function of p is made in part to ensure a manageable level of mathematical tractability after seeding is introduced. It is assumed that the initial population of the ecosystem is p(0)=p0¹0. Note that the more popular logistic model has a growth rate that is quadratic in p and its use would have led to slightly more complicated calculations without adding much to the overall effect on the system of that which is of greatest interest to us, i.e., seeding.
Our goal is not to perform an economic analysis, and we do not deal with any specific species. We merely propose a new deterministic population model that takes into account seeding and study qualitatively its implications for the evolution of the population. Throughout the paper the notation and terminology of Clark (1990) is followed. For an example of a stochastic economic model that is species-specific, see Hanson and Ryan (1998).
In the model based on Eq. 1, p'(t)®r (p®0) and p'(t)®0 (p®K); that is, the population grows at a rate close to r when p itself is small, and it stops growing as it reaches the maximum size K permitted by the environment.
Eq. 1 is elementary. For an initial population p0, variable separation and integration yield
p(t)=K+(p0-K)e-rt/K (2)
It is obvious from this that the population approaches its maximum value K asymptotically, regardless of whether the initial value p0 is smaller or greater than K (except of course in the unlikely case of p0=K). Furthermore, by differentiating Eq. 2 we see that as a function of time the growth rate behaves exponentially.
Population Growth Model with Seeding
Formally, the model with seeding is just a simple modification of Eq. 1. In this case, though, the population growths at the natural rate determined by Eq. 1 plus the effect of a individuals being added to the system at times t= 0, 1, 2, In other words,
![]() (3)
(3)
where d(·)is Diracs delta function, introduced in the 1930s and widely used in physics and engineering for its unique ability to represent "point" sources of diverse nature (here the seeding at different points in time).
The (approximate) solution of Eq. 3 may be found using the Laplace transform technique (Abramowitz and Stegun, 1970). Let F(z) be the Laplace transform of p(t), and denote this relationship p(t)ÛF(z). Then, from standard tables and recalling that p(0)=p0, the differential Eq. 3 is transformed into
![]()
The expression above (an algebraic equation for F(z)) contains a geometric series of ratio r=e-z which may be summed trivially, provided ½e-z½<1. This occurs for Re z>0, that is, on the right-half of the complex z-plane. Assuming this is the case here, after collecting terms we obtain
![]()
or, equivalently,
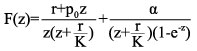 (4)
(4)
In order to determine p(t), the inverse Laplace transform of the right hand side of Eq. 4 must be found. The first term presents no difficulty since, in fact, after partial fraction decomposition (Sveshnikov and Tikhonov, 1978) it may be given a form suitable for the inversion by standard tables. After doing just that we obtain
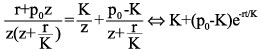 (5)
(5)
which is nothing but the solution given by Eq. 2 in the "natural" growth model.
We next find the (approximate) inverse of the second summand in Eq. 4 by using a Padé approximant of order (1,1) of the function 1/(1-e-z), namely, the function (1+z/2)/z (Bender and Orszag, 1987). After substitution and partial fraction decomposition we get
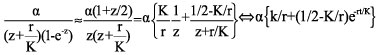 (6)
(6)
Note that one could as well have used a Padé approximant of higher order. For our purposes, however, the choice is sufficiently satisfactory.
Combining Eqs. 5 and 6 and collecting terms finally yields, from Eq. 4,
p(t)=K(1+a/r)+[p0+a/2-K(1+a/r)]e-rt/K (7)
This population growth model with seeding may be generalized to account for an extractive activity of the resource. This is accomplished by subtracting the harvesting rate h(t) from the right hand side of Eq. 3 and solving the corresponding differential equation. As was mentioned in the Introduction, we consider two cases: constant harvesting rate and harvesting rate that is proportional to the stock level. They are the subject of the following two subsections.
Constant harvesting rate
Let h(t)=hº constant. After subtraction of h from Eq. 3 the resulting equation may be written as
![]() (8)
(8)
where
![]() (9)
(9)
Since r* and K* are both constant, Eqs. 3 and 8 are equivalent. Consequently, the solution to Eq. 8 is Eq. 7 with parameters r and K replaced by r* and K* from Eq. 9, respectively; that is,
p(t)=K*(1+a/r*)+[p0+a/2-K*(1+a/r*)]e-r*t/K* (10)
Harvesting rate proportional to stock level
Let now h(t)=qEp(t), where the constants q and E are the catchability coefficient and the effort, respectively, in some appropriate units.
Proceeding as before, Eq. 3 may, after subtracting and factoring, be written in the form
![]() (11)
(11)
where
![]() (12)
(12)
Again, since Eqs. 11 and 3 are equivalent, the solution to Eq. 11 is Eq. 7 with the parameter K replaced by K** from Eq. 12, i.e.,
![]() (13)
(13)
Analysis
Two modes of harvesting (constant and proportional) and two modes of seeding (seeding and not seeding) have been considered. The combination of one type of harvesting and one of seeding, therefore, provide four situations that deserve some analysis. They are summarized in the four figures shown.
Figure 1 represents the behavior of the system in the absence of seeding and for various constant catch rates. This is Eq. 10 with the parameters given in (9) and a=0. The top curve depicts the natural growth of the population without human intervention (h=0) The population approaches K asymptotically. As the harvesting rate increases moderately, the population tends asymptotically to an ever decreasing long run limit below K. The curve next to the bottom shows what happens in the event h ® r from below: the population asymptotically vanishes. If h is large enough, there is extinction in a finite time (as shown by the lowermost curve, where the portion of the curve below the axis is left for illustration only).
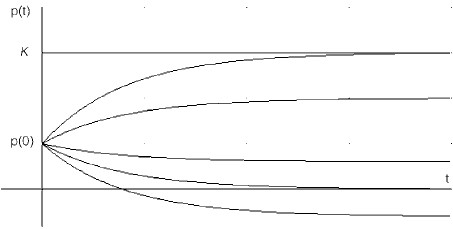
Figure 1. Constant harvesting rate h with no seeding (α=0). The curves shift downwards with increasing h.
In Figure 2 we again graph Eq. 10 with the same parameters shown in (9), except that we now have a fixed rate of harvesting h and seeding at various rates. Again, the curve which corresponds to no human influence, h=a=0, is shown (second from the top). For the given harvesting rate h, this curve coincides with the curve obtained for h=a; that is, it shows that, as could be expected, there is a non-zero seeding rate which completely cancels the effect of harvest at the same rate. With increasing a the population reaches a higher steady state limit. The lowermost curve shows that for a fixed h large enough and a sufficiently small there is the possibility of extinction in a finite time and, on the other extreme, the top curve shows that with a seeding rate sufficiently large the population may reach K in a finite time (the portions of the curves below the t-axis and above K are left for illustration only).
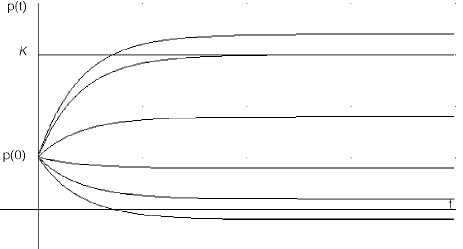
Figure 2. Constant harvesting rate (the same for all) with seeding. Increasing α displaces the curves upwards.
Figures 3 and 4 illustrate what happens to the population in the case of proportional harvesting as obtained from the solution (13) using parameter (12).
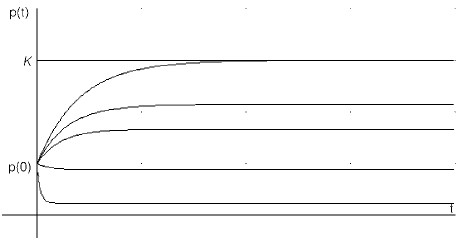
Figure 3. Proportional harvesting without seeding (α=0). With increasing qE (the proportionality coefficient) the curves are displaced in the downward direction.
In Figure 3 we combine no seeding (a=0) with a few values of qE=constant. Increasing qE leads to an expected downward shift of the population curves, showing that a larger qE implies a smaller population at equilibrium. A crucial difference with the previous two cases is that extinction of the resource is no longer a possibility, at least not in a finite time. This is so because a reduction in population size automatically triggers a reduction in harvesting.
Figure 4 shows what happens to the population in the presence of an increasing seeding rate a, beginning with a=0 (the curve at the bottom) for a fixed constant qE. After a certain critical a the population grows even in the presence of an increasing catch. This is a significant result. It means that at least during the initial interval when this is true, the captures may be increased and the population still expected to grow towards its maximum, which for certain values of the parameters may very well be K.
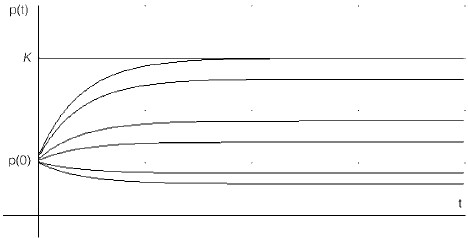
Figure 4. Proportional harvesting (qE the same for all) with seeding. As α increases the curves are displaced in the upward direction.
Conclusions
This study was born out of the need to assess qualitatively the effect of seeding in the exploitation of benthic resources in special fishing zones in Chile. Many coastal communities throughout the world, especially in developing countries, find themselves in similar circumstances of resource depletion. Although the population model employed is simple, it seems to capture the relevant aspects of the problem. And from the analysis of the two competing modes of harvesting it seems that, in the presence of seeding, proportional harvesting should be implemented in order to ensure an increased productivity and the sustainability of the economic activity. This mode has the dual benefit of replenishing the stock while at the same time, in contrast to the usual quota system prevalent in the sector, avoids the possibility of complete depletion of the resource. It is our strong belief that this model, with pertinent modifications for location and resource, may be implemented anywhere and bring important economic benefits to those involved.
References
1. Abramowitz M, Stegun IA (1970) Handbook of Mathematical Functions. Dover. New York, USA. 1059 pp. [ Links ]
2. Bender CM, Orszag SA (1987) Advanced Mathemat ical Methods for Scientists and Engineers. McGraw-Hi l l . Singapore. 593 pp. [ Links ]
3. Caddy JF, Defeo O (2003) Enhancing or restoring the productivity of natural populations of shellfish and other marine invertebrate resources. FAO Fisheries Technical Paper N° 448. 159 pp. [ Links ]
4. Clark CW (1990) Mathematical Bioeconomics. Wiley. New York, USA. 386 pp. [ Links ]
5. Diario Oficial (2005) Num. 314. Reglamento de actividades de acuicultura en áreas de manejo y explotación de recursos bentónicos. Subsecretaría de Pesca. Ministerio de Economía, Fomento y Reconstrucción. Diario Oficial de la República de Chile Nº 38.203 (06/07/2005). [ Links ]
6. Hanson FB, Ryan D (1998) Optimal Harvesting with Both Population and Price Dynamics. Math. Biosci. 148: 129-146. [ Links ]
7. Hilborn R (1988) The economic performance of marine stock enhancement projects. Bull. Mar. Sci. 62: 661-674. [ Links ]
8. Illanes J (2006) Acuicultura para la recuperación del recurso macha Mesodesma donacium (Lamarck, 1818) en áreas de manejo de comunidades artesanales. Informe Final Proyecto FONDEF D02I1131. Universidad Católica del Norte. Coquimbo, Chile. 225 pp. [ Links ]
9. Moksness E (1999) Economic evaluation of marine ranching. FAO Fish. Circ. 943: 44-52. [ Links ]
10. Nakagawa M, Okouchi H, Adachi J, Attori K, Yamashita Y (2007) Effectiveness of stock enhancement of hatchery-released black rockfish Sebastes schlegeli in Yamada Bay – Evaluation by a Fish Market survey. Aquaculture 263: 265-302. [ Links ]
11. Palma M, Chávez C (2006) Normas y cumplimiento en áreas de manejo de recursos bentónicos, Estudios Públicos 103: 237-276. [ Links ]
12. Pereira L (2004) Innovaciones tecnológicas para repoblamiento y producción de lapas chilenas de exportación (Fisurella latimarginata y F. cumingi) en áreas de manejo y centros de cultivo. Informe Final Proyecto FONDEF D00I1141. Universidad Católica del Norte. Coquimbo, Chile. 180 pp. [ Links ]
13. SERNAPESCA (2005) Evaluación técnica y económica del impacto de las Áreas de Manejo y Explotación de Recursos Bentónicos. Servicio Nacional de Pesca. Valparaiso, Chile. 24 pp. [ Links ]
14. Sveshnikov A, Tikhonov A (1978) The theory of functions of a complex variable. Mir. Moscow, USSR. 333 pp. [ Links ]
