











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.24 n.4 Caracas abr. 2009
Structure of the shrub-arboreal component of an atlantic forest fragment on a hillock in the central lowland of Rio de Janeiro Brazil.
Alexandre Gabriel Christo, Rejan R. Guedes-Bruni, Felipe De Araújo P. Sobrinho, Ary Gomes Da Silva and Ariane Luna Peixoto
Alexandre Gabriel Christo. Agronomic Engineer. M.Sc. in Botany, Escola Nacional de Botânica Tropical (ENBT), Brazil. Instituto de Pesquisas Jardim Botânico do Rio de Janeiro (JBRJ), Brazil. Address: Jardim Botânico do Rio de Janeiro, Rua Pacheco Leão, 915, 22.460-030 - Rio de Janeiro, RJ, Brazil. e-mail: achristo@jbrj.gov.br.
Rejan R. Guedes-Bruni. Biologist. D.Sc. in Ecology, Universidade de São Paulo (USP), Brazil. Senior Researcher, JBRJ, Brazil. e-mail: rbruni@jbrj.gov.br.
Felipe de Araújo P. Sobrinho. Forest Engineer. M.Sc. in Botany, ENBT, Brazil. Doctoral student, USP, Brazil. e-mail: felipesobrin@yahoo.com.br.
Ary Gomes da Silva. B.Sc. in Pharmacy. D.Sc. in Plant Biology, Universidade Estadual de Campinas (UNICAMP), Brazil. Professor, Centro Universitário Vila Velha, Brazil. e-mail: arygomes@uvv.br.
Ariane Luna Peixoto. Naturalist. D.Sc. in Plant Biology, UNICAMP, Brazil. CNPq Fellow, Associate Researcher, JBRJ, Brazil. e-mail: ariane@jbrj.gov.br.
SUMMARY
The present study describes and evaluates the horizontal and vertical structures of a lowland forest fragment on a hillock in the municipality of Silva Jardim, Rio de Janeiro State, Brazil (22o3156S and 42o2046W). Twenty plots (10´2m) totaling 0.5ha were laid out following the slope grade using DBH³5cm as the inclusion criterion. A total of 734 individuals were encountered, yielding a total density of 1468 ind./ha and a total basal area of 10783m2. The richness values (129 species/41 families), Shannon-Wiener diversity (4.22) and equitability (0.87) indices indicated an accentuated floristic heterogeneity and low ecological dominance. Lauraceae, Myrtaceae, Fabaceae and Euphorbiaceae showed the greatest species richness, corroborating other studies that indicated these species as the most representative of Atlantic Forest areas in southeastern Brazil. The species with the greatest importance values (VI) were Aparisthmium cordatum, Guapira opposita, Lacistema pubescens, Xylopia sericea, Tapirira guianensis and Piptocarpha macropoda. The high diversity observed was influenced by earlier anthropogenic actions and by the current successional stage. The forest fragment studied demonstrated closer floristic similarity to areas inventoried in a close-by biological reserve than to fragments dispersed throughout the coastal plain. Similarities in soil type, degree of soil saturation and use-history of forest resources all support these relationships. The fragmented physiognomy of the central lowland in this region and the use-history of the landscape make these small remnant forest areas important in terms of establishing strategies for landscape restoration and species conservation.
Estructura del componente arbustivo-arbóreo de un fragmento de selva atlántica en una colina de las tierras bajas del centro de Río de Janeiro, Brasil.
RESUMEN
Este estudio tuvo como objetivo describir y evaluar las estructuras horizontal y vertical de bosques de tierras bajas en pequeñas colinas (22º31'56''S y 42º20'46''O) en el municipio de Silva Jardim, Río de Janeiro, Brasil. Fueron asignadas, a lo largo de la pendiente, 20 parcelas (10´25m) totalizando 0,5ha, con criterio de inclusión de DAP³5cm. Se registraron 734 individuos, con una densidad total de 1468 ind/ha y 10783m2 de área basal total. Los valores de riqueza (129 especies/41 familias), diversidad de Shannon-Wiener (4,22) y homogeneidad (0,87) indican una acentuada heterogeneidad florística y baja dominancia ecológica. Lauraceae, Myrtaceae, Fabaceae y Euphorbiaceae presentan la mayor riqueza, corroborando estudios que las muestran como las más representativas en la composición florística de la Selva Atlántica del sudeste de Brasil. Las especies de mayor valor de importancia (VI) fueron Aparisthmium cordatum, Guapira opposita, Lacistema pubescens, Xylopia sericea, Tapirira guianensis y Piptocarpha macropoda. La elevada diversidad encontrada es justificada por la acción antrópica anterior y el actual estado sucesional. El fragmento estudiado presenta mayor similitud florística con áreas inventariadas en una reserva biológica cercana que con los fragmentos dispersos por la región llana costera. Semejanzas de tipo de suelo, saturación hídrica e histórico de uso de los recursos forestales soportan esta similitud. La condición de fragmentación que caracteriza la región de planicie costera central del estado de Río de Janeiro y el aumento de datos disponibles hacen de estos pequeños remanentes forestales áreas de marcada relevancia para el establecimiento de estrategias de restauración del paisaje y conservación de especies.
Estrutura do componente arbustivo-arbóreo de um fragmento de floresta atlântica em terras baixas sobre morrote mamelonar na parte central costeira do Rio de Janeiro, Brasil.
RESUMO
Neste estudo objetivou-se descrever e avaliar as estruturas horizontal e vertical de floresta de baixada sobre morrote mamelonar (22º31'56''S e 42º20'46''O) no município de Silva Jardim, RJ. Para tanto, alocou-se ao longo do eixo de aclive, 20 parcelas (10´25m), totalizando 0,5ha, com critério de inclusão DAP³5cm. Registrou-se 734 indivíduos, perfazendo densidade total por área de 1468 ind/ha e 10783m2 de área basal total. Os valores de riqueza (129 espécies/41 famílias), diversidade de Shannon (4,22) e de eqüitabilidade (0,87) indicam acentuada heterogeneidade florística e baixa dominância ecológica. Lauraceae, Myrtaceae, Fabaceae e Euphorbiaceae apresentaram as maiores riquezas, corroborando outros estudos que as indicam como as mais representativas na composição florística da Floresta Atlântica do Sudeste do Brasil. As espécies de maior importância (VI) foram Aparisthmium cordatum, Guapira opposita, Lacistema pubescens, Xylopia sericea, Tapirira guianensis e Piptocarpha macropoda. A elevada diversidade encontrada se justifica pelas ações antrópicas pretéritas e pelo atual estágio sucessional. O fragmento estudado apresenta maior similaridade florística às áreas inventariadas no Rebio do que aos fragmentos dispersos pela região da planície. As semelhanças do tipo de solo, grau de saturação hídrica e histórico de uso dos recursos florestais suportam esta similaridade. A condição de fragmentação que caracteriza a região da planície central costeira do Rio de Janeiro, acrescida dos dados disponíveis desta paisagem, tornam estes pequenos remanescentes florestais como áreas de relevância para o estabelecimento de estratégias de restauração da paisagem e conservação de espécies.
KEYWORDS / Atlantic Forest / Floristic Similarity / Lowland Forest / Structure /
Received: 09/29/2008. Modified: 03/16/2009. Accepted: 03/18/2009.
After five centuries of occupation and the effects of various economic cycles, the Brazilian Atlantic Forest has been reduced to about only 7.6% of its original extension (MMA, 1998). This drastic deforestation was driven by anthropogenic pressure on the forest, principally due to the selective extraction of valued arboreal species, agricultural expansion, lumbering and the use of wood as firewood by many industrial sectors (brick and tile manufacturing, residential expansion, etc.).
Within this historical context, remnant tracts of lowland forests in Rio de Janeiro State are now mostly limited to granite hillocks <300m in altitude and to fluvial plains along the coast. These low hills are surrounded by alluvial forest areas that have been occupied by homes and agricultural plots due to their leveled topography, even though they are often swampy (Guedes-Bruni et al., 2006a).
The forests that cover the low hills are generally altered remnants of the native vegetation that existed before anthropogenic processes dominated the lowland regions around Rio de Janeiro. These low hills are often totally deforested, but they are sometimes covered by remnant forests of different dimensions and in diverse successional states. Cattle-raising is the most evident sign of human intervention, and hand in hand with fire has determined the alteration of these landscapes and the species growing there (Guedes-Bruni et al., 2006a).
Although greatly fragmented, the lowland Atlantic Forest in the central region of Rio de Janeiro State is of great ecological importance, not only because of the species typical of this vegetation formation but also because it is the only habitat of the Golden-Lion-Tamarin (Leonthopitecus rosalia Lesson, 1940), a primate species endemic to this region and threatened with extinction.
Numerous authors have contributed to the knowledge of the composition and floristic structure of this environment, which is characterized by extensive fragmentation and a diversity of successional stages in the remnant areas (Guedes, 1988; Borém and Ramos, 2001; Borém and Oliveira-Filho, 2002; Carvalho et al., 2006a, Guedes-Bruni et al., 2006a; Pessõa and Oliveira, 2006: Carvalho et al., 2007; and others).
The present study describes and evaluates the horizontal and vertical structure of a section of lowland forest growing on hillocks and provides information that can contribute to conservation programs in the region.
Study area
The study area is part of the Santa Helena farm (22o3156S and 42o2046W; Figure 1), located along the BR 101 federal highway in the municipality of Silva Jardim (RJ), ~5km from the Poço das Antas Biological Reserve (Rebio) in an area of Dense Ombrophilous Lowland Forest (IBGE, 1992). The forest fragment examined covers 4.25ha and is surrounded by agricultural and grazing areas. The regional climate is classified as As by the Köeppen system: rainy-tropical, with a winter dry season (Bernardes, 1952). Average annual rainfall and temperature are 1995mm and 25.5oC, respectively (Lima et al., 2006).
The topography of the region is predominantly plain, with hillocks in the coastal lowlands (granite outcrops), whose height ranges from 19-200m, separated by flood plains and swamps (IBDF/FBCN, 1981). The surroundings of Rebio are in full field of Tertiary and Quaternary plains, with dated rocks of Pre-Cambrian and typical of the Coastal Complex Fluminense (IBDF/FBCN, 1981). The classes of soil (EMBRAPA, 1999) are divided, basically, between Oxisols, Ultisols, Incepitsols (Aquepts).
Methods
Twenty plots (10´25m) were laid out along the line of the hill slope, covering a total sampling area of 0.5ha. In laying out the plots a minimum distance of 20m was maintained from any trail so that, essentially, only the central area of the fragment was sampled. All plants with trunk diameters at 1.3m above soil level (DBH) ³5cm were measured and identified. All testimonial material was deposited in the herbarium of the Rio de Janeiro Botanical Garden (RB). The taxonomic system adopted followed APG (2003).
To elaborate a vegetation profile, a 50´5m plot was established following the slope of the hillock according to the methodology suggested by Melo (2002). The profile diagram was elaborated in true height and diameter scales, using the AutoCAD 2000 software program.
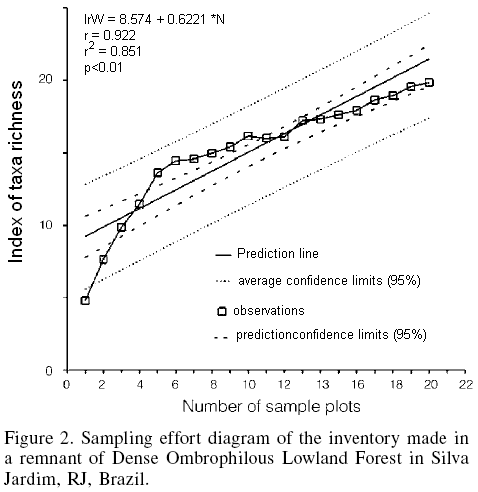
Sampling effort was measured by determining the simple linear regression line (Zar, 1999) of the accumulation of taxon richness (Whittaker, 1975) in regards to the accumulated number of observations (Loss and Silva, 2005), corresponding to the sequential implantation of the plots.
Structural analyses were made by calculating the absolute and relative values of density (DR), frequency (FR) and dominance (DoR), whose sum, in relative terms, defines the Importance Value (VI) of the species. Estimations of vegetation structure were obtained according to Müeller-Dombois and Ellenberg (1974). The Shannon-Wiener diversity index (H) and equitability (J) were calculated according to Magurrán (1988) and Pielou (1975), respectively. The Fitopac 1 software package was used to calculate all phytosociological parameters (Shepherd, 1995).
The 10 species with highest VI were grouped into three categories, pioneers, early secondary and late secondary, according Gandolfi et al. (1995), and complemented on the basis of Paula et al. (2004) and Carvalho et al., (2006a).
To evaluate the horizontal structure of the vegetation, frequency histograms of diameter classes at 5cm intervals were elaborated for all individuals encountered. The vertical structure, as a characterization of the forest layers studied, was evaluated based on the allometric relationships between diameter and height (log transformed; Sneath and Sokal, 1973), identifying the canopy with a dotted line as seen on the correlation diagram, and characterizing the canopy individuals as having a specified diameter and height, confirmed by verifying the first discontinuity among the points in the diagram (Guedes-Bruni, 1998).
A graph was elaborated of the vertical space occupied by the sampled species that attained 75% of the VI. To determine the minimum number of classes, as well as their amplitude, the formula proposed by Spiegel (1977) apud Lopes et al. (2002) was used.
Relationships of floristic similarity were sought between the study area and other forest fragments inventoried in this coastal plain region. To that end, a presence/absence matrix was elaborated of the species encountered in this study and combined with 17 other areas (totaling 279 species), eight of which were on private lands and nine of which were within the Rebio Poço das Antas. The private lands are represented by Fazendas Imbaú (Faz. Imb.), Andorinhas (Faz. And.), Estreito (Faz. Est.), Afetiva-Jorge (Faz. Aft.) and Sítio do Professor (Faz. Stp.) reported by Carvalho et al., 2006a; Santa Fé (Faz. Sfe) from unpublished data; and two topographic sequences at the Biovert Agroflorestal farm: F. Bio TPA (Borém and Ramos, 2001) and F. Bio TMA (Borém and Oliveira-Filho, 2002). The studies at the Rebio Poço das Antas covered Rebio 20a and 40a (Neves, 1999), Rebio Mor (Guedes-Bruni et al., 2006a), Rebio Aluv (Guedes-Bruni et al., 2006b), Rebio Fr1, Fr2 and Fr3 (Pessõa and Oliveira, 2006) and Rebio Ari and Cam (Carvalho et al., 2006b).
Any species only recorded at a single area was excluded from the list, as they do not cause any variation in the analysis. Based on the obtained matrix, Sorensens similarity coefficient was calculated for all the areas, forming a dendrogram by the UPGMA method. Calculations were performed utilizing the Fitopac 1 software package (Shepherd, 1995).
Results and Discussion
The results indicated a high species richness in the area, which is probably far from the real richness, as the sampling effort diagram exhibits a still ascending curve not highly significant different from a straight line in terms of the richness of the sampling points (Figure 2), indicating that there were still other taxa to be collected in the area. The vegetation sampling encountered 734 live individuals, yielding a total density of 1.468 ind/ha, a total basal area of 10783m2, and generating a taxon richness index of 19.85.
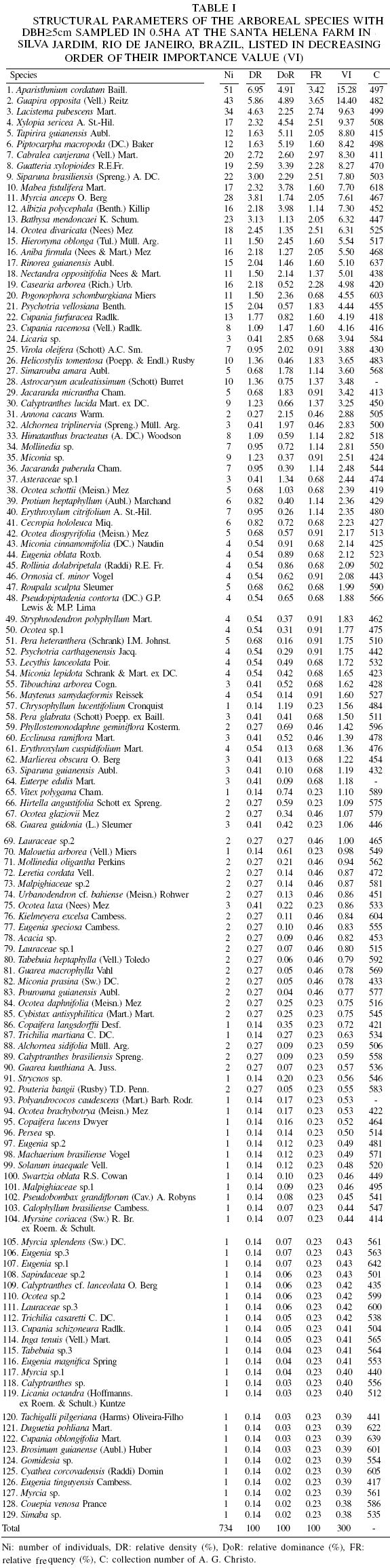
Among the sampled individuals were 129 species, subordinated to 41 families. The species, with their respective parameters, are listed in Table I in decreasing order of importance value (VI). Shannon-Wiener diversity index (H) was 4.22 and equitability (J) was 0.87.
Earlier studies in central lowland forests (Neves, 1999; Borém and Ramos, 2001; Borém and Oliveira-Filho, 2002; Guedes-Bruni et al., 2006a, b; Pessõa and Oliveira, 2006; Carvalho et al., 2007) yielded H values from 1.75 to 4.57, corroborating the general pattern of the local landscape that has suffered from continuous anthropogenic interferences, whether as a result of fire or the establishment of small farm plots or grazing areas. Added to these alterations was the construction of the Juturnaíba reservoir in the 1970s, which had a strong impact on vegetation cover along the valley of the São João River (Cunha, 1995).
The families with the greatest abundance values were Euphorbiaceae, Lauraceae, Myrtaceae, Annonaceae, Nyctaginaceae, Rubiaceae and Fabaceae. When their importance values were considered together, these families were found to be responsible for ~50% of the total VI. Previous studies in lowland and montane forests of Rio de Janeiro (Guedes-Bruni, 1998) have reported similar results, which reflect the wide geographic distribution of the species that integrate this vegetation type.
The families with the greatest species richness in the forest fragment studied were: Lauraceae (18 spp.), Myrtaceae (17), Fabaceae (11), Euphorbiaceae (8) and Meliaceae (6). These families together were responsible for 46.51% of all the species collected, with Myrtaceae alone yielding 13.95%.
Oliveira-Filho and Fontes (2000) analyzed 125 areas of Atlantic Forest (sensu lato) and reported that the families Myrtaceae, Rubiaceae, Euphorbiaceae and Melastomataceae had the greatest species richness in ombrophilous forests in southeastern Brazil. This data corroborates with that of other authors (Borém and Ramos, 2001; Borém and Oliveira-Filho, 2002; Carvalho et al., 2006a; Guedes-Bruni et al., 2006a, b; Pessõa and Oliveira, 2006; among others) in floristic analyses of the central coastal region of the state of Rio de Janeiro.
Myrtaceae and Lauraceae are characteristic in more mature forests (Tabarelli, 1997). Therefore, despite their representation in species richness, both families with many late successional species, contributed little in the composition of the VI.
The distribution of 10 species with higher VI (33.65% of sampled individuals) in successional groups showed a predominance (90%) of early secondary species, indicating that the forest is in a development process, or that there is some factor(s) preventing the full development of this fragment. Among these factors is the selective felling of some species (Euterpe edulis and Xylopia sericea), which was observed during data collection.
The families Euphorbiaceae, Lauraceae, and Annonaceae all contributed with 39.0% to the total basal area. The elevated number of individuals of the first two stood out, while Annonaceae showed the highest average basal area of individuals.
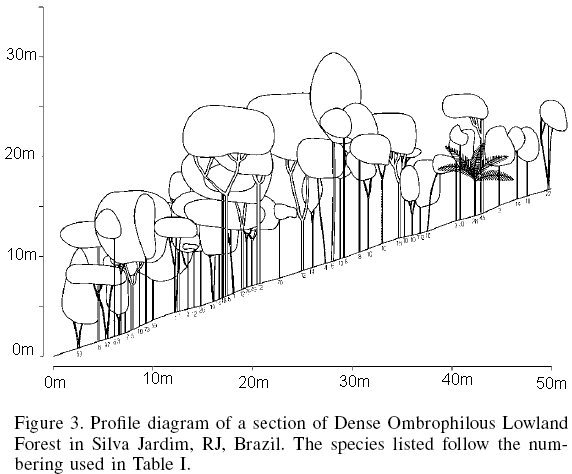
In terms of the sizes of the various populations occurring in the fragment, the species Aparisthmium cordatum, Guapira opposita, Lacistema pubescens, Myrcia anceps and Bathysa mendoncaei stood out as having the largest numbers of individuals. As this forest was in an intermediate stage of succession, large numbers of individuals of these species were to be expected. Xylopia sericea, Jacaranda micrantha and Piptocarpha macropoda had the largest individuals (26, 23, and 21m, respectively) encountered in the survey, while an individual of Licaria sp had the largest recorded diameter (51.3cm). A schematic physiognomic structural profile of the vegetation is shown in Figure 3.
The most frequent (FR) species in the sampling area was G. opposita, which was present in 80% of the surveyed plots. This species was followed by A. cordatum (75%), Cabralea canjerana (70%) and L. pubescens (65%). This was expected considering the altered state of the vegetation, not only because they are characteristically initial secondary species (Gandolfi et al., 1995) but as a result of this status they are among the most widely geographically distributed species, showing the highest frequencies in most inventories performed in Atlantic forests (Siqueira, 1994; Oliveira-Filho and Fontes, 2000).
P. macropoda, Tapirira guianensis, A. cordatum, G. opposita, and X. sericea had the highest dominance (DoR) values. Together, these five species retained 24.64% of the total relative dominance of the fragment studied. Of a total of 129 species, 43 were responsible for 75.54% of the importance value, with A. cordatum showing the highest VI of all the species present.
The large VIs of these species were due either to their large relative density, large relative dominance or large relative frequency. The relative density of A. cordatum (6.95%) was larger, for example, than that of G. opposita (5.86%); the relative dominance of P. macropoda (5.19%) was greater than that of T. guianensis (5.11%); and the relative frequency of G. opposita (3.65%) was larger than that of A. cordatum (3.42%). Although L. pubescens demonstrated only intermediate levels of density, dominance and frequency as compared to the other species, these consistent values were sufficient to place it among the species with the highest overall VI values.
Borém and Oliveira-Filho (2002) previously reported E. edulis, Astrocaryum aculeatissimum, Pseudopiptadenia contorta, Casearia sylvestris Sw., and Vochysia laurifolia Warm. as having large VI values; while Neves (1999) reported Miconia cinnamomifolia, Gochnatia polymorpha (Less.) Cabrera, Pithecellobium pedicellare (DC.) Benth., A. aculeatissimum and Sparattosperma leucanthum (Vell.) K. Schum. in an area under recovery for 20 years, and L. pubescens, Myrcia fallax (Rich.) DC., Cupania racemosa, Cupania schizoneura and Guatteria sp. in an area left undisturbed for 40 years. Pessõa and Oliveira (2006) evaluated three forest fragments inside the Poço das Antas Biological Reserve and recorded Senefeldera verticillata (Vell.) Croizat, Trema micrantha (L.) Blume, Andradea floribunda Allemão, Annona cacans and A. aculeatissimum in fragment II, which was approximately the same size as the fragment examined in the present study.
The histogram presented in Figure 4 demonstrates that most of the 734 individuals encountered (88.01%) were concentrated in the 5-20cm diameter classes, indicating the presence of very few large individuals. The average diameter observed during the sampling was only 11.77 ±7.06cm (average ±SD), and only four species (Licaria sp, T. guianensis, Chrysophyllum lucentifolium and A. cacans) had individuals with diameters >40cm.
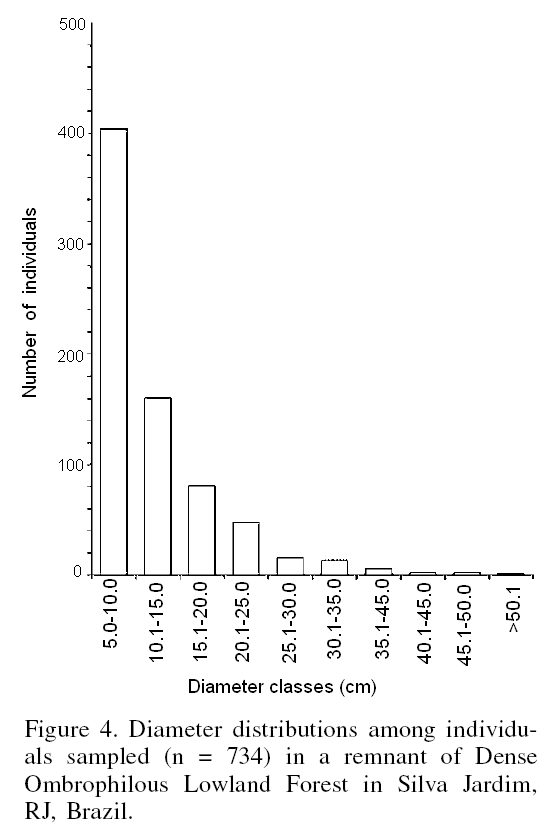
Analyses of the fragment showed a concentration of individuals in the smallest diameter classes (and inverted J pattern), indicating natural regeneration of the site.
The three species with the greatest VI (A. cordatum, G. opposita and L. pubescens) also demonstrated an inverted J pattern (Figure 5) indicating, again, the process of natural regeneration in the fragment.
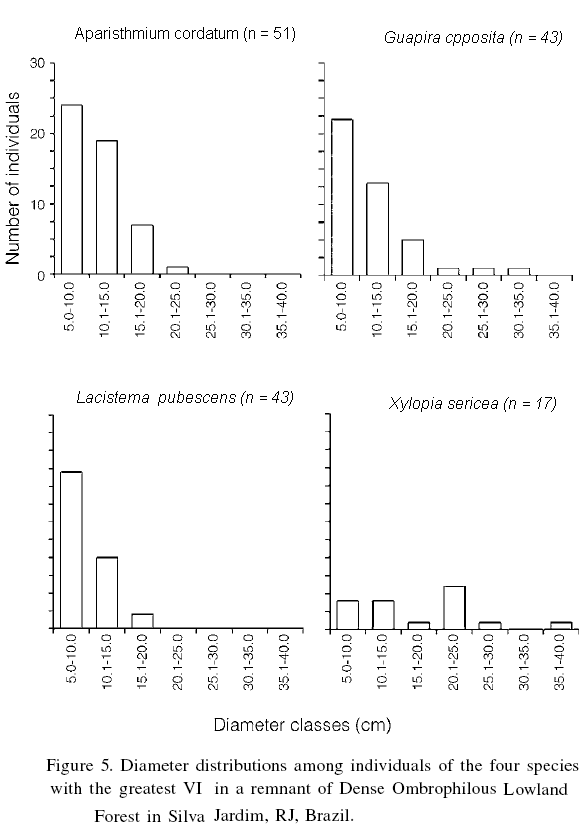
The individuals of X. sericea demonstrated a discontinuous aging structure pattern, which may be explained by the selective felling of these trees by the local population. This species is preferentially used for building roofs for houses and barns, and individuals with diameters 5-15cm are harvested for this purpose (Christo et al., 2006).
A selective extraction of species from the fragment was also indicated by the population size of the palm species E. edulis (0.41% of the relative frequency), as well as by physical evidence of harvesting in the area. The whole individual must be cut in order to obtain the palm-heart, as this species does not ramify. This species is considered one of the most important plants in the Atlantic Forest and is one of the principal non-wood forest resources (Fantini and Guries, 2007) available in both well-conserved and disturbed areas (Guedes, 1988; Borém and Oliveira-Filho, 2002).
Some authors consider the absence of large trees to be related to selective felling and to natural mortality as a result of edge-effects, as has been observed in tropical forest fragments (Laurance et al., 2000; Scariot et al., 2003). Scariot et al. (2003) recognized that the removal of canopy individuals facilitated the growth of juvenile plants, provoking alterations in successional processes and in the internal organization of the forest.
The evaluation of the allometric relationships between diameter and height of the 734 individuals sampled did not demonstrate any clear stratification in the fragment, as the dispersal diagram did not show any height discontinuities in relation to arboreal/shrub diameters (Figure 6).
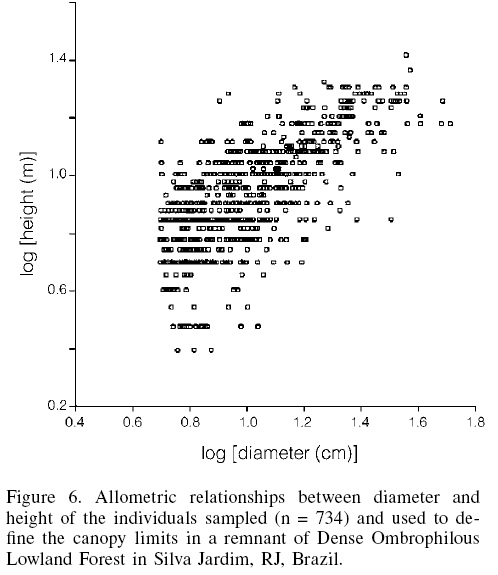
The individuals within the fragment had an average height of 9.44 ±3.96m (average ±SD) and were distributed among ten height classes at 2.25m intervals, which was rounded off here to 2.5m due to the calibration of the tree height measure in 0.5m units. Most individuals were 5-10m tall, with only one individual of each of the species X. sericea, P. macropoda, and J. micrantha reaching heights greater than 20m.
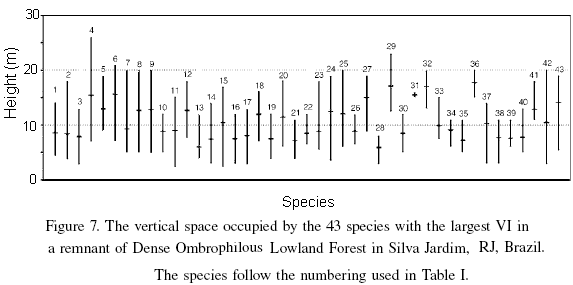
Figure 7 shows the vertical space occupied by 43 species, representing ~75% of the total VI, classified in increasing order of their Importance Value index. X. sericea, Ocotea diospyrifolia, Licaria sp, C. canjerana, Guatteria xylopioides, and Mabea fistulifena stood out as demonstrating the greatest amplitude within the vertical space they occupied. These species are not only typical of lowland forests in Rio de Janeiro (Guedes-Bruni et al., 2006a, b) but are also characteristic of the canopy layer, where they are encountered as large adult individuals, sometimes emergents, especially O. diospyrifolia and C. canjerana in well-conserved forests.
The dendrogram of floristic similarity produced by grouping analysis based on the presence/absence of species in the 18 areas evaluated can be seen in Figure 8. The results demonstrate a considerable fit of the similarity matrix to the estimated clustering that produce the dendrogram (rcs= 0.8806) and reveals the formation of five groups with a fusion level of 30%. Group 1 is formed by six areas: Faz. Imb., Faz. Est., Faz. Stp., Faz. And., Faz. Aft. and Faz. Sfe, all of which are located in the District of Imbaú. This assemblage is justified by having most of its species in the initial stages of ecological succession, and because these areas are close by one another and are all on private lands and, thus, more vulnerable to disturbances. Group 2 is formed by two areas that have gone undisturbed for 20 and 40 years (Rebio 20a and Rebio 40a, respectively), the present study area Faz. She, as well as two areas with inclusion criteria of DBH³10cm (Rebio Aluv and Rebio Mor) which are located in the interior of the Rebio; these areas are characterized by being better conserved, having been protected from felling, and having a wide range of canopy species and species that belong to later stages of ecological succession. Group 3 is formed by two privately held areas belonging to the same topographic sequence (F. Bio TPA and F. Bio TMA) but having distinct use-histories. Group 4 is formed by three different sized fragments that are isolated from each other but located within the Rebio (Fr1, Fr2 and Fr3). Group five is formed by the remaining two areas, which are also located in the interior of the Rebio (Ari and Cam) but are located in alluvial areas and contain species typical of this physiognomy, and are distinguishable from the other areas in terms of their floristic composition.
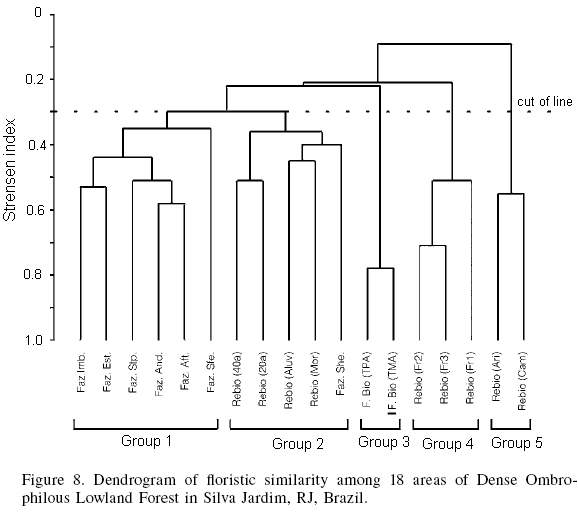
The species of the 18 areas analyzed are distributed among 52 botanical families, with 10 families (Fabaceae (sensu lato), Lauraceae, Myrtaceae, Euphorbiaceae, Annonaceae, Sapotaceae, Rubiaceae, Bignoniaceae, Meliaceae and Melastomataceae) concentrating 59.5% of the species diversity.
The species with the widest distribution (occurring in at least 10 areas) are C. sylvestris, G. opposita, X. sericea, Alchornea triplinervia, Brosimum guianense, T. guianensis, A. aculeatissimum, L. pubescens, Apuleia leiocarpa, Pseudopiptadenia contorta, Miconia cinnamomifolia, C. canjerana, and Guarea guidonia.
Experimental plantations in the Rebio, based on floristic-structural studies (Morães et al., 2006) have shown the importance of a number of species recorded in the present inventory, including G. guidonia, Calophyllum brasiliense, Copaifera langsdorffii, Alchornea triplinervia, Pseudobombax grandiflorum (Cav.) A. Robyns, T. guianensis, Nectandra oppositifolia, E. edulis and Guapira opposita; and these taxa have significant potential for use in the recuperation of altered areas in the region.
Conclusions
The presence of known primary successional species among the plants with the greatest VI (Aparisthmium cordatum, Guapira opposita, Lacistema pubescens, and Xylopia sericea), associated with an accentuated concentration of individuals in the 5-20cm diameter classes, and with the diminishing use of the forest resources by humans, are all signs that the study fragment is in a secondary phase of succession and natural regeneration.
The floristic similarity observed between this fragment and other areas inventoried in the interior of the Rebio is supported by their geographic proximity, similarities of soil types, water content of the soils and the use-histories of the forest resources, while at the same time differentiating it from other fragments encountered in the lowland region.
It is therefore recommended that restoration efforts directed towards degraded areas in the central coastal region of the Baixada Fluminense include the planting of L. pubescens, Cabralea canjerana, Mabea fistulifera, Tapirira guianensis, and G. opposita. These taxa stand out in terms of their high VI in the survey fragment as well as in terms of their wide distribution in lowland regions, and their association with late-sucessional understory species. Their planting will help generate well diversified forests.
The characteristic fragmented condition of the central coastal plain and the northern portion of Rio de Janeiro State make these small fragments very relevant to the establishment of landscape restoration strategies and subsequent species conservation.
ACKNOWLEDGEMENTS
The authors thank PETROBRAS for financially supporting the "Programa Mata Atlântica" of the Rio de Janeiro Botanical Garden, including the present study; the Reserva Biológica de Poço das Antas/IBAMA for their logistic support; the Escola Nacional de Botânica Tropical (ENBT) for the use of their facilities; Antônio Tavares de Oliveira, Adilson Pintor and Jonas Alves Dias of the "Programa Mata Atlântica" for their help with the field work; the taxonomists of the Rio de Janeiro Botanical Garden, including Alexandre Quinet, Haroldo Cavalcanti de Lima, José Fernando A. Baumgratz and Marcelo Costa Souza; the owners of the Santa Helena Farm for allowing free access to the study area; the CAPES for the Masters grant to the first author; Roy Funch for the English version and its revision and Maurício Salazar Yepes for the Spanish version of the summary.
REFERENCES
1. APG (2003) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Bot. J. Linn. Soc. 141: 399-436. [ Links ]
2. Bernardes LNC (1952) Tipos de clima do estado do Rio de Janeiro. Rev. Bras. Geog. 1: 57-80. [ Links ]
3. Borém RAT, Ramos DP (2001) Estrutura fitossociológica da comunidade arbórea de uma toposseqüência pouco alterada de uma área de floresta atlântica, no município de Silva Jardim-RJ. Árvore 25: 131-140. [ Links ]
4. Borém RAT, Oliveira-Filho AT (2002) Fitossociologia do estrato arbóreo em uma topossequência alterada de Mata Atlântica, no município de Silva Jardim-RJ, Brazil. Árvore 26: 727-742. [ Links ]
5. Carvalho FA, Nascimento MT, Braga JMA (2006a) Composição e riqueza florística do componente arbóreo da Floresta Atlântica submontana na região de Imbaú, Município de Silva Jardim, RJ. Acta Bot. Bras. 20: 727-740. [ Links ]
6. Carvalho FA, Nascimento MT, Braga JMA, Rodrigues PJFP (2006b) Estrutura da comunidade arbórea da Floresta Atlântica de baixada periodicamente inundada na Reserva Biológica de Poço das Antas, Rio de Janeiro, Brazil. Rodriguesia 57: 503-518. [ Links ]
7. Carvalho FA, Nascimento MT, Braga JMA (2007) Estrutura e composição florística do estrato arbóreo de um remanescente de Mata Atlântica submontana no município de Rio Bonito, RJ, Brazil (Mata Rio Vermelho). Árvore 31: 717-730. [ Links ]
8. Christo AG, Guedes-Bruni RR, Fonseca-Kruel VS (2006) Uso de recursos vegetais em comunidades limítrofes à Reserva Biológica de Poço das Antas, Silva Jardim, RJ: estudo de caso na Gleba Aldeia Velha. Rodriguesia 57: 519-542. [ Links ]
9. Cunha SB (1995) Impactos das Obras de Engenharia sobre o Ambiente Biofísico da Bacia do rio São João (Rio de Janeiro-Brasil). Instituto de Geociências da UFRJ. Rio de Janeiro, Brazil. 378 pp. [ Links ]
10. EMBRAPA (1999) Sistema Brasileiro de Classificação de Solos. Embrapa Solos. Rio de Janeiro, Brazil. 412 pp. [ Links ]
11. Fantini AC, Guries RP (2007) Forest structure and productivity of palmiteiro (Euterpe edulis Martius) in the Brazilian Mata Atlântica. Forest Ecol. Manag. 242: 185-194. [ Links ]
12. Gandolfi S, Leitão-Filho HF, Bezerra CLF (1995) Levantamento florístico e caráter sucessional das espécies arbustivo-arbóreas de uma floresta mesófila semidecídua no município de Guarulhos, SP. Rev. Bras. Biol. 55: 753-767. [ Links ]
13. Guedes RR (1988) Composição florística e estrutura de um trecho de mata perturbada de baixada no município de Magé, Rio de Janeiro. Arq. Jard. Bot. Rio Jan. 29: 155-200. [ Links ]
14. Guedes-Bruni RR (1998) Composição, Estrutura e Similaridade Florística de Dossel em Seis Unidades de Mata Atlântica no Rio de Janeiro. Thesis. Universidade de São Paulo, Brazil. 175 pp. [ Links ]
15. Guedes-Bruni RR, Silva-Neto SJ, Morim MP, Mantovani W (2006a) Composição florística e estrutura de dossel em trecho de Floresta Ombrófila Densa Atlântica sobre morrote mamelonar na Reserva Biológica de Poço das Antas, Silva Jardim, Rio de Janeiro, Brazil. Rodriguesia 57: 429-442. [ Links ]
16. Guedes-Bruni RR, Silva-Neto SJ, Morim MP, Mantovani W (2006b) Composição florística e estrutura de trecho de Floresta Ombrófila Densa Atlântica Aluvial na Reserva Biológica de Poço das Antas, Silva Jardim, Rio de Janeiro, Brazil. Rodriguesia 57: 413-428. [ Links ]
17. Guedes-Bruni RR, Silva AG, Mantovani W (2009) Rare canopy species in communities within the Atlantic Coastal Forest in Rio de Janeiro State, Brazil. Biodiv. Cons. 18: 387-403. [ Links ]
18. IBDF/FBCN (1981) Plano de Manejo da Reserva Biológica de Poço das Antas. Instituto Brasileiro de Desenvolvimento Florestal/Fundação Brasileira para a Conservação da Natureza. Brasília, Brazil. 95pp. [ Links ]
19. IBGE (1992) Manual Técnico da Vegetação Brasileira. Ser. Manuais Técnicos em Geociências. Instituto Brasileiro de Geografia e Estatística. Rio de Janeiro, Brazil. 166pp. [ Links ]
20. Laurance WF, Delamônica P, Laurance SG, Vasconçelos HL, Lovejoy TE (2000) Rainforest fragmentation kills big trees. Nature 404: 836. [ Links ]
21. Lima HC, Pessõa SVA, Guedes-Bruni RR, Morães LFD, Granzotto SV, Iwamoto S, Di Ciero J (2006) Caracterização fisionômico-florística e mapeamento da vegetação da Reserva Biológica de Poço das Antas, Rio de Janeiro, Brazil. Rodriguesia 57: 369-389. [ Links ]
22. Lopes WP, Silva AF, Souza AL, Meira-Neto JAA (2002) Estrutura fitossociológica de um trecho de vegetação arbórea no Parque Estadual do Rio Doce - Minas Gerais, Brasil. Acta Bot. Bras. 16: 443-456. [ Links ]
23. Loss ACC, Silva AG (2005) Comportamento de forrageio de aves nectarívoras de Santa Teresa - ES. Natureza on line 3: 48-52. [ Links ]
24. Magurrán AE (1988) Ecological Diversity and its Measurement. Princeton University Press, Princeton, NJ, USA. 192 pp. [ Links ]
25. Melo MMRF (2002) Diagrama de Perfil. In Sylvestre LS, Rosa MMT (Eds.) Manual Metodológico para Estudos Botânicos na Mata Atlântica. Seropédica / Universidade Federal Rural do Rio de Janeiro. Brazil. 123 pp. [ Links ]
26. MMA (1998) Diretrizes para a Política de Conservação e Desenvolvimento Sustentável da Mata Atlântica. Caderno Nº 13. Ministério do Meio Ambiente. Brasília, Brazil. 30 pp. [ Links ]
27. Morães LFD, Assumpção JM, Luchiari C, Pereira TS (2006) Plantio de espécies arbóreas nativas para a restauração ecológica na Reserva Biológica de Poço das Antas, Rio de Janeiro, Brazil. Rodriguesia 57: 477-489. [ Links ]
28. Müeller-Dombois D, Ellenberg H (1974) Aims and Methods of Vegetation Ecology. Wiley. New York, USA. 262 pp. [ Links ]
29. Neves GMS (1999) Florística e estrutura da comunidade arbustivo-arbórea em dois remanescentes de Floresta Atlântica secundária - Reserva Biológica de Poço das Antas, Silva Jardim, RJ. Thesis. Universidade Federal do Rio de Janeiro. Brazil. 115 pp. [ Links ]
30. Oliveira-Filho AT, Fontes MAL (2000) Patterns of floristic differentiation among Atlantic Forests in southeastern Brazil and the influence of climate. Biotropica 32: 793-810. [ Links ]
31. Paula A, Silva AF, Souza AL, Santos FAM (2004) Sucessão ecológica da vegetação arbórea em uma Floresta Estacional Semidecidual, Viçosa, MG, Brazil. Acta Bot. Bras. 18: 407-423. [ Links ]
32. Pessõa SVA, Oliveira RR (2006) Análise estrutural da vegetação arbórea em três fragmentos florestais na Reserva Biológica de Poço das Antas, Rio de Janeiro, Brazil. Rodriguesia 57: 391-411. [ Links ]
33. Pielou EC (1975) Ecological Diversity. Wiley. New York, USA. 165 pp. [ Links ]
34. Scariot A, Freitas SR, Neto EM, Nascimento MT, Oliveira LC, Sanaiotti T, Servilha AC, Villela DM (2003) Efeitos da fragmentação sobre a biodiversidade: vegetação e flora. In Rambaldi DM, Oliveira DAS (Eds.) Fragmentação de Ecossistemas: Causas, Efeitos sobre a Biodiversidade e Recomendações de Políticas Públicas. MMA/SBF, Brasília, Brazil. 506 pp. [ Links ]
35. Shepherd GJ (1995) FITOPAC 1: Manual do Usuário. Universidade Estadual de Campinas. Brazil. 93 pp. [ Links ]
36. Siqueira MF (1994) Análise florística e ordenação de espécies arbóreas da Mata Atlântica através de dados binários. Thesis. Universidade Estadual de Campinas. Brazil. 143 pp. [ Links ]
37. Sneath PHA, Sokal RR (1973) Numerical Taxonomy. Freeman. San Francisco, CA. USA. 573 pp. [ Links ]
38. Tabarelli M, (1997) A Regeneração da Floresta Atlântica Montana. Thesis. Universidade de São Paulo. Brazil. 104pp. [ Links ]
39. Whittaker RH (1975) Communities and Ecosystems. MacMillan, New York, USA. 387 pp. [ Links ]
40. Zar JH (1999) Biostatistical Analysis. Prentice Hall. Upper Saddle River, NJ, USA. 663 pp. [ Links ]
