











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.34 n.9 Caracas sep. 2009
Análisis de la abundancia, dominancia y diversidad de la comunidad de peces demersales de profundidad de 90 a 540 metros en el Golfo de California, México
Alejandro Acevedo-Cervantes, Juana López-Martínez, Eloisa Herrera-Valdivia y Jesús Rodríguez-Romero
Alejandro Acevedo Cervantes. Biólogo y Maestro en Ciencias Marinas, Instituto Politécnico Nacional (IPN), México. Estudiante de Doctorado, Universidad Autónoma de Nayarit (UAN), México. Catedrático, Instituto Tecnológico de Guaymas, México.
Juana López Martínez. Doctora en Ciencias Marinas, IPN; México. Investigadora, Centro de Investigaciones Biológicas del Noroeste (CIBNOR), México. Dirección: Km. 2.35 Carretera a Las Tinajas, S/N Colonia Tinajas, Guaymas, Sonora, México. C. P. 85460. e-mail: jlopez04@cibnor.mx
Eloisa Herrera Valdivia. Bióloga y Maestría en Manejo de Zona Costera, Instituto Tecnológico de Guaymas, México. Técnico, CIBNOR, México.
Jesús Rodríguez Romero. Doctor en Ciencias, UAN, México. Investigador, CIBNOR, México.
RESUMEN
Se hace un análisis de la comunidad de los peces demersales del talud continental de la costa oriental del Golfo de California, México. Las muestras provienen de tres cruceros realizados con una red de arrastre a profundidades de 90-540m. Los diagramas de temperatura-salinidad (T-S) mostraron que a esas profundidades se encuentra la masa de agua del Golfo de California y por debajo de ella, la masa de agua sub-superficial del Pacífico. Se encontró que la temperatura y el O2 disuelto disminuyeron gradualmente hasta la profundidad máxima de muestreo (540m), siendo en esta última de 7°C y 0.1ml·l-1, respectivamente. Se registraron 71 especies pertenecientes a 35 familias. El valor de diversidad máxima y mínimo promedio (H) fue 2,2 y 0,8 bits/ind, y la riqueza específica (D) fue de 3,6 y 0,2 Las especies Caelorinchus scaphopsis, Merluccius productus, Pleuronichthys verticals, Sebastes spinorbis, Microlephidium verecundum, Lepophidium prorates y Lophiodes spilurus fueron las dominantes de acuerdo al índice de importancia relativa. La comunidad de peces del talud se conformo de dos comunidades diferentes, la primera por especies de ambito costero que abarca de 90 a 270m, caracterizada por valores moderados y altos de diversidad y riqueza especifica, con la dominancia compartida entre un mayor número de especies, mientras que la segunda comunidad estuvo formada por especies típicas de profundidad, de baja diversidad, y la dominancia esta soportada en pocas especies. Se infiere que las variables ambientales juegan un papel importante en la conformación de esta estructura comunitaria.
Abundance, dominance and diversity analysis of the demersal fish community analysis at 90 to 540 meters of depth in the Gulf of California, Mexico
SUMMARY
An analysis of the demersal fish community of the eastern continental slope of the Gulf of California, México, was performed. Samples were obtained with a trawl net at 90-540m of depth by three separate research surveys. Temperature-salinity diagrams showed two main water masses, one represented by the Gulf of California water mass, down to a depth of 540m, and a deeper one corresponding to the sub-surface Pacific water mass. Depth profiles for temperature and dissolved O2 were obtained, and plots constructed from them. These profiles showed that temperature and O2 concentration decreased gradually along a smooth gradient to a minimun of 7°C and 0.1ml·l-1, respectively, at 540m deep. Seventy-one species belonging to 35 families were found. Average maximum and minimum overall diversity (H´) values were 2.2 and 0.8 bits/ind and the specific richness (R) index value were 3.6 and 0.2. Caelorinchus scaphopsis, Merluccius productus, Pleuronichthys verticals, Sebastes spinorbis, Microlephidium verecundum, Lepophidium prorates. and Lophiodes spilurus were the most dominant species, based on the relative importance index. The fish community structure suggests two different communities with two groups of species, one at 90-270m deep that shows strong associations with moderate to high diversity values and specific richness, with shared dominance by numerous species, and a second community made out of typical deep water species, with low diversity and dominance featured by a few species. The data suggests that environmental conditions play an important role in the distribution and the community structure of these species.
Análise da abundância, dominância e diversidade da comunidade de peixes demersais de profundidade de 90 a 540 metros no Golfo da Califórnia, México
RESUMO
Faz-se uma análise da comunidade dos peixes demersais do talude continental da costa oriental do Golfo da Califórnia, México. As amostras provêm de três cruzeiros realizados com uma rede de arrasto a profundidades de 90-540m. Os diagramas de temperatura-salinidade (T-S) mostraram que a essas profundidades se encontra a massa de água do Golfo da Califórnia e por baixo dela, a massa de água subsuperficial do Pacífico. Encontrou-se que a temperatura e o O2 dissolvido diminuiram gradualmente até a profundidade máxima de amostragem (540m), sendo nesta última de 7°C e 0.1ml·l-1, respectivamente. Registraram-se 71 espécies pertencentes a 35 famílias. O valor de diversidade máximo e mínimo médio (H) foi 2,2 e 0,8 bits/ind, e a riqueza específica (D) foi de 3,6 e 0,2. As espécies Caelorinchus scaphopsis, Merluccius productus, Pleuronichthys verticals, Sebastes spinorbis, Microlephidium verecundum, Lepophidium prorates e Lophiodes spilurus foram as dominantes de acordo ao índice de importância relativa. A comunidade de peixes do talude foi conformada a partir de duas comunidades diferentes, a primeira por espécies de âmbito costeiro que abrange de 90 a 270m, caracterizada por valores moderados e altos de diversidade e riqueza específica, com a dominância compartida entre um maior número de espécies, enquanto que a segunda comunidade esteve formada por espécies típicas de profundidade, de baixa diversidade, e a dominância está suportada em poucas espécies. Infere-se que as variáveis ambientais têm um papel importante na conformação desta estrutura comunitária.
PALABRAS CLAVE / Borde Continental / Demersal / Golfo de California / Peces de Aguas Profundas /
Recibido: 13/11/08. Aceptado: 03/09/2009.
Introducción
La denominación de peces de aguas profundas se aplica a aquellas especies de peces que pasan todo su ciclo de vida por debajo de la zona fótica del océano. Los ecosistemas y los recursos pesqueros de profundidad son altamente vulnerables a la explotación debido a que poseen alta longevidad, crecimiento lento, madurez sexual tardía, baja fecundidad (son estrategas K), una baja resiliencia y pueden ser altamente vulnerables a la sobrepesca (Koslow et al., 2000).
A nivel mundial, algunas comunidades de peces demersales de aguas profundas son objeto de pesca comercial (Koslow et al., 2001; Gordon et al., 2003) y en muchos casos el conocimiento sobre ellas es escaso, requiriéndose información a nivel de ecosistema y biología poblacional, con la finalidad de llegar a un manejo sustentable de estos recursos (Koslow et al., 2001; Gordon et al., 2003; Fossen et al., 2008).
Los estudios acerca de peces de profundidad son escasos y entre ellos la matoría son estudios realizados en el océano Atlántico (Large et al., 2003; Gordon et al., 2003; Powell et al., 2003). Son pocos los trabajos sobre la estructura comunitaria de los peces de profundidad en el Pacifico Oriental (Arana et al., 2002; Reyes, 2007) y para el Golfo de California son aun más escasos. En particular en el Golfo de California, existe información de síntesis del conocimiento de los peces de aguas profundas, principalmente de especies bentopelágicas y pelágicas (Castro-Aguirre y Balart, 1996); sin embargo, se desconoce la estructura de las comunidades a esas profundidades.
La idea que los fondos marinos son zonas donde las especies son escasas por las condiciones ambientales no es del todo cierta (Koslow et al., 2000). Existe controversia sobre el grado de diversidad en ambientes marinos profundos, habiéndose planteado una alta diversidad por algunos autores (Grassle, 1989; Castro-Aguirre y Balart, 1996; Grant, 2000) y sugiriéndose diversos mecanismos para explicar esa alta diversidad, tales como el efecto de la interacción de competencia y depredación (Grant, 2000) y la historia evolutiva de los propios ecosistemas (Castro-Aguirre y Balart, 1996). Así mismo, se ha planteado que la diversidad varia según los tipos de sedimentos, ya que de ello depende la disponibilidad de alimento determinante en la abundancia de especies (Gage y Tyler, 1991). Se ha propuesto que existe una clara diferencia entre la fauna de la plataforma continental y del talud continental, y que las variables que influyen en el cambio de las comunidades de peces de profundidad son difíciles de establecer (Powell et al., 2003), aludiéndose gradientes de factores como nivel de luz, sustrato y sedimentos, así como la temperatura, oxígeno disuelto y la salinidad (Fujita et al., 1995).
Los trabajos que relacionan la composición de las especies en aguas profundas con parámetros ambientales y características del ambiente son escasos en México, por lo cual el objetivo del presente trabajo fue determinar la estructura funcional en base a la composición, abundancia, diversidad y dominancia en las comunidades de peces en la porción oriental del Golfo de California entre Puerto Peñasco, Sonora, y Topolobampo, Sinaloa.
Material y Métodos
Se obtuvieron muestras de peces provenientes de tres cruceros realizados dentro del Golfo de California (GC), en septiembre 2004, febrero 2005 y mayo 2005. Los lances se efectuaron en la costa oriental del GC desde el sur de Puerto Peñasco, Sonora, hasta Topolobampo, Sinaloa, siguiendo el derrotero mostrado en la Figura 1. Los equipos de pesca utilizados fueron una red de arrastre de fondo construida en material de polietileno trenzado, de 38´34m y perímetro de la boca en paño estirado de 68m, con una relinga superior de 38m. La red operó por popa a profundidades de 90-540m. De cada lance se registro el total de la captura y se tomó una muestra de 20kg que fue conservada en congelación hasta su posterior análisis en el laboratorio de especialidades de pesquerías del CIBNOR. En cada lance de la red se registró la profundidad, temperatura, salinidad y O2 disuelto por medio de una sonda (CTD).
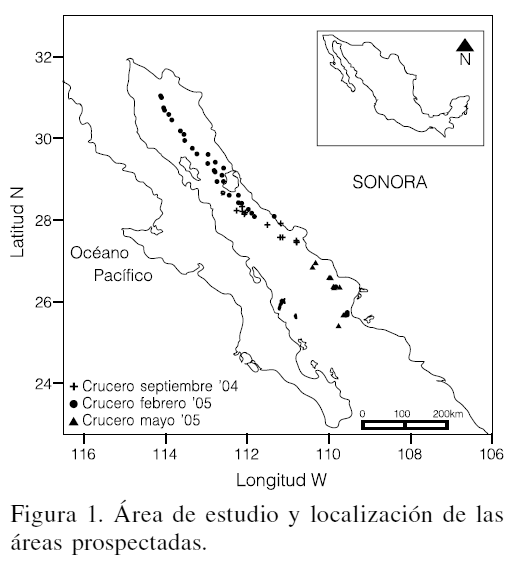
De la muestra tomada, los peces fueron separados, se determinó la especie y se efectuaron muestreos biológicos consistentes en mediciones de longitud total (mm), longitud patrón (mm) y peso. La determinación taxonómica fue efectuada mediante las claves de Fisher et al. (1995). Se valoró la abundancia relativa (Rodríguez-Romero et al., 1994), la diversidad a través del índice de Shannon-Weaver (Brower y Zar, 1977), la riqueza específica (Margaleff, 1969), la equitatividad (Brower y Zar, 1977), la dominancia mediante el Índice de valor biológico (IVB) de Sanders (Loya y Escofet, 1990) y el índice de importancia relativa (IIR, Pinkas et al., 1971). Con el fin de analizar la distribución de la diversidad y la dominancia por niveles de profundidad, se consideró formar estratos de profundidad cada 90m. Si bien se efectuaron lances entre las profundidades de 360 y 450m, no fue posible obtener lances positivos (con captura) a esas profundidades.
Resultados
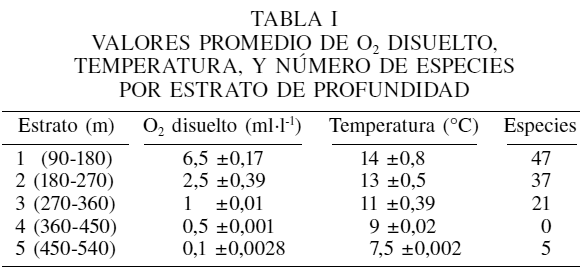
El número de especies, así como los valores de las variables ambientales registrados se presentan, por estrato de profundidad, en la Tabla I. De acuerdo con los diagramas de temperatura-salinidad (T-S; Figura 2), estos parámetros corresponden a dos masas de agua: masa de agua del Golfo de California y masa de agua sub-superficial del Pacífico (Lavin et al., 1997).
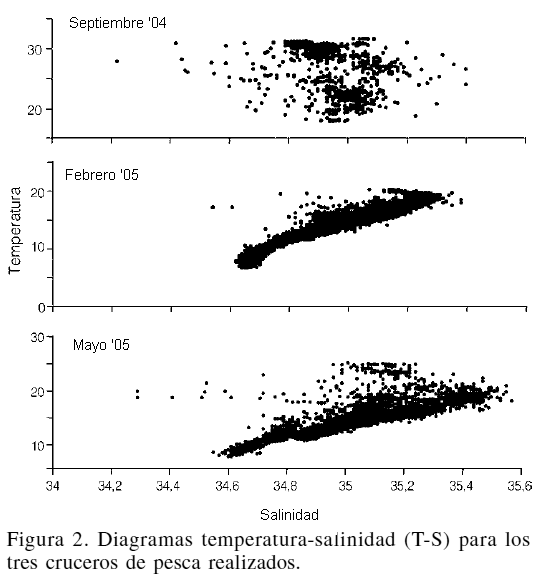
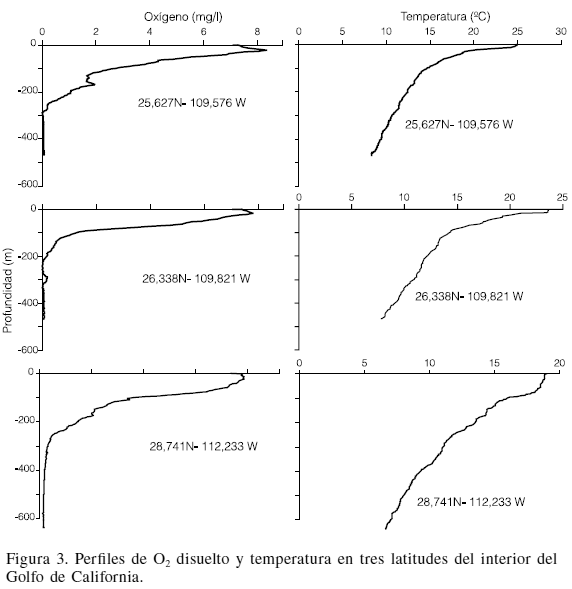
La concentración de O2 disuelto varió desde 8ml·l-1 en la superficie hasta ~0,1ml·l-1 a partir de los 450m de profundidad. En la región norte del GC, donde se alcanzó a medir profundidades de 340m, el valor mínimo fue de 2ml·l-1, mientras que en región central y sur del GC fue <0,5ml·l-1. La temperatura también disminuyó rápidamente desde 14ºC a 90m hasta 7,5ºC a 450m (Figura 3).
Se colectaron un total de 9898 peces, con una biomasa total de 470,1kg y que correspondieron a 71 especies (Tabla II) pertenecientes a 35 familias. En los cruceros de septiembre 04, febrero 05 y mayo 05 fueron capturados especimenes de 33, 47 y 49 especies, respectivamente. Las familias mas importantes fueron Paralichthydae (8 especies), Serranidae (6), Scorpaenidae y Triglidae (5), Ophidiidae y Scianidae (4) y Moridae (3 especies).
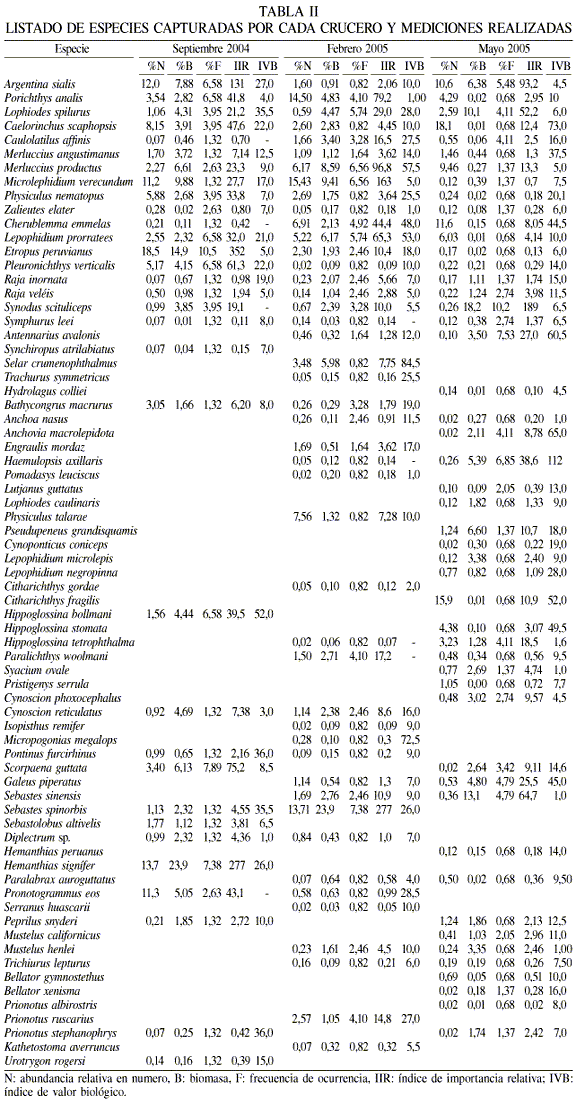
De acuerdo al índice de abundancia relativa (IAR), 19 especies aportaron las mayores abundancias. En septiembre, 7 especies acumularon el 71%, en febrero 6 acumularon el 56%, y para mayo 6 aportaron el 71% (Tabla II). Así se tiene que Etropus peruvianus (18%) es el más importante en septiembre, Sebastes spinorbis (14%) en febrero, y Caelorhinchus scaphopsis (17%) y Citharhichthys fragilis (16%) en mayo. El IAR en peso mostró que para septiembre 10 especies acumularon el 70% de la biomasa, mientras que en febrero 6 especies aportaron el 59% y en mayo 7 acumularon el 64%. Hippoglossina bollmani fue la que aportó mayor peso en septiembre, con un 14%, Sebastes spinorbis en febrero con 23.9% y en mayo Synodus scituliceps con 18.2%.
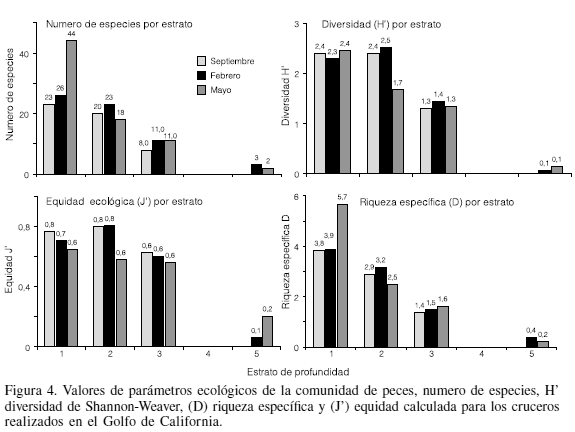
El número de especies registradas por estrato disminuyó conforme aumento la profundidad. El estrato de profundidad que mayor número de especies presentó fue el de 90-180m, destacando el mes de mayo, cuando se capturaron 44 especies, mientras que el estrato con menor número de especies fue el de 450-540m con 5 especies.
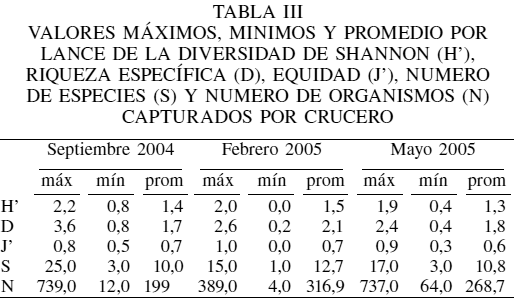
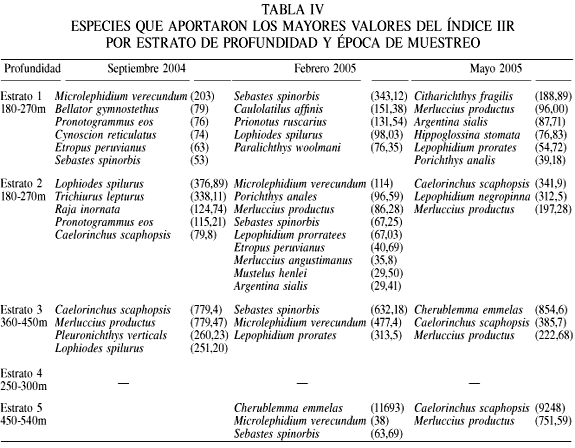
La diversidad calculada mediante el índice de Shannon (H), riqueza específica (D) y la equidad (J) se muestran en la Tabla III. La diversidad presentó valores entre 0,8 y 2,2 bit/ind, que corresponden a diversidades medias, contrario a lo que se cree comunmente, y mostrando una tendencia a disminuir conforme aumenta la profundidad, siendo los estratos de 90-180 y 180-270 los que presentaron los mayores valores (Figura 4). La equidad J presento valores entre 0,6 a 0,85 en los primeros tres estratos, los cuales son consistentes con los valores de diversidad arriba mencionados. El estrato de los 450-540 presentó los menores valores (0,2), mostrando una alta dominancia en la abundancia de unas pocas especies. Se encontró que en estratos de mayor profundidad son pocas las especies que muestran ser dominantes, siendo la dominancia específica estimada a través del índice de importancia relativa (Tabla IV).
Discusión
Este trabajo proporciona los primeros datos de la comunidad de peces del talud continental de la costa este del GC, a profundidades mayores a 90m, de la cual no se tiene referencia previa. De las familias encontradas, Paralichthydae, Serranidae, Scorpaenidae, Scianidae y Triglidae aportaron mayor número de especies. Todas ellas son comúnmente encontradas en la región costera de todos los mares del mundo; sin embargo, se han reportado como componentes de la fauna demersal de la parte externa de la plataforma continental (Castro-Aguirre y Balart, 1996). De las 71 especies registradas, las 19 que presentaron las mayores abundancias (Tabla II) son típicas habitantes de la zona profunda en el GC y sus registros son escasos. Los estratos de 90-180 y 180-270m de profundidad, además de presentar el mayor número de especies, presentaron solamente especies de distribución costera, tanto de la plataforma continental, como del talud continental (Powell et al., 2003). La disminución en el número de especies conforme aumenta la profundidad se puede explicar por el hecho que las condiciones ambientales (disminución de la temperatura y de la concentración de O2 disuelto) cambian drásticamente a mayor profundidad, haciendo el ambiente menos apropiado para sostener la vida y por ello solo se presentan especies capaces de soportar condiciones frías y casi anóxicas. En el GC existe una capa de agua con concentración baja de O2 (0,2-0,6ml·l-1) denominada zona de oxígeno mínimo, la cual afecta la distribución y abundancia de los organismos (Hendrickx, 2001; Lluch-Cota et al., 2007).
Los valores de diversidad de Shannon-Wiener y de riqueza específica encontrados fueron valores medios, sobre todo en el estrato de 90-180m, hecho que coincide con lo reportado por Grassle (1989) y Grant (2000). La disminución de la diversidad con el aumento en la profundidad puede ser reflejo de las condiciones estresantes de estos ambientes, haciendo que la dominancia de las especies adaptadas a condiciones anóxicas se incremente.
De acuerdo al patrón estructural hallado en el presente estudio, en los estratos más someros se registraron especies más costeras y con una mayor diversidad, presentándose así una comunidad diferente en esos estratos. La ictiofauna costera es un componente importante de los ambientes marinos costeros e influye en gran medida en la alta diversidad de estos ecosistemas. Una parte importante de especies de peces costeros que incursionan o realizan migraciones verticales sobre el talud continental propician que la diversidad sea mayor en los estratos de 90-270m de profundidad, constituyendo por lo tanto un ecotono, tal como lo documentó Margaleff (1991), zona de transición natural entre dos ecosistemas distintos. Powell et al. (2003) mencionan que la diversidad varia dependiendo de la profundidad y que tanto la diversidad como la riqueza específica son mayores en la parte alta del talud continental y menores en la parte baja del miamo; sin embargo, Fossen et al. (2008) muestran valores de diversidad altos a profundidades de 1000 a 2000m, situación que no se refleja en la tendencia a disminuir de la diversidad en este estudio, tal vez porque los muestreos efectuados en este trabajo no llegaron mas allá de la zona de oxigeno mínimo, donde se sabe que se incrementan nuevamente los niveles de O2 (Hendrickx y Hastings, 2007; Lluch-Cota et al., 2007).
Los resultados de diversidad y dominancia sugieren la presencia de dos comunidades diferentes, la primera formada por especies de ámbito costero y que esta bien adaptada a condiciones de temperatura y concentración de O2 bajas, la cual se ubica entre los 90 y 270m de profundidad, considerada como la zona superior del talud continental, caracterizada por una alta diversidad y pocas especies dominantes; mientras que la segunda comunidad se ubica a mayor profundidad, en la zona media y profunda del talud continental, y cuya principal característica es la baja diversidad y la presencia de pocas especies dominantes con familias típicas de aguas profundas. Entre las dos comunidades existiría un ecotono compuesto principalmente por especies migratorias. Se sugiere que los factores ambientales, en especial la concentración de O2, tienen un efecto importante en la distribución y composición estructural de estas especies,
AGRADECIMIENTOS
Los autores agradecen el apoyo del personal técnico del Laboratorio de Pesquerías de la Unidad Guaymas del CIBNOR, México, del Centro de Investigaciones Biológicas del Noroeste, S.C., donde se llevo a cabo la investigación, y el proyecto SAGARPA-CONACYT 2003 002-024.
REFRENCES
1. Arana PE, Barrios M, Moraga J, Mujica A, Alarcón R (2002) Evaluación Directa del Camarón Nailon entre la II y VIII Regiones, 1999. Informe Final. FIP. 99-08 Universidad Católica del Norte. Coquimbo, Chile. 208 pp. [ Links ]
2. Brower JE, Zar HJ (1977) Field and Laboratory Methods for General Ecology. Brown. Dubuque, IO, EEUU. 194 pp. [ Links ]
3. Castro-Aguirre JL, Balart EF (1996) Contribución al conocimiento del origen y las relaciones de la ictiofauna de aguas profundas del Golfo de California, México. Hidrobiológica 6: 67-76. [ Links ]
4. Fisher W, Krupp F, Schneider W, Sommer C, Carpenter KE, Niem VH (1995) Guía FAO para la Identificación de Peces para los Fines de Pesca. Pacífico Centro-oriental. Vol. II y III. Vertebrados. Partes 1 y 2. pp. 647-1813. [ Links ]
5. Fossen I, Cotton CF, Bergstad OA, Dyb JE (2008) Species composition and distribution patterns of fishes captured by long-lines on the mid-Atlantic ridge. Deep Sea Res. 55: 203-217. [ Links ]
6. Fujita T, Inada T, Ishito Y (1995) Depth gradient structure of the demersal fish community on the continental shelf and upper slope off Sendai Bay, Japan. Mar. Ecol. Prog. Ser. 118: 13-23. [ Links ]
7. Gage JD, Tyler PA (1991) Deep Sea Biology: A Natural History of Organisms at the Deep Sea Floor. Cambridge Univesity Press. Cambridge, RU. 504 pp. [ Links ]
8. Gordon JDM, Bergstad AO, Figueiredo I, Menezes G (2003) Deep water fisheries of the Northeast Atlantic: I. Description and current Trends. J. Northw. Atl. Fish. Sci. 31: 137-150. [ Links ]
9. Grant A (2000) Deep sea diversity: Overlooked messages from shallow waters sediments. Mar. Ecol. 21: 97-112. [ Links ]
10. Grassle JF (1989) Species diversity in deep-sea communities. Trends Ecol. Evol. 4: 12-15. [ Links ]
11. Hendrickx ME, Hastings PA (2007) Información ecológica de Myxine circifrons Garman, 1899 (Myxiniformes: Myxinidae) en el Golfo de California, México. Hidrobiológica 17: 273-276. [ Links ]
12. Hendrickx ME (2001) Occurrence of a continental slope deep-water decapod crustacean community along the edge of the minimum oxygen zone in the southeastern Gulf of California, México. Belg. J. Zool. 131: 71-86. [ Links ]
13. Koslow JA, Boehlert GW, Gordon JDM, Haendrich RL, Lorance P, Parin N (2000) Continental slop and deep-sea fisheries: implications for a fragile ecosystem. ICES J. Mar. Sci. 57: 548-557. [ Links ]
14. Koslow JA, Gowlett-Holmes K, Lowry JK, O'Hara T, Poore GCB, Williams A (2001) Seamount benthic macrofauna off southern Tasmania: community structure and impacts of trawling. Mar. Ecol. Progr. Ser. 213: 111-125. [ Links ]
15. Large PA, Hammer C, Bergstad AO, Gordon JDM, Lorance P (2003) Deep-water fisheries in the Northeast Atlantic: II Assessment and Management Approaches. J. Northw. Atl. Fish. Sci. 31: 151-163. [ Links ]
16. Lavin MF, Beber E, Badan A (1997) Estructura hidrográfica y circulación del Golfo de California: Escala estacional e interanual. En Contribuciones a la Oceanografía Física en México. Monografía Nº 3. Unión Geofísica Mexicana. pp. 141-171. [ Links ]
17. Lluch-Cota SE, Aragón-Noriega EA, Arreguín-Sánchez F, Aurioles-Gamboa D, Bautista-Romero JJ, Brusca RC, Cervantes-Duarte R, Cortés-Altamirano R, Del-Monte-Luna P, Esquivel-Herrera A, Fernández G, Hendrickx ME, Hernández-Vázquez S, Herrera-Cervantes H, Karu M, Lavín M, Lluch-Belda D, Lluch-Cota DB, López-Martínez J, Marinone SG, Nevárez-Martínez MO, Ortega-Garcia S, Palacios-Castro E, Parés-Sierra A, Ponce-Díaz G, Ramírez-Rodríguez M, Salinas-Zavala CA, Schwartzlose RA, Sierra-Beltran AP (2007) The Gulf of California: Review of ecosystem status and sustainability challenges. Progr. Oceanogr. 73: 1-26. [ Links ]
18. Loya-Salinas DH, Escofet A (1990) Aportaciones al cálculo del Índice de Valor Biológico (Sanders, 1960). Cien. Mar. 16: 97-115. [ Links ]
19. Margaleff DR (1969) Perspectives in Ecological Theory. Chicago University Press. Chicago, IL, EEUU. 111 pp. [ Links ]
20. Margaleff DR (1991) Ecología. 1a ed. Omega. Barcelona, España. 951 pp. [ Links ]
21. Pinkas L, Oliphant M, Iverson ILK (1971) Food habits of albacore, bluefin tuna and bonito in Californian waters. Calif. Fish Game 152: 1-105. [ Links ]
22. Powell SM, Haendrich RL y McEachran JD (2003). The deep demersal fish fauna of the Northern Gulf of Mexico. J. Northw. Atl. Fish. Sci. 31: 19-33. [ Links ]
23. Reyes PR (2007) Peces abisales de la "Triple Unión", lugar de contacto de las placas Antártica, Sudamericana y Nazca (archipielago patagónico noroccidental). Rev. Biol. Mar. Oceanogr. 42: 37-47. [ Links ]
24. Rodríguez-Romero J, Abitia-Cárdenas LA, Galván-Magaña F y Chávez-Ramos H (1994) Composición, abundancia y riqueza específica de la ictiofauna de Bahía Concepción, Baja California Sur, México. Cien. Mar. 20: 321-350. [ Links ]
