











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInvestigación Clínica
versión impresa ISSN 0535-5133versión On-line ISSN 2477-9393
Invest. clín v.51 n.4 Maracaibo dic. 2010
Human hepatoma cell line (Hep G2) cellular response to hypothermic stress with recovery. Induction of Hsp70, Hsp60 and Hsf1 expression.
Alegna Rada 1, Elizabeth Merentes 2, Marianela Rodríguez 1 Guillermo Anselmi 1 and Mirian Strauss 1
1 Sección de Biología Celular, Instituto de Medicina Tropical, Facultad de Medicina y
2 Laboratorio de Cultivo de Tejidos y Biología de Tumores, Facultad de Ciencias. Universidad Central de Venezuela. Caracas, Venezuela.
Corresponding author: Mirian Strauss. Sección de Biología Celular, Instituto de Medicina Tropical, Facultad de Medicina, Universidad Central de Venezuela. Apdo 47019. Caracas 1041A, Venezuela. Phone: 00-58-212- 6053650, Fax: 00-58-212-2434685. E-mail: mstraussve@gmail.com.
Abstract.
The cell response of human HepG2 cells exposed to hypothermia with rewarming was analyzed. Ultrastructural findings in hypothermic stressed cells showed swollen mitochondria, dispersed chromatin, vacuoles and ring-shape nucleolar reorganization. These changes were coupled with significative differences in the induction of Hsp60, inducible Hsp70 and monomeric Hsf1 in all treated samples, but not in Hsc 70 expression. Cellular response to hypothermia could be associated with the synergistic induction of Hsp expression.
Key words: heat shock proteins, ultrastructural damage, hypothermia with recovery, HepG2 cells.
Respuesta celular de línea de hepatoma humano (HepG2) al estrés hipotérmico con recuperación. Inducción de la expresión de Hsp60, Hsp70, y Hsf1.
Resumen.
En este trabajo se analizó la respuesta celular de células HepG2 expuestas a hipotermia con posterior recuperación. Los hallazgos ultraestructurales en células sometidas a estrés hipotérmico incluyeron mitocondrias edematizadas, núcleos picnóticos, vacuolas y reorganización nucleolar en forma de anillo. Tales cambios están relacionados con diferencias significativas en la inducción de la expresión de Hsp60, Hsp70 inducible y Hsf 1 monomérico en todas las muestras tratadas, pero no de Hsc70. La respuesta celular a la hipotermia puede ser relacionada con la inducción sinergística de las Hsp.
Palabras clave: proteínas de shock térmico, daño estructural, hipotermia con recuperación, células HepG2.
Received: 30-11-2009. Accepted: 06-05-2010.
INTRODUCTION
Thermal stress stimulates a sort of complex responses which are fundamentals in the preservation of cell survival (1). Particularly in mammals, exposure to hypothermia or hyperthermia has been related to morphological and physiological modifications. While tissue exposure to hypothermia has been considered a strategy of preservation and protection (3, 4) a number of evidences of cell damage have been described in mammals exposed to hypothermic stress as a result of the alteration of homeostasis. Such physiological modifications (2) include losing the integrity of plasma membrane (3), cell breakage, edema, presence of dispersed chromatin, as well as chromatin condensation (4, 5), decrease of enzymatic rate, losing calcium and coagulation homeostasis, free radical provoked-damage, cytoskeleton and plasma membrane alterations, cell growth delay and cell death (6). Besides the modifications previously described as a result of thermal stress, cells respond by inducing the transient expression of heat shock proteins (Hsps) or stress proteins. These proteins participate in numerous functions including folding of newly synthesized proteins, transport of proteins into cell compartments, disaggregation of protein complexes and others functions (7).
The induction of the Hsps expression is mediated by the induction of the heat shock transcription factors (HsFs) through binding to the heat shock element (HSE) in the promoter region of the Hsp genes. The member of the Hsf family, Hsf1 is involved in the Hsps induction owed to heat shock and other kinds of stress (8-10). The mechanism of Hsf1 is regulated by phosphorylation at the post-translational level or interaction with other proteins. Upon a stressful condition the Hsf monomer is translocated into the nucleus and trimerizes. The trimer is able to bind the HSE in the promoter region of Hsp genes (11).
In human HepG2 cells, sublethal hypothermia induces an increased Hsp70 expression and accumulation. In addition its localization follows a stress dependent pattern in these cells by an unknown mechanism (12). Although little is known about the molecular mechanism implied in the induction of Hsps during hypothermia, the expression and accumulation of different Hsps could be linked with a synergistic network of mechanisms related to the stabilization of protein homeostasis. Indeed, the inductions of Hsp60 and Hsp90 in mouse adipose tissue (13) and Hsp90 in human keratinocytes have been described (4). This work studied the expression of Hsps in a human hepatoma cell line (HepG2) treated under hypothermic conditions, followed by rewarming together with the possible subcellular damage associated.
MATERIALS AND METHODS
Cell culture
Human hepatoma HepG2 cell line kindly provided by Dr. Antonio De Maio from the University of California, San Diego, USA, was used in all experiments. The cells were maintained in Dulbecco’s Modified Eagle Medium (Invitrogen), supplemented with heat inactivated 10% fetal bovine serum (Invitrogen), in a humidified atmosphere of 5% CO2 at 37°C.
Thermal treatment
The cells were incubated at 4°C during 40 min, 1 or 2 hours to assess the hypothermic stress condition. Then cells were transferred back to a 37°C incubator during 1 for rewarming while control cells were maintained at 37°C.
Ultrastructure methods
Cell samples exposed to 4°C during 1h and recovered at 37°C 1h, where the highest induction of Hsp70 and Hsp60 was observed, were analyzed by transmission electron microscopy. The cells were washed with Millonig buffer and centrifuged at 1200 rpm during 5 minutes, fixed in Karnovsky (320 mosmol, pH 7.4, 2 h, 4°C) and post-fixed in presence of osmium tetroxide (2% osmium tetroxide in Milloning buffer 0.12 M, 320 mosmol, pH 7.4, 2 h, 4°C). The samples were subsequently dehydrated by incubation in increasing concentrations of acetone (50%, 70%+uranyl, 80%, 95%, 100%) during 20 minutes and embedded in Araldyte epoxic resin and, before polymerization, at 60° during 48h. Thin sections of 100 mm were cut in a Reichert Om U3 ultramicrotome and counterstained with uranyl acetate (45 min, 60°C) and lead citrate (3 min, 25°C). The sections were analyzed in a transmission electron microscope Hitachi H-300, 75 kV.
Western blot analysis
Control and thermally stressed cells were lysed in a buffer containing 10 mM Tris-HCl pH 7.4, 0.1M EDTA, 2 mM PMSF, 10mM NaCl and 0.5% Triton X-100 at 4°C. The lysate was clarified by centrifugation at 14000 r.p.m. for 10 min at 4°C. Equal amounts of proteins (8.3 µg), determined by Bradford microassay using albumin as a standard, were boiled in SDS sample buffer (10mM Tris-HCl, pH 6.8, 2% SDS, 10% 2-bmercaptoethanol, 10% glycerol and bromephenol blue) during 5 min. Samples of equal protein loading were separated by SDS-PAGE (Laemmli, 1970) in duplicated gels using a Bio-Rad mini-gel system, one of them was stained with Coomassie Brilliant Blue G-250 (Bio-Rad Laboratories) while the second gel was blotted onto a nitrocellulose membrane (0.45µm, Bio-Rad Laboratories) and stained with Ponceau Red before inmunoblotting. Nonspecific protein binding sites were blocked by incubation in PBS pH 7.4; 0.1% Tween 20 and 5% skimmed milk. To examine Hsps expression the samples were incubated with the following antibodies (1:5000) Anti-Actin (rabbit polyclonal, Sigma, St. Louis, USA), anti-Hsp70 (rabbit polyclonal, Stressgen Bioreagents, Ann Arbor, USA), anti-Hsc 70 (rabbit polyclonal, Stressgen Bioreagents, Ann Arbor, USA) anti-Hsp60 (mouse monoclonal, Sigma, St. Louis, USA) and anti-monomeric Hsf1 (rabbit polyclonal anti-mouse, Laboratorio de Bioquímica de Parásitos-IMT-UCV, Caracas, Venezuela). Actin was used as a loading control. After washing several times with PBS-0.1% Tween 20, the membrane was incubated with 1:5000 horseradish peroxidase-conjugated rabbit anti-mouse IgG Peroxidase (Sigma, St. Louis, USA) or goat Anti-rabbit IgG (Pierce, Rockford, USA). Immunoreactive bands were detected by an enhanced chemiluminescence detection kit (Pierce) and exposed to Kodak X-ray film for 10 s. Protein immunoblots were scanned by GS-800 Bio-Rad Densitometer using the Multi-Analyst program (Bio-Rad).
Statistical analysis
Relative optical density values of each protein compared to actin from Western blot analysis are expressed as mean ± SEM of three independent experiments. The statistical significance of the differences between values was assessed by one-way Analysis of Variance (ANOVA) followed by Duncan multiple comparison post hoc test. A P<0.05 was considered to be statistically significant. Statistical analysis was performed using Statistica V.6.0 (StatSoft, Tulsa, USA).
RESULTS
A variety of morphological changes are provoked by hypothermia.
The ultrastructural findings included controls resembling normal appearance (Fig. 1a,b). In thermally treated samples (4°C,1h and 37°C, 1h; Fig. 1 c-f) lipid drops were not observed. In contrast, generalized edema (c-f) dispersed chromatin (c,f), swollen mitochondria (c-f) and autophagic vacuoles (e) were distinguished giving the appearance of necrotic cells, particularly in d and e. Ring-shape reorganization was observed (d,f) in the nucleolus of the thermal treated samples.
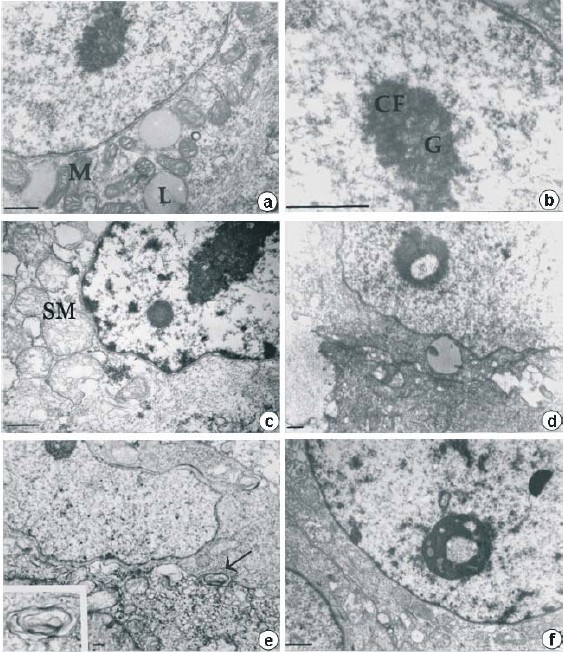
Fig. 1. Transmission electron micrographs of human HepG2 cells. Control (a, b) and hypothermically stressed cells with recovery (c-f). There are evident lipids drops (L) and abundant mitochondria in the perinuclear region (M). The typical nucleolus structure with the fibrillar dense (CF) and the granular (G) components is easily distinguished in b. In the cells under hypothermia 4°C during 1h and recovered at 37°C 1h some changes are evident: generalized edema (c-f), picnotic), swollen mitochondria (SM, c-f), as well as autophagic vacuoles (e, arrow and insert) are distinguished. Picnotic nucleus (c,f) were present and in the nucleolus of the thermal treated samples the ring-shape reorganization was observed (d, f). Bar=0, 5ìm.
Hypothermia induces Hsp60, the inducible Hsp70 and monomeric Hsf1 but not Hsc 70 expression. Western blot analysis (Fig. 2) and semiquantitative densitometry (Fig. 3) showed that Hsp60, Hsp70 and Hsf1 expression was increased in all treated samples, in contrast to control group. The cells treated hypothermically during 1h with 1h of recovery, showed the higher increase of Hsp60 and Hsp70. In the case of Hsf1, the denser band belonged to the group treated during 2h with 1h of recovery. The lower levels of expression were observed in Hsc70, while Hsp70 and Hsf1 were the most induced. As ANOVA showed, no significative changes in Hsc70 were observed between treatments. In the case of Hsp60, Hsp70 and Hsf1 the relative D.O. difference was significative to each treated sample compared to the control group (P<0.05).
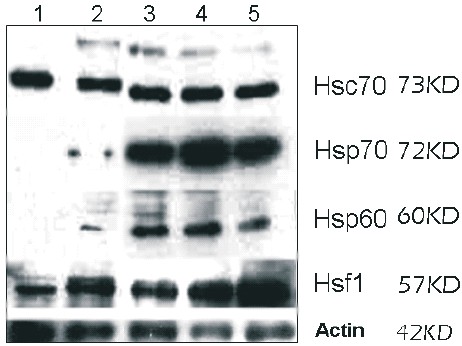
Fig. 2. Hsc70, Hsp70, Hsp60, Hsf1 and actin expression of hypothermically stressed and recovered human HepG2 cells. Western blot analysis of cells maintained at 37°C (1,2); hypothermically treated 4°C, 40 min (3), 1h (4), 2h (5). Thermal stressed cells were exposed to 37°C, 1h after treatment to rewarming. Relative molecular mass is indicated on the right. Actin was used as a loading control.
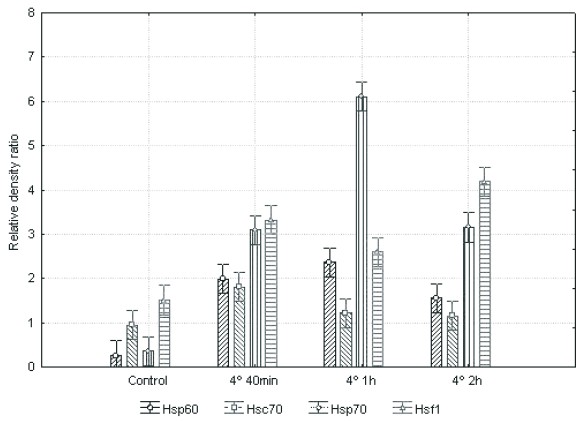
Fig. 3. Densitometric analysis of the bands of Hsp60, Hsc70, Hsp70 and Hsf1 in human HepG2 cells hypothermically treated at 4°C during different times as indicated, with one hour of recovery at 37°C. Relative optical density values of each protein compared to actin from Western blot analysis are expressed as mean ± SEM of three independent experiments. The statistical significance of the differences between values was assessed by (ANOVA) followed by Duncan multiple comparison post hoc test. A P<0.05 was considered to be statistically significant. The O.D. showed that Hsp60, Hsp70 and Hsf1 expression was increased in all treated samples, in contrast to control group. The higher increase of Hsp60, Hsp70 was observed in cells hypothermically treated during 1h. To Hsf1 the denser band belonged to the group treated during 2h. The lower levels of expression were observed in Hsc70, while Hsp70 and Hsf1 were the most induced. ANOVA showed no significative changes in Hsc70 between treatments. In the case of Hsp60, Hsp70 and Hsf1 the relative O.D. difference was significative to each treated sample compared to the control group (P<0.05).
DISCUSSION
This paper reports for the first time monomeric Hsf1 and Hsp60 induction under hypothermic stress in HepG2 cells, and corroborates the accumulation of monomeric Hsf1, as has been reported in mice (14). Moreover, Hsp60 has also been reported in rat brown adipose tissue under whole body hypothermia (13)
The increased expression of the heat shock proteins is possibly a natural protective and homeostatic response of the cell to the deleterious effects of the hypothermic stress. This response has been associated with the decrease in the rates of enzymatic processes, enhancement of free radical production and protein unfolding and misfolding promoted by hypothermia (15). The differential accumulation of Hsp60 and the inducible form of Hsp70 in all the stress conditions studied could be related to the assistant in the appropriate folding of denatured or newly synthesized peptides (16).
Likewise, it has been suggested that Hsps could be involved in tumorigenesis and to be essential in the survival of different cancer cells, possibly by the modulation of the activity of proteins involved in cell cycle and apoptosis (17, 18), as these proteins are frequently up-regulated in that kind of cells, as in hepatocellular carcinoma, (19). In this regard, Hsf1 itself is involved in overriding cell-cycle checkpoints and enhanced metastasis not dependent on Hsps (20, 21), as well downstream products of Hsf1, including Hsp70 and 27, are related to the increase of the invasive and/or metastatic capacity of tumors (22). Hsp90 has been involved in maintaining the quality of proteins in breast and prostate cancer (23). Cooperative promotion as a coordinated response of Hsp expression might represent one of the mechanisms involved in the protective effect of hypothermia widely used in medical strategies and preservation of biological samples (24), possibly due to thermotolerance.
Even though cold exposure without rewarming is related with decreased expression of several Hsps (25), the cytoprotective function of Hsp70 is associated with its anti-apoptotic effect at the rewarming, instead of during stress (26). Indeed, it is also understood that a general mechanism still unknown is involved in the regulation of Hsf1 activity during rewarming in response to cold stress in various species (11, 14). This mechanism could take place towards the activation of several transcription factors without its phosphorylation, while in hyperthermia occurs by the unique activation of Hsf1 preceded by its hyperphosphorylation (14). Additionally, in response to heat or cold stress an alternative splicing of the Hsf pre-mRNA has been identified (11). These observations suggest that the differences in Hsps expression in hypothermia or hyperthermia conditions are possibly related with the induction mechanisms.
The response to hypothermic stress has also been studied in HepG2 cells by Ohsaka et al. (26, 27). These have identified the induction of mitogen and stress activated protein kinases, which are closely related with the mechanisms of signaling implicated in cell repair and survival. It has been also suggested that the increasing of activity of antioxidants mechanisms such as the synthesis of catalase, superoxide dismutase and hemo-oxigenase 1 are involved in the response to hypothermia (28), together with the synthesis of other glycine-rich polypeptides of unknown function (27).
The possible explanation of the morphological changes has been related with the effect of the temperature in the molecules and changes in the rates of metabolic processes. The distribution of cell material is consistent with the changes in the cytoskeleton provoked by the hypothermia (3) revealed in the retraction of the material of the nucleus and nucleolar reorganization. The proteins of the nuclear matrix has been recognized as the most thermal labile proteins in the cell (29).
The aggregation of the material around the nucleus in hepatocytes exposed to hypothermia has been described in non tumoral hepatic cells (5). The retraction of material to the nucleus and the ring-shape nucleolar reorganization, and associated severe cell damage has been recognized in different pathologies in a variety of biological systems (30). In hypothermia, it has been also possible the recognition of nucleolar components segregation in the ring-shape structure. In depth this has been interpreted as a consequence of the disengagement between transcription and protein processing. Hsp70 is associated with transcription stabilization (31), its detection in the nucleolus could be related with those processes (30).
The production of oxygen radicals, ATP depletion and the increased Ca2+concentration are among the mechanisms associated with the hypothermia-induced morphological changes in the mitochondria after rewarming (32, 33). The swollen and roundshaped appearance of the mitochondria showed here, has been also described in proximal tubular cells (33), together with the generalized cell swelling in glial cells (34).
Synergistic promotion as a coordinated response of Hsp expression might represent one of the mechanisms involved in the protective effect of hypothermia widely used in medical strategies and preservation of biological samples. Cold stress could be an alternative to thermal stress, capable of inducting Hsp expression, by an unknown mechanism that needs to be determined.
ACKNOWLEDGEMENTS
This work has been supported by the Consejo de Desarrollo Científico y Humanístico (CDCH) from the Universidad Central de Venezuela and Laboratorios Elmor S.A. We thank Dr. Antonio De Maio who kindly provided HepG2 cell line, Dr. Tomás Hermoso and Lic. María Narváez for the anti-Hsf1 antibody and Mr. Raúl Colina for his excellent technical assistance.
REFERENCES
1. Sonna L, Fujita J, Gaffin S, Lilly C. Effects of heat and cold stress on mammalian gene expression. J Appl Physiol 2002; 92:1725-1742. [ Links ]
2. Lepock J. How do cells respond to their thermal environment?. Int J Hyperthermia 2005; 21: 681-687. [ Links ]
3. Welch W. Mammalian stress response: cell physiology, structure/function of stress proteins, and implications for medicine and disease. Physiol Rev 1992; 72:1063-1081. [ Links ]
4. Holland D, Roberts S, Wood E, Cunliffe W. Cold shock induces the synthesis of stress proteins in human keratinocytes. J Inv Dermatol 1993; 101:196-199. [ Links ]
5. Griffiths N, Evans P. Ultraestructural changes in hypothermically preserved hepatocytes. Cryobiology 2000; 40:176-181. [ Links ]
6. Stefanovich P, Ezzel M, Sheehan S, Tompkins R, Yarmush M, Toner M. Effects of hypothermia on the function, membrane integrity and cytoeskeletal structure of hepatocytes. Cryobiology 1995; 32:389-403. [ Links ]
7. Zeng X, Bhasin S, Wu X, Lee J, Maffi S, Nichols C, Lee K, Taylor J, Greene L, Eisenberg E. Hsp70 dynamics in vivo: effect of heat shock and protein aggregation. J Cell Sci 2004; 117: 4991-5000. [ Links ]
8. Baler R, Dahl G, Voellmy R. Activation of human heat shock genes is accompanied by oligomerization, modification and rapid translocation of heat shock transcription factor HSF1. Mol Cell Biol 1993; 13: 2486-2496. [ Links ]
9. Sarge K, Murphy S, Morimoto R. Activation of heat shock gene transcription by heat shock factor 1 involves oligomerization, acquisition of DNA-binding activity, and nuclear localization and can occur in the absence of stress. Mol Cell Biol 1993; 13:1392-1407. [ Links ]
10. Ruell P, Thompson M, Hoffman K. Heat shock proteins as an aid in the treatment and diagnosis of heat stroke. J Thermal Biol 2009; 34:1-7. [ Links ]
11. Fujikake N, Nagai Y, Popiel H, Kano H, Yamaguchi M, Toda T. Alternative splicing regulates the transcriptional activity of Drosophila heat shock transcription factor in response to heat/cold stress. FEBS Lett 2005; 579:3842-3848. [ Links ]
12. Rada A, Tonino P, Anselmi G, Strauss M. Is hypothermia a stress condition in HepG2 cells? Expression and localization of Hsp70 in human hepatoma cell line. Tissue Cell 2005; 37: 59-65. [ Links ]
13. Matz J, Blake M, Tatelman H, Lavoi K, Holbrook N. Characterization and regulation of cold-induced heat shock protein expression in mouse brown adipose tissue. Am J Physiol 1995; 269:R38-R47. [ Links ]
14. Cullen K, Sarge N. Characterization of hypothermia-induced cellular stress response in mouse tissues. J Biol Chem 1997; 272: 1742-1746. [ Links ]
15. Mestril R. Stress protein involvement in cardioprotection induced by hypothermia. J Mol Cell Cardiol 2001; 33:2075-2078. [ Links ]
16. Young J, Agashe V, Siegers K, Hartl F. Pathways of chaperone-mediated protein folding in the cytosol. Nat Rev Mol Cell Biol 2004; 5:781-791. [ Links ]
17. Garrido C, Gurbuxani S, Ravagnan L, Kroemer G. Heat shock proteins: endogenous modulators of apoptotic cell death. Biochem Biophys Res Commun 2001; 286:433-442. [ Links ]
18. Calderwood SK, Khaleque A, Sawyer D, Ciocca Daniel R. Heat shock proteins in cancer: chaperones of tumorigenesis. Trends Biochem Sci 2006; 31:164-172. [ Links ]
19. Lim S, Park S, Yoo J, Park Y, Kim H, Jang K, Cho J, Yoo B, Jung G, Park C. Expression of heat shock proteins (HSP27, HSP60, HSP70, HSP90, GRP78, GRP94) in hepatitis B virus-related hepatocellular carcinomas and dysplastic nodules. World J Gastroenterol 2005; 11:2072-2079. [ Links ]
20. Hoang AT, Huang J, Rudra-Ganguly N, Zheng J, Powell WC, Rabindran SK, Wu C, Roy-Burma P. A novel association between the human heat shock transcription factor 1 (HSF1) and prostate adenocarcinoma. Am J Pathol 2000; 156:857-864. [ Links ]
21. Wang Y, Theriault J, He H, Gong J, Calderwood S. Expression of a dominant negative heat shock factor-1 construct inhibits aneuploidy in prostate carcinoma cells. J Biol Chem 2004; 279: 32651-32659. [ Links ]
22. Ciocca D, Calderwood S. Heat shock proteins in cancer: diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperones 2005; 10:86-103. [ Links ]
23. Isaacs JS, Xu W, Neckers L. Heat shock protein 90 as a molecular target for cancer therapeutics. Cancer Cell 2003; 3:213-217. [ Links ]
24. Nagao S, Irie K, Kawai N, Nakamura T, Kunishio K, Matsumoto Y. The use of mild hypothermia for patients with severe vasospasm: a preliminary report. J Clin Neurosci 2003; 10: 208-212.
25. Sonna L, Kuhlmeier M, Carter H, Hasday J, Lilly C, Fairchild K. Effect of moderate hypothermia on gene expression by THP-1 cells: a DNA microarray study. Physiol Genomics 2006; 16:91-98. [ Links ]
26. Kao Y, Goto S, Jawan B, Nakano T, Hsu L, Lin Y, Pan M, Lai C, Sun Ch, Cheng Y, Tai M, Huang H, Chen Ch. Heat preconditioning ameliorates hepatocyte viability after cold preservation and rewarming, and modulates its immunoactivity. Transplant Immunol 2008; 18: 220-231. [ Links ]
27. Ohsaka Y, Ohgiya S, Hoshino T, Ishizaki K. Mitochondrial genome-encoded ATPase subunit 6+8 mRNA increases in human hepatoblastoma cells in response to nonfatal cold stress. Cryobiology 2000; 40: 92-10. [ Links ]
28. Matsumoto K, Honda K, Kobayashi N. Protective effect of heat preconditioning of rat liver graft resulting in improved transplant survival. Transplantation 2001; 71:862-868. [ Links ]
29. Roti Roti J, Kampinga H, Malyapa R, Wright W, Van der Waal R, Xu M. Nuclear matrix as a target for hyperthermic killing of cancer cells. Cell Stress Chaperones. 1998; 3:245-255. [ Links ]
30. Morcillo G, Gorab E, Tanguay R, Diez J. Specific intranucleolar distribution of Hsp70 during heat shock in polytene cells. Exp Cell Res 1997; 1:361-370. [ Links ]
31. Beck S, De Maio A. Stabilization of protein synthesis in thermotolerant cells during heat shock. Association of heat shock protein -72 with ribosomal subunits of polysomes. J Biol Chem 1994; 269:21803-21811. [ Links ]
32. Brinkkoetter P, Song H, Lösel R, Schnetzke U, Gottmann U, Feng Y, Hanusch C, Beck G, Schnuelle P, Wehling M, van der Woude F, Yard B. Hypothermic injury: the mitochondrial calcium, ATP and ROS love-hate triangle out of balance. Cell Physiol Biochem 2008; 22:195-204. [ Links ]
33. Bartels-Stringer M, Kramers C, Wetzels J, Russel F, Groot H, Rauen U. Hypothermia causes a marked injury to rat proximal tubular cells that is aggravated by all currently used preservation solutions. Cryobiology 2003; 47:82-91. [ Links ]
34. Healy D, Daly P, Docherty N, Murphy M, Fitzpatrick J, Watson R. Heat shock-induced protection of renal proximal tubular epithelial cells from cold storage and rewarming injury. J Am Soc Nephrol 2006; 17:805-812. [ Links ]

