










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkZootecnia Tropical
versión impresa ISSN 0798-7269
Zootecnia Trop. v.24 n.2 Maracay jun. 2006
Índice de condición de los mejillones Perna perna y Perna viridis (Bivalvia:Mytilidae) bajo un sistema suspendido de cultivo en la Ensenada de Turpialito, Golfo de Cariaco, Venezuela
Vanessa Acosta1*, Antulio Prieto1 y Cesar Lodeiros2
1Departamento de Biología, Escuela de Ciencias. Universidad de Oriente. Cumana, estado Sucre. Venezuela. *Correo electrónico: vanessaacosta@yahoo.com
2Departamento de Biología Pesquera. Instituto Oceanográfico de Venezuela. Cumaná, estado Sucre. Venezuela.
RESUMEN
Con la finalidad de conocer la talla mínima de comercialización en fresco, se comparó el índice de condición de los mejillones Perna perna y Perna viridis colocados bajo sistema de cultivo suspendido en la Ensenada de Turpialito, Golfo de Cariaco (Julio 2002 a 2003). Mensualmente, se determinó la dimensión de la concha en función de la longitud máxima de su eje dorso-ventral, las masas secas del músculo, gónadas y el índice de condición en ambas especies. Al final del estudio, Perna perna presentó el mayor promedio de talla (95,92 mm) con respecto a P. viridis (73,61 mm). La variación en peso de los diferentes tejidos secos se relacionó con el índice de condición de ambas especies. P perna alcanzó su más alto índice (39,89%) en diciembre, mientras que P. viridis lo presentó en enero (20,62%), coincidiendo en ambos con el incremento de los tejidos reproductivos y el aumento de la clorofila a durante el período de surgencia. Sin embargo, la mayor variación del índice de condición fue observada en P. viridis, debido posiblemente a una mayor intensidad de los procesos de maduración y desove, lo cual se evidenció con el mas alto coeficiente de variación (0,19), indicando que la fluctuación anual en el proceso reproductivo es más acentuado, comportamiento típico de las especies tropicales, a pesar de generar menor tejido reproductivo por unidad de tiempo que P. perna en los períodos de desove.
Palabras clave: Índice de condición, Golfo de Cariaco, Perna perna, Perna viridis.
Condition index of the mussels Perna perna and Perna viridis (Bivalvia:Mytilidae) under a suspended cultivation system in the Turpialito Bay, Cariaco Gulf, Venezuela
Summary
With the purpose of knowing the minimum size of commercialization in fresh, it was compared the condition index of the mussels Perna perna and Perna viridis placed under a suspended cultivation system in Turpialito Bay, Gulf of Cariaco (July 2002 to 2003). Monthly, the dimension of the shell was determined as a function of the maximum ventral axis length, dry mass of the muscle, gonads, rest of soma, and the index of condition in both species. At the end of the study, P. perna presented the highest size growth (95.92 mm) compared to P. viridis (73.61 mm). The variation in weight of the dry mass was related to the condition index of both species. Perna perna showed the highest value (39.9%) in December, while P. viridis presented it in January (20.6%), coinciding in both species with an increment of the reproductive tissue and the increase of the chlorophyll a during the period of upwelling. However, the highest variability in the condition index was observed in P. viridis due to the higher intensity in the maturation processes and spawning, which was evidenced by the largest coefficient of variation (0.19). This indicates that the annual fluctuation in the reproductive process is more accentuated a typical behavior of tropical species, in spite of generating smaller reproductive tissues per unit of time than P. perna in the spawning periods.
Keywords: Condition index, Gulf of Cariaco, Perna perna, Perna viridis.
Recibido: 10/01/06 Aceptado: 21/04/06
INTRODUCCIÓN
Los cultivos marinos de algunas especies de mejillones de la familia Mytilidae han llegado a constituir una parte importante en la producción de alimento para el consumo humano en países como España, Francia, Holanda e Italia (Hickman 1992). Aunque en Venezuela esta actividad no ha alcanzado niveles considerables de desarrollo (Salaya et al., 1973), puede llegar nuevamente a convertirse en una industria próspera. La costa norte del estado Sucre constituye prácticamente la única zona mejillonera del país, de donde se extrae aproximadamente el 93% del mejillón que se consume a nivel nacional; gracias a la explotación de dos especies de mejillones, Perna perna (Linné 1758) y Perna viridis (Linné 1758). Estas dos especies forman extensas poblaciones que actualmente aportan el 2,1% de la producción pesquera anual (Salaya, 1999). Ambos mejillones constituyen especies potencialmente cultivables por su rápido crecimiento y alta capacidad reproductiva (Salaya et al., 1973; Sreenivasan et al., 1989; León et al., 1998). Se han reportado elevadas tasas de crecimiento para P. perna, con tallas de hasta 90 mm aproximadamente en un año, bajo sistema de cultivo suspendido (Vélez, 1970; Prieto et al., 1999), mientras que Perna viridis es de crecimiento variable y se caracteriza por ser un mejillón grande, donde la talla adulta promedio en la India y áreas adyacentes, se encuentra entre los 80 y 165 mm de largo y ocasionalmente puede alcanzar hasta 300 mm (Ragopal et al., 1998)
La determinación del tamaño comercial en moluscos es de gran importancia, tanto para la explotación racional y manejo sustentable de las poblaciones naturales, como para establecer los costos de producción en cultivo. En este sentido, el índice de condición o cantidad relativa de carne en los bivalvos, según Cruz y Villalobos (1993), ha sido utilizado para estimar el valor del organismo para su explotación y consumo, siendo por lo general proporcional a la talla. Además, permite conocer como responde internamente el organismo ante las variaciones ambientales. Este índice ha sido asociado intrínsicamente a la reproducción, ciclos de utilización y almacenamiento de energía y extrínsecamente esta influenciado por la disponibilidad de alimento y temperatura (Prieto et al., 1999; Arrieche et al., 2004), siendo los tejidos de buena calidad de aspecto cremoso, mientras que los tejidos translucidos, con alto contenido de agua, poseen baja calidad (Hikcman e Illingworth, 1980; Bayne y Newell, 1993). Por lo tanto, la calidad de los mejillones es juzgada por el sabor y apariencia de la carne y llega a nivel óptimo cuando el bivalvo alcanza la madurez sexual; por esta razón los mejillones son consumidos principalmente cuando están próximos a efectuar su reproducción.
Con la finalidad de determinar la talla mínima de comercialización en fresco y como un requerimiento básico para aplicar normas de conservación y/o para la explotación de éstos mejillones con alto potencial de cultivo, se compararon los índices de condición de Perna perna y Perna viridis, bajo sistema de cultivo suspendido en la Ensenada de Turpialito, Golfo de Cariaco.
MATERIALES Y MÉTODOS
Se sembraron 90 semillas de cada especie de mejillón Perna perna (35,49 ± 3,16 mm) y Perna viridis (34,28 ± 3,99 mm) de manera separada, en cuerdas de caucho de 1,20 m de longitud. Un total de 72 cuerdas (36 de cada especie) fueron suspendidas en un Long Line durante 1 año (julio de 2002 hasta julio 2003). El Long Line fue colocado perpendicularmente a unos 30 m de la costa, en la Ensenada de Turpialito (10º27´30¨ N, 64º 01´52¨ O), localizada en la costa nororiental de Venezuela (Golfo de Cariaco). Los mejillones fueron obtenidos en forma manual de un banco natural existente en Guayacán, Península de Araya (estado Sucre, Venezuela).
Con la finalidad de estimar el crecimiento, mensualmente se extrajeron de las cuerdas 30 ejemplares de cada especie, a las cuales previamente se les eliminó todos los epibiontes presentes en la concha. La dimensión de la concha se estableció en función a la longitud máxima (Lt) en su eje dorso-ventral, con un vernier digital marca Mytutoyo de 0,01 mm de apreciación, mientras que el peso de la masa visceral fresca y de las valvas se determinó con una balanza analítica marca Denver, modelo TR204 de 0,001 g de apreciación. El músculo (Pm), lóbulos gonadales (pg) y resto del soma (Ps) constituido por branquias, manto, pie y glándula digestiva, fueron disectados y luego deshidratados en una estufa a temperatura continua a temperatura continua de 60ºC durante 72 h y luego se obtuvieron los valores de masa seca con una balanza analítica de 0,001 g de apreciación.
A cada especie se le determinó el índice de condición (Pfc/Pv x 100), según Nascimento et al. (1980) y Cabrera et al. (1983), donde Pfc es el peso fresco de la carne y Pv es el peso valva. También se determino el coeficiente de variación (Zar, 1984). En este estudio, el aumento del peso seco de la gónada, se consideró como indicador de la gametogénesis, mientras una disminución se interpretó como ocurrencia de desove.
Parámetros ambientales
A fin de evaluar la influencia de los factores ambientales (temperatura y disponibilidad de alimento), sobre el índice de condición de los mejillones bajo sistema de cultivo, se tomaron muestras semanales de agua en la zona de cultivo, con una botella Niskin de 2 L por triplicado. Las muestras de agua fueron filtradas en envases de 2 L con una malla de 153 µm, para estimar la disponibilidad del alimento mediante la biomasa del fitoplancton (clorofila a). Para el análisis de clorofila a se empleó el método espectrofotométrico, mediante las técnicas descritas en Strickland y Parsons (1972). En el sitio de cultivo se instalo un termógrafo electrónico marca Sealog-Vemco, para registrar la temperatura del agua a intervalos de 30 min.
Para determinar diferencias en relación al índice de condición entre las especies de mejillón analizadas, se aplicó un análisis de varianza sencilla ANOVA, previa comprobación de homogeneidad de su varianza. El nivel de significancia empleado para la aplicación del ANOVA fue del 95% (Zar, 1984).
RESULTADOS
Crecimiento de la concha
El patrón de crecimiento de ambas especies medido a través de la longitud máxima (Lt) fue significativamente igual (F=2,82; P>0,05), durante los primeros cuatro meses (julio-noviembre 2002; Figura 1). A partir de diciembre se produjo una diferenciación del crecimiento (F=11,32; P<0,05). Posteriormente entre enero y marzo nuevamente ambas especies alcanzaron tasas de crecimiento similares (F=12,82; P>0,05). No obstante, a partir de abril P. perna incremento su tasa de crecimiento, hasta alcanzar al final del estudio una talla superior (95,30 ± 7,91 mm) a la obtenida por P. viridis (73,3 ± 6,99 mm).
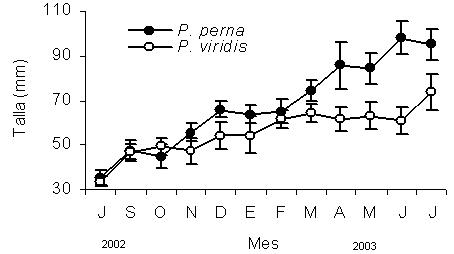
Figura 1. Longitud antero-posterior de los mejillones Perna perna y Perna viridis cultivados en la Ensenada de Turpialito, Golfo de Cariaco. Las líneas verticales en cada uno de los muestreos representan el error estándar con respecto a la media.
Músculo
En relación al peso seco del músculo, para el momento de la siembra, P. perna y P. viridis presentaron masas similares (Figura 2). No obstante, a partir de abril se comenzaron a observar diferencias significativas (F=23,01; P<0,05), en cuanto al crecimiento del músculo en ambas especies, siendo más pronunciado el crecimiento en P. perna (0,42 ± 0,23 g) al final del estudio, que en P. viridis (0,178 ± 0,18 g).
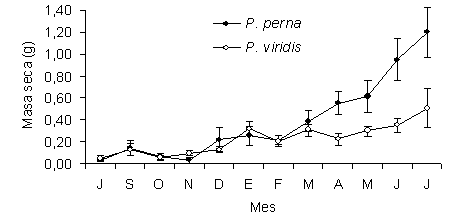
Figura 2. Masa seca del músculo de los mejillones P. perna y P. viridis cultivados en la Ensenada de Turpialito, Golfo de Cariaco. Las líneas verticales en cada uno de los muestreos representan el error estándar con respecto a la media.
Tejido gonadal
La masa seca de la gónada en ambas especies mostró desde el inicio de la siembra (julio) hasta octubre valores muy similares (0,02 ± 0,009 g P. perna y 0,018 ± 0,006 g P. viridis). A partir de noviembre en P. perna se produjo un incremento significativo de la masa gonadal (F=15,12; P<0,05), alcanzando en diciembre su máximo valor (0,82 ± 0,25 g), para posteriormente en enero producirse una caída brusca (0,313 ± 0,07 g), indicativo de un desove y el cual se extendió hasta febrero (0,160 ± 0,092 g). Posteriormente se produjo una leve recuperación, que se tradujo en aumento de los lóbulos gonadales, el cual fue progresivo, originándose un incremento de la masa de la gónada de aproximadamente 1,22 g, llegando a alcanzar 1,51 ± 0,52 g. Al finalizar el estudio Perna viridis presento un comportamiento muy diferente, ya que desde el inicio de la siembra hasta noviembre mostró una mínima producción de gónadas (0,018 ± 0,06 g a 0,025 ± 0,013 g). A partir de diciembre se produjo un ligero aumento de la masa gonadal hasta alcanzar en enero su mayor valor (0,239 ± 0,084 g), para posteriormente descender (desove) durante tres meses seguidos (febrero-abril), y luego se produjo una leve recuperación la cual se mantuvo constante hasta el final del estudio (Figura 3).
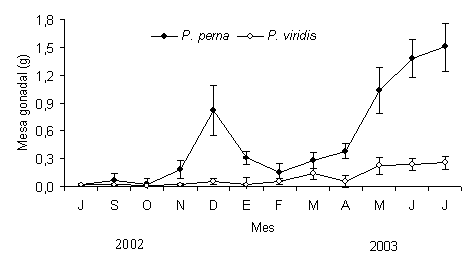
Figura 3. Variación mensual del tejido gonadal de los mejillones P. perna y P. viridis cultivados en la Ensenada de Turpialito, Golfo de Cariaco. Las líneas verticales en cada uno de los muestreos representan el error estándar con respecto a la media.
Índice de condición
En los cuatro primeros meses de cultivo (julio-octubre) P. perna y P. viridis mostraron el mismo índice de condición (Figura 4). Entre noviembre y diciembre se obtuvieron diferencias significativas entre ambas especies (F=15,13; P<0,05). Perna perna alcanzo su mayor índice en diciembre (39,98 ± 5,33%), el cual descendió bruscamente hasta febrero (16,68 ± 3,86%) y fue correlativo con el periodo de desove que se produjo para la misma época. A partir de marzo (21,82 ± 3,33%), se inicio un aumento significativo que se prolongó hasta el mes de mayo (31,99 ± 2,93%), el cual se mantuvo prácticamente constante hasta finalizado el estudio (33,08 ± 4,88%) y estuvo relacionado con el aumento de la masa gonadal que se produjo en los últimos cuatro meses.
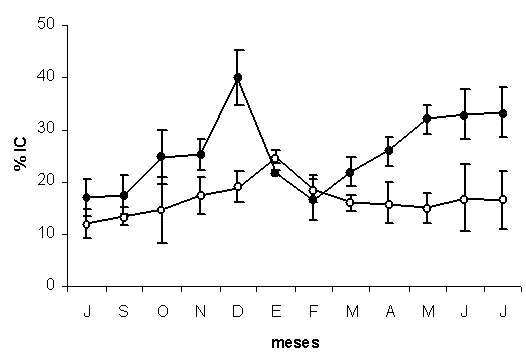
Figura 4. Variación del índice de condición (%) de los mejillones Perna perna y Perna viridis cultivados en la Ensenada de Turpialito Golfo de Cariaco. Las líneas verticales en cada uno de los muestreos representan el error estándar con respecto a la media.
En P. viridis, el índice de condición presento un leve incremento desde el inicio (11,98 ± 2,6%) de la siembra hasta alcanzar su máximo valor en enero (22,62 ± 1,20%), el cual coincidió con su mayor producción gonádica (Figura 2). Posteriormente se produjo un descenso que prácticamente se mantuvo constante hasta el final.
El coeficiente promedio de variación fue significativamente menor (P<0,05) en P. perna (0,16 ± 0,04), con respecto al obtenido en P. viridis (0,19 ± 0,071). El índice de condición en ambas especies fue correlativo a los periodos de madurez y desove de cada uno de los mejillones estudiados.
Parámetros ambientales
La temperatura superficial del agua mostró una fuerte variación con valores que alcanzaron 29,9°C en octubre 2002, para descender progresivamente hasta finales de enero 2003, cuando se registraron las temperaturas mas bajas (22-22,5°C; Figura 5A). Entre febrero y mediados de marzo 2003, la temperatura se mantuvo menor a 24°C, para luego incrementar a 26°C a finales de abril y mantenerse en este nivel hasta el final del estudio.
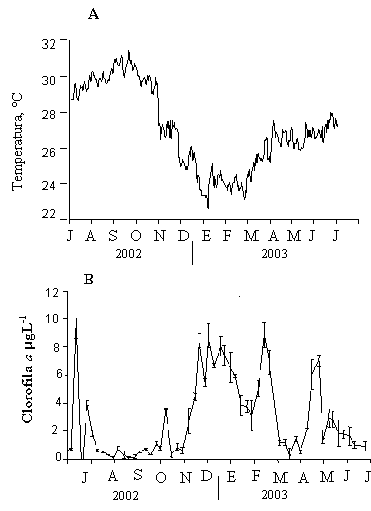
Figura 5. (A) Variación de la temperatura (ºC) y (B) Abundancia fitoplanctónica, estimada por la clorofila a (µg/L), durante el período de cultivo de los mejillones P. perna y P. viridis en la Ensenada de Turpialito, Golfo de Cariaco.
La biomasa fitoplanctónica, estimada por la concentración de clorofila a, mostró un patrón de variación inverso a la temperatura (Figura 5B). Este comportamiento es típico de la región, debido a los fenómenos de surgencia costera, establecidas por los regímenes de vientos en la zona (Lodeiros y Himmelman, 2000). Cuando las temperaturas fueron elevadas, la concentración de clorofila a estuvo por debajo de 1 µg L-1, desde mediados de agosto hasta finales de noviembre 2002, mientras que el resto del periodo estuvo caracterizado por concentraciones superiores a 4 µg L-1.
Discusión
En su máximo estadio de madurez, ambos mejillones presentaron un incremento en biomasa de tejido e índice de condición. En el caso del mejillón marrón Perna perna alcanzó su mayor índice en diciembre (cinco meses de cultivo), con una talla promedio de 65,84 ± 3,50 mm, la cual estuvo directamente influenciada por el peso de la gónada. Esto sugiriere que para esta época la especie presenta condiciones óptimas para la cosecha, lo que implicaría comercio y/o consumo. Estos resultados concuerdan con los reportados por Arrieche et al. (2004), quienes señalan que en los bancos naturales lo más recomendable es realizar la explotación de ejemplares de P. perna con una longitud superior a los 60 mm, que es cuando alcanza la mayor eficiencia de rendimiento de la carne, ya que en condiciones de cultivo, a los 5 meses se puede cosechar, lo que sugiere una rápida tasa de crecimiento de P. perna. No obstante, entre enero y febrero se observaron ligeras fluctuaciones en el peso de la masa seca del músculo, del tejido gonadal, lo que sugiere una liberación intensa de gametos (desove total), definiéndose hasta aquí el curso del ciclo reproductivo del mejillón. Por su parte, Perna perna invirtió una mayor cantidad de energía durante su ciclo reproductivo, demostrando un comportamiento característico de especie subtropical. Este comportamiento reproductivo estuvo relacionado con la mayor disponibilidad de alimento, principalmente biomasa fitoplanctónica tal y como se ha reportado en otros bivalvos, en donde factores ambientales como la disponibilidad de alimento y la temperatura influyen sobre los ciclos estacionales de engorde y de masa de tejido debido a la alta demanda energética que genera la gametogénesis (Nacimiento 1980; Lodeiros y Himmelman, 2000).
Perna viridis alcanzó su mayor índice de condición (20,62 ± 1,20%) en enero (a los seis meses de cultivo), con una talla de 55,70 ± 3,08 mm, mostrando un comportamiento reproductivo totalmente diferente al de P. perna, ya que la producción de gónadas de P. viridis se mantuvo prácticamente constante durante el estudio, con desoves parciales y continuos entre febrero, marzo y abril, presentando un comportamiento característico de una especie tropical. Sin embargo, su principal indicio de maduración se produjo en enero, coincidente con la mayor disponibilidad de alimento. Se ha reportado que en los trópicos los invertebrados marinos se caracterizan por presentar desoves prolongados la mayor parte del año, lo cual tiene una incidencia positiva sobre el índice de condición del organismo (Giese, 1959; Villalobos, 1980). No obstante, nuestros resultados demostraron un comportamiento diferente en el caso de P. viridis, ya que bajo cultivo suspendido, mostró un bajo índice de condición.
El peso de la gónada ejerció una marcada influencia sobre el índice de condición en P. perna y en menor proporción en P. viridis, de tal manera que la producción de gónadas es determinante en la estimación de la condición generalizada de los mejillones, ya que tiende a coincidir con la mayor biomasa de tejido (tejido húmedo, peso del músculo, peso de las gónadas, etc.). En este sentido muchos autores señalan que el comportamiento reproductivo de los bivalvos es un factor determinante en la estimación de la condición del organismo. Así, tenemos que los valores más bajos del índice de condición coinciden con la ocurrencia de gametogénesis y los valores más altos generalmente se combinan con la madurez (Villalejo-Fuerte y Ochoa-Báez. 1993). Este mismo comportamiento fue evidenciado en P. perna y en P. viridis bajo el sistema de cultivo suspendido.
El índice de condición alcanzado por P. perna en este estudio fue similar (16-39%) al reportado por Arrieche et al. (2004) en la misma especie (15-30%) y superior al de Perna canaliculus (8-15%) por Hickman e Illingworth (1980), mientras que el índice de P. viridis fue mucho más bajo que el reportado (11-20%) por Cheung (1991).
En ambos mejillones se observó que la masa seca total presentó cambios pronunciados en su peso antes y después de la liberación de gametos (desove), sugiriendo que existe una posible canalización de energía depositada en los tejidos somáticos hacia la gónada. Lo antes mencionado explica que los mejillones muestran una condición fisiológica dependiente de factores endógenos como la reproducción, con una notable interacción con los factores ambientales, principalmente la temperatura y la disponibilidad de alimento. Se ha reportado que, en condiciones naturales el alto índice de condición en P. perna está asociado con la biomasa fitoplanctonica, que es la que condiciona su ciclo reproductivo (Vélez, 1970; Arrieche et al., 2004). Este comportamiento es similar al observado en Anadara similis (Cruz, 1984), Mitella guyanensis (Sobaja, 1986), Chione subrugosa (Cruz, 1986) y Tagelus peruvianus (Rojas et al., 1988), los cuales presentan el desove masivo en los primeros meses del año, coincidente con mayor disponibilidad de alimento en el medio. Este comportamiento reproductivo podría estar relacionado con los afloramientos que se producen a principios de año en las aguas costeras que se explican en alguna medida por los valores altos en la productividad primaria señalados por Ferraz-Reyes (1987). El Golfo de Cariaco (zona de cultivo) se caracteriza por presentar aguas de bajas temperaturas y altos valores de nutrientes, oxígeno y biomasa fitoplantónica durante los primeros meses del año.
CONCLUSIONES
Nuestros resultados indican el hecho de que los factores ambientales tuvieron una marcada influencia sobre el crecimiento e índice de condición de los mejillones. Perna perna alcanzó su mayor índice de condición a los cinco meses de cultivo, con un rápido crecimiento y producción de biomasa, mientras que Perna viridis la producción de tejido fue muy baja y con lento crecimiento, lo que indica que las condiciones ambientales que acontecen en el Golfo de Cariaco no son favorables para el crecimiento de P. viridis.
AGRADECIMIENTOS
Se agradece al Consejo de Investigación de la Universidad de Oriente por la subvención de este estudio a través del Proyecto CI-5-1001-1155/03.
BIBLIOGRAFÍA
1. Arrieche D., B. Licet, N. García, C. Lodeiros y A. Prieto. 2004. Índice de condición, gonádico y de rendimiento del mejillón marrón Perna perna (Bivalvia:Mylilidae), del Morro de Guarapo, Venezuela. Interciencia, 27(11): 613-619 [ Links ]
2. Bayne B. y R. Newell. 1993. Physiological energetic of marine mollusks. En Sledding A.S. y K.M. Wilbur (Eds) The Mollusk. Academic Press, New York. Vol. 4, pp. 407-515. [ Links ]
3. Cabrera J., E. Zamora y O. Pacheco. 1983. Determinación del tamaño comercial de la ostra de manglar, Crassostrea rhizophorae (Guilding 1828) en sistema de cultivo suspendido en Estero Vizcaya, Limón, Costa Rica. Rev. Biol. Trop., 31: 257-261. [ Links ]
4. Carvajal J. 1969. Fluctuación mensual de las larvas y crecimiento del mejillón Perna perna y las condiciones ambientales de la Ensenada de Guatapanare, estado Sucre. Venezuela. Bol. Inst. Oceanogra. (Univ. Oriente), 8(1-2): 13-20. [ Links ]
5. Cruz R. 1984. Algunos aspectos reproductivos y variación mensual del índice de condición de Anadara similis (Pelecypoda: Arcidae) de Jicaral, Puntarenas, Costa Rica. Brenesia, 22: 95-105 [ Links ]
6. Cruz R. y C. Villalobos. 1993. Monthly changes in tissue weight and biochemical composition of the mussel Mytella guyanensis (Bivalvia:Mytilidae) in Costa Rica. Rev. Biol. Trop., 31: 175-179 [ Links ]
7. Cheung S. 1991. Energetic of transplanted populations of the green lipped mussel Perna viridis (Linnaeus) (Bivalvia:Mytilidae) in Hong Kong. II: Integrated energy budget. Ass. Mar. Biol., 8: 133-147. [ Links ]
8. Ferraz-Reyes E. 1987. Productividad primaria del Golfo de Cariaco, Venezuela. Bol. Inst. Oceanogra. (Univ. Oriente), 26(1-2):91-110. [ Links ]
9. Giese A. 1959. Comparative physiology: annual reproductive cycle of Marine invertebrates. Ann. Rev. Physiology, 21:547-576. [ Links ]
10. Hickman R. 1992. Mussel cultivation. En Gosling E. (Ed) The Mussel Mytilus: Ecology, Physiology, Genetics and Culture. Developments in Aquaculture and Fisheries Science, Vol. 25, Elsevier Science Publishers, NY. pp. 465-510 [ Links ]
11. Hickman R. y J. Illingworth. 1980. Condition cycle of the green lipped mussel Perna canaliculus in New Zealand. Mar. Biol., 60: 27-38. [ Links ]
12. León J., L. León y J. Millán. 1998. Cultivo suspendido del mejillón Perna perna L. 1758 (Mollusca:Bivalvia) en la isla de Cubagua, Venezuela. Caribb. Mar. Stud., 6: 12-18. [ Links ]
13. Lodeiros C. y H. Himmelman. 2000. Identification of factors affecting growth and survival of the tropical scallop Euvola (Pecten) ziczac in the Golfo de Cariaco, Venezuela. Aquaculture, 182: 91-114. [ Links ]
14. Nascimento I. y S. Pereira. 1980. Changes in the condition index for mangrove oyster (Crassostrea rhizophorae) from Todos os Santos Bay, Salvador, Brazil. Aquaculture, 20: 9-15. [ Links ]
15. Nakal A. 1980. Contribución a la ecología de la pepitona Arca zebra (Swainson 1883). Aspectos gametogénicos. Tesis de Grado, Licenciado en Biología. Universidad de Oriente. Cumaná. [ Links ]
16. Prieto A. 1980. Contribución a la ecología de Tivela mactroides (Born 1778). Aspectos reproductivos. Bol. Inst. Oceanogra. (Univ. Oriente), 29(2):323-328. [ Links ]
17. Prieto A., M. Vázquez y L. Ruiz. 1999. Energetic dynamics of growth in a mussel population Perna perna (Filibranchia:Mytilidae) in the northeast of Sucre state, Venezuela. Rev. Biol. Trop., 47(3): 399-410. [ Links ]
18. Ragopal S., V. Venugopalan, K, Nair, G. Van der Velde, H. Jenner y C. Hartog. 1998. Reproduction, growth rate and culture potential of the green mussel, Perna viridis (L) in Edaiyur backwaters, east coast of India. Aquaculture, 162 (3-4): 187-202. [ Links ]
19. Rojas J., C. Villalobos, F. Chartier y C. Villalobos. 1988. Tamaño, densidad y reproducción de la barba de hacha Tagelus peruvianus (Bivalvia: Solecurtidae) en el estero de Puntarenas, Costa Rica. Rev. Biol. Trop., 36: 479-483. [ Links ]
20. Salaya J., I. Beupertuy y J. Martínez. 1973. Estudio sobre la biología pesquera y cultivo del mejillón Perna perna (L.) en Venezuela. Proyecto MAC-PNUD-FAO. Informe Téc. Nº 62. Caracas. [ Links ]
21. Salaya J. 1999. La pesca y el cultivo de los moluscos bivalvos en Venezuela, situación y tendencias a nivel de Latinoamérica y el Caribe. Memorias Taller Venezolano sobre Aprovechamiento y Comercialización de Moluscos Bivalvos. Universidad de Oriente, Núcleo Nueva Esparta, Isla de Margarita, Venezuela. pp. 5-11. [ Links ]
22. Sibaja W.G. 1986. Madurez sexual en el mejillón Chora Mytella guyanensis, Lamarck, 1819 (Bivalvia:Mytilidae) del Manglar en Jicaral, Puntarenas, Costa Rica. Rev. Biol. Trop., 34: 151-156. [ Links ]
23. Sreenivasan P., R. Thangavelu y P. Poovannan. 1989. Biology of the green mussel Perna viridis (Linnaeus) cultured in Mutukadu lagoon. Madras. Indian J. Fish, 36: 149-155. [ Links ]
24. Strickland J. y T. Parsons. 1972. A practical handbook of seawater analysis. Fisheries Research Board of Canada. Bull. Nº 167. 2da ed. 310 p. [ Links ]
25. Tejera E., I. Oñate, M. Núñez y C. Lodeiros. 2000. Crecimiento inicial del mejillón marrón (Perna perna) y verde (Perna viridis) bajo condiciones de cultivo suspendido en el Golfo de Cariaco, Venezuela. Bol. Centro Invest. Biol., 34(2): 143-158. [ Links ]
26. Villalejo-Fuerte M. y R. Ochoa-Báez. 1993. El ciclo reproductivo de la almeja catarina Argopecten circularis (Sowerby, 1835), en relación con la temperatura y el fotoperiodo en Bahía Concepción B.C.S., México. Cienc. Mar., 19:181-202. [ Links ]
27. Vélez A. 1970. Fluctuación mensual del índice de engorde del mejillón Perna perna natural y cultivado. Bol. Inst. Oceanogr. (Univ. Oriente), 10(2): 3-8. [ Links ]
28. Vélez A. 1977. Ciclo anual de reproducción del ostión Crassostrea rhizophorae (Guilding) de Bahía de Mochima. Bol. Inst. Oceanogr. (Univ. Oriente), 16(1-2): 87-97. [ Links ]
29. Villalobos C. 1980. Variations in population structure in the genus Tetraclita (Crustacea:Cirrepedia) between temperate and tropical populations. IV. The age structure of T. stalactifera and conclunding remarks. Rev. Biol. Trop., 28: 353-359. [ Links ]
30. Zar J. 1984. Biostatistical Analysis. 2da ed. Prentice Hall, New Jersey. 120 pp. [ Links ]
