











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkBoletín de Malariología y Salud Ambiental
versión impresa ISSN 1690-4648
Bol Mal Salud Amb v.51 n.1 Maracay jun. 2011
Phlebotomine sand fly species co-occurrence at the local scale: Differences between agricultural and forested areas
Patrones de co-ocurrencia de especies de flebótomos a escala local: Diferencias entre áreas agrícolas y forestales
Luis Fernando Chaves*
Department of Environmental Studies, Emory University, Atlanta GA, 30322 USA.
Programa de Investigación en Enfermedades Tropicales, Escuela de Medicina Veterinaria, Universidad Nacional, Heredia, Costa Rica
*Autor de correspondencia: lfchave@emory.edu
SUMMARY
Phlebotomine sand flies (Diptera: Psychodidae: Phlebotominae) are vectors of Leishmania parasites, etiological agents of leishmaniases in humans and other mammals worldwide. A previous study showed that at large biogeographical scales sand fly patterns of co-occurrence are segregated, indicating that sand fly communities at such scales are likely shaped by environmental factors and inter-specific interactions. However, no study has addressed co-occurrence patterns at the local scale of disease transmission. Here, a null-model analysis of species co-occurrence patterns is presented, showing that at the local scale patterns of sand fly species co-occurrence shift from a segregated structure to random when comparing a forest to a farmland. These patterns may explain the ecological fitting of sand flies with regard to the choice of diurnal resting sites in transformed landscapes, which may impact the transmission of Leishmania parasites by disrupting patterns of blood foraging across vertebrate hosts.
Key Words: Phlebotominae, Leishmaniasis, C-score, null models, ecological fitting.
RESUMEN
Los flebotómos (Diptera: Psychodidae: Phlebotominae) son vectores de los parásitos Leishmania, agentes etiológicos de las leishmaniasis en el ser humano y otras especies de mamíferos a nivel mundial. Investigaciones previas demostraron que en escalas regionales las especies de angoletas se segregan, posiblemente por la influencia de factores ambientales e interacciones inter-específicas. Sin embargo, los patrones de co-ocurrencia no han sido estudiados a nivel local. Aquí se presenta un análisis de modelos nulos de patrones de co-ocurrencia que muestra que los patrones de co-ocurrencia de especies de flebotómos pasan de segregados a agregados cuando se compara un bosque con una granja. Tal resultado podría explicarse en el ajuste ecológico de los flebotómos con respecto a la selección de sitios de descanso diurno en paisajes transformados. Este ajuste ecológico podría impactar la transmisión de los parásitos de Leishmania al cambiar los patrones de alimentación sanguínea de los flebotómos con respecto a varias especies de hospedadores vertebrados.
Palabras clave: Phlebotominae, Leishmaniasis, C-score, modelos nulos, ajuste ecologico.
Recibido el 30/12/2010
Aceptado el 14/03/2011
INTRODUCTION
Sand flies are vectors of Leishmania spp., flagellates which cause leishmaniases in humans and other mammals worldwide. Leishmaniasis foci are often inhabited by several sand fly species (Jimenez et al., 2000; Rotureau et al., 2006; Salomon et al., 2008). Although several studies have made the point that in environments with higher vertebrate diversity the likelihood of sustaining high overall prevalence of Leishmania infections across all hosts is reduced (Alexander et al., 2001; Travi et al., 2002), little or no attention has been given to the community structure of sand fly vectors.
Patterns of species co-occurrence can be studied using null-model tests, a tool originally proposed to examine randomness in community assemblages (Gotelli, 2000). Nowadays, null-models are considered to be pattern-generating models based on randomisation of ecological data or random sampling from a known or imagined distribution (Gotelli, 2000). The results of studies using null models to study sand fly species co-occurrence patterns at large biogeographical scales (encompassing ecological life zones and altitudinal gradients) revealed these species to be more clustered than expected at random. This supports the hypothesis that large-scale sand fly species distribution is probably determined by the environment and inter-specific interactions (Chaves & Añez, 2004).
However, It is still unclear what patterns of sand fly species co-occurrence exist at the local scale; Do species tend to be clustered around some kind of habitat?, Are they associated with specific habitats that exclude other species, or Are these patterns completely unstructured/random?. This information can be very useful for targeting vector control measures, allowing specific habitats to be treated with insecticides or rendered unsuitable through environmental modification. Similarly, it is not known whether large scale landscape transformation affects sand fly co-occurrence patterns at the local scale. Patterns of species co-occurrence at the local scale could change with landscape transformation and impact the dynamics of Leishmania spp. parasite transmission, given differential access to reservoir and incidental hosts (Chaves et al., 2008). Also land use change can modify the composition of the vector community. Sand fly species with large "ecological fitting", i.e., species with the ability to use new or transformed habitats without the need for adaptation (Janzen, 1986), can dominate communities that used to be very diverse before land use change. It can be expected that undisturbed environments, like forests, when compared to transformed landscapes, like farmlands, would have communities that are more structured with regard to their specificity in use of resting sites, which are of great importance for leishmaniasis transmission risk, given the limited dispersal of these insects (Morrison et al., 1993). In the present study patterns of sand fly species co-occurrence are analyzed using null-model tests, looking at published data from sand flies captured at resting sites in two transmission foci with different degrees of human intervention, a farmland and a forest.
METHODS
Data
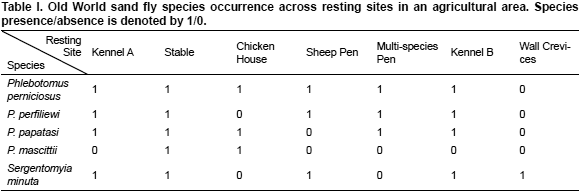
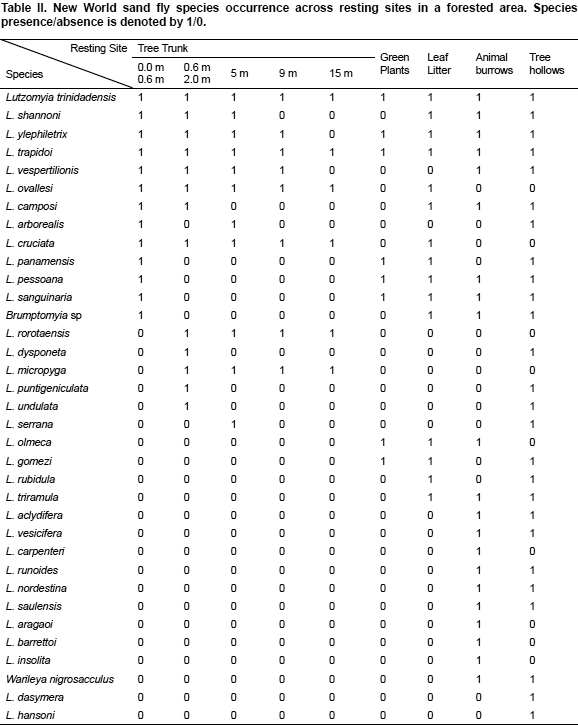
After the examination of the Web of Science and PubMed only two studies had detailed local data on sand fly species co-occurrence suitable for the analysis with null-model tests, one in a farmland, the other in a forest. Sand flies from a farmland focus were collected in Camerino, a hill town in the Marche region of Central Italy, using sticky traps and aspirator hand collection (Bongiorno et al., 2003). The insects were sampled bi-weekly over two four-month periods (June-September of 2000 and 2001) in a variety of sand fly diurnal resting sites, which included two kennels, a sheep pen, a stable, a chicken house, a multi-species pen (chicken, sheep, etc) and drainage openings within crevices of the walls of a human dwelling. Sand fly species collected from all resting sites in this study are summarized in Table I. Sand flies from the Forest focus were collected at Limbo Field Station which is located about 8 km Northwest of Gamboa, Panama using aspirator hand collection (Chaniotis et al., 1972). Samples were taken during a period slightly longer than two years (1969-1971), no information was reported on the sampling frequency. Sand fly species collected from all resting sites in this study are summarized in Table II.
Null Model Tests
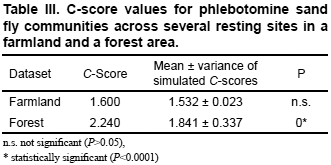
To test the null hypothesis that sand fly species co-occurrence at the local scale of transmission is random, C-scores (Stone & Roberts, 1990) were calculated using the data shown Table I and Table II.
Null-model tests for C-scores were carried out using the program Co-Occur (Ulrich, 2006). The null models were fixed–fixed, i.e., row and column sums of the original matrix were preserved. This algorithm was chosen because of its good statistical properties, including the low frequencies of type I and II errors when tested against random and structured matrices (Gotelli, 2000). The random matrices were created using a sequential swap algorithm (Gotelli & Entsminger, 2003). Null model tests were done using 5000 randomizations of the original dataset. If the C-score value of the dataset was significantly larger than the one obtained with the randomizations, species are considered to be non- aggregated. When a C-score is significantly smaller species are aggregated and when the two values are similar co-occurrence patterns are random (Stone & Roberts, 1992). Further details about null models and C-scores applied to the community ecology of vectors are presented in Chaves et al. (2010).
RESULTS
Results for the null model tests are shown in Table III. The C-score null-model test for the sand fly community structure in the farmland corresponded to that expected under random conditions, i.e., without differences between the expected value and the simulations. The C-score null-model test for the sand fly community structure in the forest was segregated, with the expected value being significantly higher than the value obtained by simulations.
DISCUSSION
Studies on sand fly ecology are very scarce when compared to studies on other vectors like mosquitoes or kissing bugs. Although it would have been ideal to study local patterns restricted to either New or Old World sand flies, this information has not been published for any given site with the high level of detail presented by Chaniotis et al. (1972) and Bongiorno et al. (2003). In addition, it can be argued that comparing New and Old World sand fly communities to study the use of resting sites is sound. For example, similar patterns of decreased diversity in agriculturally transformed landscapes have been reported across gradients of habitat intervention, as well as the widespread microhabitat usage by the most common sand fly species (Travi et al., 2002; Warburg et al., 1991). At the local scale of transmission, patterns changed from segregated in the forest to random in the farmland, a trend which probably is linked to the expansion of the disease to multiple hosts following environmental modification. This phenomenon has been partially studied in the Neotropics, where the extraction of selected tree species at a site in the Amazon forest did not change the rate of infection in Sand Flies following a logging event that led to the reduction of reservoirs (Pessoa et al., 2007). Although in the forest site both Lutzomyia trapidoi (Fairchild & Hertig) and Lu. trinidadensis (Newstead) were present across all microhabitats their densities were low in some of them (Chaniotis et al., 1972). Thus, the homogenization of the environment can favor the contact between most host species and sand fly species like Lu. trapidoi and Lu. trinidadensis. The latter can be explained by the "ecological fitting" (Janzen, 1985) of those sand fly vector species, i.e., their ability to be present everywhere and to exploit hosts that likely were associated with other sand fly species that have a diminished ability to contact several mammal host species. The fact that farmland sand fly species can rest across all suitable sites may indicate their ability to forage on blood from all hosts in the farmland, when compared to the forest where some species are strongly restricted to specific microhabitats. This finding precludes interventions based on the idea that the disease will disappear with the primary forest (Chaves et al., 2008), and favors measures directed at avoiding contact with vectors. However, more ecological studies on sand fly microhabitat and its relationship to blood foraging are needed to determine whether the choice of resting places is influenced by environmental conditions that may vary across other elements of the local landscape of transmission like climate.
ACKNOWLEDGEMENTS
The author is thankful to Dr. Bruce Alexander for his valuable comments on an earlier draft. This work was supported by Swedish International Development Agency – Netropica grant No. 04-N-2010 and the Gorgas Award from the American Society of Tropical Medicine and Hygiene.
REFERENCES
1. Alexander B., Agudelo L. A., Navarro F., Ruiz F., Molina J., Aguilera G. & Quiñones M. L. (2001). Phlebotomine sandflies and leishmaniasis risks in Colombian coffee plantations under two systems of cultivation. Med. Vet. Entomol. 15: 364-373. [ Links ]
2. Bongiorno G., Habluetzel A., Khoury C. & Maroli M. (2003). Host preferences of phlebotomine sand flies at a hypoendemic focus of canine leishmaniasis in central Italy. Acta Tropica. 88: 109-116. [ Links ]
3. Chaniotis B. N., Tesh R. B., Correa M. A. & Johnson K. M. (1972) Diurnal resting sites of phlebotomine Sandflies in a panamanian tropical forest. J. Med. Entomol. 9: 91-98. [ Links ]
4. Chaves L. F. & Añez N. (2004). Species co-occurrence and feeding behavior in sand fly transmission of American cutaneous leishmaniasis in western Venezuela. Acta Tropica. 92: 219-224. [ Links ]
5. Chaves L. F., Cohen J. M., Pascual M. & Wilson M. L. (2008). Social exclusion modifies climate and deforestation impacts on a vector-borne disease. PLoS Negl Trop Dis. 2: e176. [ Links ]
6. Chaves L. F., Harrington L. C., Keogh C. L., Nguyen A. M. & Kitron U. D. (2010). Blood feeding patterns of mosquitoes: random or structured? Frontiers in Zoology. 7: 3. [ Links ]
7. Gotelli N. J. (2000). Null model analysis of species co-occurrence patterns. Ecology. 81: 2606-2621. [ Links ]
8. Gotelli N. J. & Entsminger G. L. (2003). Swap algorithms in null model analysis. Ecology. 84: 532-535. [ Links ]
9. Janzen D. H. (1985) On ecological fitting. Oikos. 45: 308–310. [ Links ]
10. Jimenez A. E., Rojas J. C., Vargas F. & Herrero M. V. (2000). Temporal and spatial variation of phlebotomine (Diptera : Psychodidae) community diversity in a cutaneous leishmaniasis endemic area of Costa Rica. J. Med. Entomol. 37: 216-221. [ Links ]
11. Morrison A. C., Ferro C., Morales A., Tesh R. B. & Wilson M. L. (1993). Dispersal of the Sand Fly Lutzomyia longipalpis (Diptera, Psychodidae) at an Endemic Focus of Visceral Leishmaniasis in Colombia. J. Med. Entomol. 30: 427-435. [ Links ]
12. Pessoa F. A. C., Medeiros J. F. & Barrett T. V. (2007). Effects of timber harvest on phlebotomine sand flies (Diptera: Psychodidae) in a production forest: abundance of species on tree trunks and prevalence of trypanosomatids. Memorias Do Instituto Oswaldo Cruz. 102: 593-599. [ Links ]
13. Rotureau B., Gaborit A., Issaly J., Carinci R., Fouque F. & Carme B. (2006). Diversity and ecology of sand flies (Diptera: Psychodidae: Phlebotominae) in coastal French Guiana. Am. J. Trop. Med. Hyg. 75: 62-69. [ Links ]
14. Salomon O. D., Quintana M. G. & Zaidenberg M. (2008). Urban distribution of Phlebotominae in a cutaneous leishmaniasis focus, Argentina. Memorias Do Instituto Oswaldo Cruz. 103: 282-287. [ Links ]
15. Stone L. & Roberts A. (1990). The Checkerboard Score and Species Distributions. Oecologia. 85: 74-79. [ Links ]
16. Stone L. & Roberts A. (1992). Competitive-Exclusion, or Species Aggregation - an Aid in Deciding. Oecologia. 91: 419-424. [ Links ]
17. Travi B. L., Adler G. H., Lozano M., Cadena H. & Montoya-Lerma J. (2002). Impact of habitat degradation on Phlebotominae (Diptera: Psychodidae) of tropical dry forests in northern Colombia. J. Med. Entomol. 39: 451-456. [ Links ]
18. Ulrich W. (2006). CoOccurrence – a FORTRAN program for species co-occurrence analysis. www.uni.torun.pl/~ulrichw. [ Links ]
19. Warburg A., Montoya-Lerma J., Jaramillo C., Cruzruiz A. L. & Ostrovska K. (1991). Leishmaniasis Vector Potential of Lutzomyia Spp in Colombian Coffee Plantations. Med. Vet. Entomol. 5: 9-16. [ Links ]
