










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Facultad de Ciencias Veterinarias
versión impresa ISSN 0258-6576
Rev. Fac. Cienc. Vet. v.48 n.2 Maracay dic. 2007
Factores que Afectan la Producción y Composición de la Leche de Cabra: Revisión Bibliográfica.
Factors that Affect Yield and Composition of Goat Milk: A Bibliographic Review.
Alejandro Salvador*1, y Gonzalo Martínez**
*Departamento de Producción e Industria Animal, Facultad de Ciencias Veterinarias, Universidad Central de Venezuela, Apartado 4563. Maracay 2101. Estado Aragua, Venezuela.
** Instituto de Producción Animal, Facultad de Agronomía, Universidad Central de Venezuela. Correo-E: salex_77@cantv.net
Resumen
Se realizó una revisión bibliográfica sobre los diversos factores que afectan la producción y composición de la leche de cabra, estructurándose el trabajo en factores intrínsecos o propios del animal y factores extrínsecos. Los factores intrínsecos se clasificaron en genéticos, tomando en consideración la diferencia en producción y composición de la leche entre grupos o razas y los diferentes polimorfismos que afectan las fracciones proteicas de la leche; y factores no genéticos tales como edad o número de partos, estado de la lactancia, tipo de parto, duración del periodo seco previo, talla corporal, y características reproductivas como el ciclo estral y la preñez. Entre los factores extrínsecos se encuentran el efecto de la época y año de parto, las prácticas de ordeño tomando en consideración el ordeño mecánico, la duración del intervalo entre ordeños, duración del ordeño, frecuencia de ordeño y ordeño a fondo; el efecto del ejercicio, los cambios en la composición por efecto del procesamiento de la leche para la conservación o producción de queso, el efecto de la salud de la ubre y por último, el efecto de la nutrición y la alimentación. Se constató la necesidad de investigar más estos factores a nivel tropical, utilizando grupos genéticos mestizos que conforman la mayoría de los rebaños en estas zonas.
(Palabras clave: Cabra, producción lechera, composición de la leche, genética, edad, estaciones del año, reproducción)
Abstract
A review of the literature on several factors that affect the production and composition of goat milk was done. The search was based on both intrinsic and extrinsic factors. The intrinsic factors involved were classified as: genetic, taking into consideration the difference in milk production and composition among breeds and groups as well as the different polymorphisms that affect milks protein fractions; and non-genetic factors such as: age, number of parturitions, lactation stage, type of kidding, duration of previous dry period, body size and reproductive status, such as estrous cycle and pregnancy. The extrinsic factors examined were: effect and year of parturition, milking practices, with special emphasis on mechanical milking, milking duration, milking frequency, interval between milking duration, thorough milking; effect of exercise, changes in milk composition due to its processing for the conservation or production of cheese, effect of udder health and, finally, effect of nutrition and feeding. The results of the review show the need for further research of the above mentioned factors in tropical conditions using genetic crossbred groups which comprise most of the herd of these regions.
(Key words: Goats, milk production, milk composition, genetics, age, seasons, reproduction)
Recibido: 09/04/07 - Aprobado: 06/12/07
Introducción
Se estima que la población caprina en América es de 39 millones de cabezas, de las cuales el 57% está localizado en América Latina. Venezuela tiene una población estimada de 1.320.000 caprinos (FAO, 2005), constituyéndose en el cuarto país de América Latina en población de estos animales. Esto probablemente se deba a que los sistemas de producción caprina se adaptan bien a las zonas de vida áridas y semi – áridas, por lo cual tienen un nicho ecológico natural abundante en Venezuela. En el país existe una superficie de 41.023 km2 de estos ecosistemas, lo cual representa el 4,75% del territorio nacional, enmarcado básicamente en los estados Lara, Falcón y Zulia, pero con la característica principal de que el 94% de las explotaciones son de tipo extensivo – tradicionales, con producciones de leche muy bajas básicamente para el autoconsumo (Blanchard, 2001).
Estos sistemas de producción se caracterizan por utilización de cabras principalmente del tipo Criollo, ausencia de prácticas racionales de manejo de los rebaños, con pastoreo en vegetación natural, muy baja productividad de los rebaños (producciones de carne en el orden de los 6 kg/canal, en cabritones de 5 a 6 meses de edad, y de 200 a 250 g de leche por día, en lactancias que no sobrepasan los 100 días, propios de la zona semiáridas (Blanchard, 2001). Sin embargo, aunque la mayoría de las explotaciones de caprinos en Venezuela son de tipo extensivo, esta especie tiene un gran potencial productivo y social en la población, ya que aparte de poder utilizar ecosistemas no útiles para otras especies domésticas, tiene la posibilidad de tener un mayor número de animales por unidad de área que otras especies, un corto intervalo generacional y una elevada prolificidad, además del valor agregado de los productos derivados (principalmente quesos) y que la leche de cabra es mucho más digerible para los pacientes que no toleran la leche de vaca por alergia a sus proteínas o el tamaño de sus glóbulos de grasa (Infante et al., 2003), todo lo cual hace que la explotación de esta especie sea altamente rentable y sus productos derivados sean muy apreciados y cotizados en nuestras condiciones.
La producción cuantitativa y cualitativa de leche de cabra depende de muchos factores. Estos factores pueden ser agrupados en intrínsecos del animal (tales como genéticos, raza, nivel de producción, estado de lactancia, estado fisiológico, etc.) y extrínsecos (como la estación, temperatura, prácticas de manejo, sistema de ordeño, alimentación, estado de salud, duración del periodo seco, etc.; Haenlein, 1996).
Esta revisión bibliográfica tiene como objetivo recopilar los factores y cómo éstos afectan la producción y composición de la leche de cabra, principalmente en condiciones tropicales, de manera de hacer una contribución al conocimiento de la explotación de esta especie.
Revisión Bibliográfica
1. Factores intrínsecos
1.1.Genéticos
1.1.1. Variación genética entre grupos o razas
En diversos reportes (Zeng et al., 1996; Pacheco et al., 1998) han encontrado diferencias estadísticamente significativas (P<0,05) en la producción de leche en diferentes razas de cabras. De las razas de cabras lecheras, la Saanen es conocida como la Holstein de las cabras en el ámbito mundial, ya que produce altas cantidades de leche con bajos niveles de grasa. En el otro extremo se encuentra la raza Nubian que podría compararse también en el ganado bovino con la Jersey, ya que produce menos leche pero con un alto contenido de grasa. Otras razas como la Toggenburg, La Mancha, Oberhasli y Alpina están en un nivel intermedio entre las dos razas mencionadas, en cuanto a producción y contenido graso de la leche (Haenlein, 1996). En la Tabla 1 se observa la producción y la composición de la leche de algunas razas de cabras lecheras.
Tabla 1. Diferencias en la producción y la composición de la leche de diferentes razas de cabras lecheras
|
| Alpina | Saanen | Poitevine |
| N° observaciones | 156879 | 134672 | 639 |
| Días en lactancia | 273 | 278 | 248 |
| kg de leche | 754 | 774 | 475 |
| Proteína (g/kg) | 31,4 | 30,1 | 31,0 |
| Grasa (g/kg) | 35,8 | 33,3 | 34,6 |
Fuente: Hervé y Sigwald, (2001)
Sin embargo, los datos de producción de leche promedio de diferentes especies, han sido usados con cuidado, porque dentro de cada una de ellas hay grandes diferencias genéticas debido a las razas y a la selección de familias o líneas (Haenlein, 1996).
Muñoz (1997) obtuvo diferencias significativas (P<0,01) en la producción de leche total en diferentes grupos raciales en condiciones de clima semiárido en Venezuela.
La composición de la leche de cabra puede tener grandes diferencias dependiendo de la raza. Park et al. (1996) reportaron que el porcentaje de grasa en la leche está entre 2,3% y 6,9%, con un promedio de 3,3% y el porcentaje de proteína se ubica entre el 2,2% hasta 5,1%, con un promedio de 3,4%, existiendo una correlación negativa entre el rendimiento lechero y la composición, es decir, bajas producciones de leche tienen más alto contenido de sólidos (grasa y proteína) y viceversa.
La grasa es el constituyente más variable de la leche, mientras que la lactosa y los minerales son los menos variables. Sin embargo, las diferencias entre individuos dentro de una misma raza, son a menudo mayores que entre las diferentes razas. Esto, en gran parte, se debe al polimorfismo genético de las proteínas de la leche, el cual tiene gran importancia a nivel comercial debido a su efecto sobre el rendimiento quesero (Haenlein, 1996).
Fernández (2000) reportó una mayor producción y un mayor contenido de grasa y proteína en leche de cabras Alpinas en comparación con las mestizas de Alpina x Criolla.
Zeng et al. (1996) demostraron que las cabras Nubian producen leche con un contenido porcentual más alto de grasa, proteína y sólidos no grasos que las cabras Alpinas; al mismo tiempo, no encontraron diferencias significativas entre las razas en cuanto al recuento de células somáticas.
1.1.2. Polimorfismos
Dentro de las mismas razas se pueden identificar considerables diferencias en la composición de la leche. Parte de esta diferencias se debe al polimorfismo genético de las proteínas de la leche, el cual tiene un importante impacto comercial en la fabricación de queso ya que influye en su rentabilidad, rendimiento, sabor y potencial valor nutricional (Remeuf y Lenoir, 1986; Martin, 1993; Kalantzopoulos, 1994; Moioli et al., 1998). La mayoría de las proteínas lácteas presentan polimorfismo genético, siendo las más estudiadas la b-lactoglobulina, y las as1-, b- y k-caseína. Se ha encontrado que el polimorfismo de las proteínas lácteas afecta la firmeza y viscosidad del yogurt, el tiempo de coagulación de la leche por acción del cuajo, la estabilidad al calor, el pH, el contenido de caseínas, los sólidos totales y el fósforo. Se han identificado variaciones en la secuencia de aminoácidos de estas proteínas en caprinos, ovinos y bovinos, las cuales se corresponden con cambios en la secuencia de nucleótidos en el ADN de los genes que las codifican (Martin, 1993). Este polimorfismo de las proteínas es el responsable de cambios en la funcionalidad de las mismas: los cambios producidos en las características físico-químicas de las proteínas están relacionados con funciones específicas, tales como retención de agua, gelificación, emulsificación y formación de espuma (Moioli et al., 1998).
Los mayores contenidos de grasa y proteína, asociados con el polimorfismo genético, mejoran las propiedades de coagulación de la leche y su rendimiento en la producción de quesos y según Zullo et al. (2005) varios autores han demostrado que esas propiedades de coagulación están influenciadas, en leche de cabra específicamente, por el polimorfismo de la as1-caseína. En los últimos veinte años, diversos estudios han reportado la existencia de por lo menos siete (7) alelos asociados con cantidades variables de as1-caseína en la leche. Considerando la contribución aproximada de cada alelo al contenido de caseína total de la leche, los clasificaron en tres grupos: a) alelos A, B, C, con contribución Alta de 3,6 g caseína/L; b) alelo E, con contribución Media de 1,6 g caseína/L y; c) alelos F, D, O con contribución Baja o Nula de 0,6 g caseína/L (Park, 2007). En este sentido, los resultados de los trabajos de Jordana et al. (1996) demostraron que la distribución de los alelos en las razas caprinas ibéricas (Murciana-Granadina, Malagueña y Payoya) son bastante similares a las otras razas europeas, donde el alelo E es el predominante (rango entre 60 y 75%); por otro lado, las razas Canarias (insulares), y en particular el grupo étnico Canaria-Palmera, muestran un claro predominio de los alelos A y B. En la Tabla 2 se observa la frecuencia de alelos en diversas razas caprinas.
Tabla 2. Frecuencia media de alelos de polimorfismo de a-s-1-caseína en razas lecheras de cabras europeas
| Raza (número de muestras) | Tipos de polimorfismo | |||||
| Alto | Intermedio | bajo | ||||
| A | B | C | E | F | D+0 | |
| Murciana-Granadina (109) | 0,08 | 0,23 | - | 0,59 | 0,08 | 0,02 |
| Malagueña (373) | 0,09 | 0,09 | - | 0,65 | 0,04 | 0,13 |
| Payoya (111) | 0,05 | 0,19 | - | 0,76 | - | - |
| Canaria (74) | 0,28 | 0,32 | - | 0,20 | - | 0,20 |
| Palmera (22) | 0,68 | 0,23 | - | 0,09 | - | - |
| Majorera (21) | 0,07 | 0,38 | - | 0,24 | - | 0,31 |
| Tinerfeña (31) | 0,15 | 0,35 | - | 0,32 | - | 0,18 |
| Alpina Francés (213) | 0,14 | 0,05 | 0,01 | 0,34 | 0,41 | 0,05 |
| Alpina Italiana (80) | - | - | - | 0,35 | 0,59 | 0,06 |
| Saanen Francés (159) | 0,07 | 0,06 | - | 0,41 | 0,43 | 0,03 |
| Saanen Italiana (70) | 0,03 | 0,03 |
| 0,49 | 0,46 | - |
| Poitevine (209) | 0,05 | 0,35 | - | 0,45 | 0,14 | - |
| Corse (106) | 0,06 | 0,13 | - | 0,14 | 0,59 | 0,08 |
| Rove (147) | 0,12 | 0,05 | - | 0,62 | 0,10 | 0,11 |
| Garganica (54) | 0,61 | 0,37 | - | - | 0,02 | - |
| Maltesa (81) | 0,33 | 0,28 | - | 0,11 | 0,27 | 0,01 |
Fuente: Jordana et al. (1996)
La leche producida por cabras con los genotipos AA y BB en el locus de la as1 caseína presentaron los contenidos más elevados de grasa, proteína y sólidos totales. Este tipo de leche también presentó las mejores características de coagulación, con un tiempo más corto de la fase enzimática, mayor velocidad de coagulación y una mayor consistencia de la cuajada. Por el contrario, la leche proveniente de los genotipos EE y FF tenían una composición porcentual más baja, conjuntamente con una cuajada de menor consistencia (Zullo et al., 2005).
Los tipos de b-caseína de la leche de cabra también afectan las propiedades de fabricación del queso. Se han encontrado varios alelos de la b-caseína que se expresan en diversos niveles, incluso se ha reportado leche de cabra que carece totalmente de esta proteína, en la cual se presentó un tiempo de coagulación más prolongado y una cuajada más débil (Park et al., 2007).
1.2. Factores no genéticos
1.2.1. Edad y número de partos
Estos dos factores están muy relacionados. La mayoría de las cabras (dependiendo de la raza y del manejo) paren generalmente en el primer año de edad. Algunos autores mencionan que el volumen de leche aumenta hasta el cuarto o quinto año. Después de esto el volumen disminuye con el aumento de la edad del animal (Haenlein, 1996). La velocidad con la cual la producción disminuye es más baja que la velocidad con la cual aumentó, hasta el máximo rendimiento lácteo. La moda de vida productiva es alrededor de 12 años (Haenlein, 1996).
Milerski y Mare (2001) reportaron que la producción más alta por día ocurre entre los tres y cuatro años de edad en cabras; mientras que las que tenían un año de edad, eran significativamente inferiores en producción láctea. Los porcentajes de los componentes de la leche mostraron un incremento con el aumento en la edad sin diferencias significativas por clases. Al contrario, Večerová y Kríek en 1993 citados por Haenlein, (1996) encontraron aumento en la producción de leche hasta la décima lactancia, con disminución del porcentaje de grasa en la misma.
Diferentes autores (Zeng et al., 1995; Browning et al., 1995; Pacheco et al., 1998; Antunac et al., 1998; Fernández, 2000; Antunac et al., 2001) encontraron diferencias significativas en la producción y composición de la leche según el número de lactancia (número de parto). Todos ellos citan menor producción en la primera lactancia y acotan que en las primeras cuatro lactancias, la leche tiene más contenido de materia seca, sólidos no grasos, grasa y proteína en la leche con diferencias significativas (P<0,05). Sin embargo, Browning et al. (1995) encontraron que el rendimiento en grasa aumenta con el número de lactancia (P<0,01, 499 lactancias).
Oliver et al. (2001), encontraron una mayor producción de leche en cabras cuyo primer parto fuese entre los 15 y 24 meses de edad, pero no encontraron diferencias en la composición de la leche. Al contrario Iloeje et al. (1980); citado por Pinkerton 1987, determinaron diferencias en la producción y composición de leche de cabras con diferentes edades en las razas Saanen, Toggenburg y Nubian, como se observa en la Tabla 3.
Tabla 3. Efecto de la edad sobre la producción de leche (kg) y de grasa (kg) en algunas razas caprinas.
|
| Saanen | Toggenburg | Nubian | |||
| Edad (años) | Leche | Grasa | Leche | Grasa | Leche | Grasa |
| 1 | 672 | 23,1 | 672 | 22,7 | 552 | 23,6 |
| 2 | 829 | 29,5 | 811 | 26,3 | 641 | 28,6 |
| 3 | 876 | 31,3 | 835 | 29,5 | 694 | 31,8 |
| 4 | 839 | 29,9 | 879 | 28,6 | 733 | 32,7 |
| 5 y + | 679 | 24,5 | 743 | 24,0 | 639 | 28,6 |
| Nº de registros | 3307 | 4599 | 7456 | |||
Fuente: Iloeje et al. (1980) citado por Pinkerton, 1987
1.2.2. Estado de Lactancia
El estado de lactancia, independientemente de la especie o de la raza que se trate tiene una gran influencia en la composición de la leche. Muchos componentes de la leche ya sea de ovejas, cabras o vacas, especialmente grasa y proteína, son altos en calostro, al principio de la lactancia, mucho menor cuando llegan al pico de producción de leche y luego aumentan nuevamente a medida que baja la producción (Haenlein, 1996).
La producción diaria de leche aumenta firmemente durante las 4 semanas siguientes al parto y luego decrece gradualmente (Haenlein, 1996). Zeng et al. (1997) indicaron que este incremento se produce hasta los primeros 50 – 80 días después del parto. Durante este periodo de lactancia, hay una relación inversa entre los niveles de producción de leche y el contenido de grasa. En cabras Murciano-Granadinas tanto el porcentaje total de grasa como la composición en la leche varían. Al final de la lactancia, el contenido de grasa y proteína aumenta a medida que disminuye el rendimiento lácteo (Haenlein, 1996).
A medida que avanza la lactancia, aumenta el contenido porcentual de grasa, proteína, caseína, minerales, sólidos totales, sólidos no grasos, sodio, calcio, fósforo, magnesio y acidez titulable, mientras que el contenido de lactosa, potasio y citrato disminuyen significativamente como se observa en la Tabla 4 (Voutsinas et al., 1990).
Tabla 4. Efecto del estado de lactancia en la composición de la leche en 42 cabras Alpinas en Grecia
|
| Semana de lactancia | ||||
| 8-12 | 17-21 | 26-30 | 35-38 | 39-42 | |
| Grasa, % | 3,34 | 2,93 | 3,15 | 4,10 | 4,58 |
| Proteína, % | 2,79 | 2,95 | 3,32 | 3,91 | 4,25 |
| Caseína, % | 2,11 | 2,17 | 2,40 | 2,87 | 3,15 |
| Lactosa, % | 4,46 | 4,42 | 4,35 | 4,08 | 3,96 |
| Minerales, % | 0,72 | 0,78 | 0,80 | 0,82 | 0,84 |
| Total sólidos, % | 11,17 | 10,98 | 11,54 | 12,78 | 13,47 |
| Sólidos no grasos, % | 7,83 | 8,05 | 8,39 | 8,68 | 8,89 |
| pH | 6,58 | 6,61 | 6,57 | 6,54 | 6,52 |
| Acidez titulable | 16,60 | 16,30 | 16,65 | 17,38 | 17,44 |
| Densidad | 1,031 | 1,030 | 1,030 | 1,028 | 1,027 |
| Na, mg/100g | 50,20 | 52,20 | 54,00 | 54,75 | 55,50 |
| K, mg/100g | 170,40 | 166,00 | 155,20 | 146,75 | 144,50 |
| Na/K | 0,29 | 0,31 | 0,35 | 0,37 | 0,39 |
| Ca, mg/100g | 134,51 | 135,80 | 139,75 | 145,40 | 149,50 |
| P, mg/100g | 99,40 | 104,20 | 112,40 | 117,25 | 121,50 |
| Ca/P | 1,36 | 1,30 | 1,24 | 1,24 | 1,23 |
| Mg, mg/100g | 12,85 | 13,08 | 13,50 | 14,30 | 14,87 |
| Citrato, mg/100g | 145,42 | 114,50 | 99,64 | 88,23 | 81,17 |
Fuente: Voutsinas et al. (1990).
Antunac et al. (2001) encontraron durante las 5 primeras lactancias, un contenido más alto de materia seca, sólidos no grasos, lactosa, calcio y parcialmente fósforo al principio de la lactancia (primeros 50 días), en comparación con el período medio de la lactancia (100-150 días) siendo éstas significativas (P<0,01).
El contaje de células somáticas (CCS) en leche, aumenta a medida que progresa la lactancia (Zeng et al., 1997; Paapa et al., 1992). Zeng et al., 1997 encontraron altos CCS en leche durante las dos primeras semanas en lactancia y menores CCS en el segundo mes de lactancia. Sin embargo, la persistencia en la curva de lactancia en cabras es menor que en vacas y podría contribuir a elevar el CCS en estados avanzados de lactancia (Paapa et al. 1992). Variaciones diarias marcadas de CCS en cabras fueron observadas por Zeng et al. (1997).
Se han discutido muchas causas (edad, mes de lactación, etc.) de variación del CCS, en animales exentos de infección. Sin embargo, estas variaciones solamente son realmente importantes en leche calostral o cerca del secado, cuando las producciones son muy bajas. La Figura 1 refleja la variación producida en el CCS a lo largo de la lactación ovina, de un animal sin mastitis que normalmente presenta un aumento en el contaje de células somáticas tanto al principio como al final de la lactancia.
Figura 1. Curva de la lactancia y contaje de células somáticas en Ovino sin Mastitis.
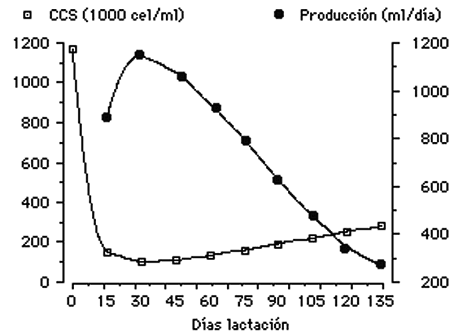
Fuente: Garcia – Bojalil (1992)
El estado de la lactancia también afecta el diámetro del coagulo de la grasa de la leche, el cual varía desde 1 a 10 m (en promedio 2 m) y su diámetro normalmente disminuye a medida que avanza la lactancia (Haenlein, 1996).
1.2.3. Tipo de parto
Varios autores (Gipson y Grossman, 1990; Večerová y Kríek, 1993 citados por Haenlein, (1996); Milerski y Mare, 2001) reportaron menor producción de leche en cabras con un solo cabrito por parto. Sánchez et al. (2006) describieron que cabras con partos gemelares produjeron más leche (1036,2±37,9 g/d) que las de parto sencillo (742,5±17,4 g/d). De acuerdo a esos autores, esta mayor producción de leche de las cabras con partos múltiples, sobre las cabras con partos simples, se debe al mayor estímulo que ejercen dos crías en lugar de una, sobre la succión de la ubre que conlleva a aumentar la producción de leche. Sin embargo, Fernández (2000) no encontró efecto del tamaño de la camada o número de cabritos por parto sobre la producción de leche.
Browning, et al. (1995) y Milerski y Mare (2001) mostraron que las cabras con partos simples tenían menor producción de leche que aquellas con partos dobles o triples (P<0,01). Milerski y Mare (2001) reportaron diferencias significativas en el porcentaje de grasa y proteína entre cabras con 1, 2, ó 3 y más crías, demostrando que las cabras con partos sencillos tenían menos leche pero más alto porcentaje de grasa y proteína. El rendimiento en grasa también es afectado por las características del parto (P<0,01; Browning et al., 1995).
1.2.4. Periodo seco
Un periodo sin producción de leche de 45-60 días entre lactancias, es requerido para obtener una óptima producción de leche en la próxima lactancia. Así mismo, períodos secos muy cortos o muy largos reducen la producción de leche en la subsiguiente lactancia.
Se necesita un promedio de 34 días de período seco, o de descanso productivo, para que el tejido glandular de la ubre sufra un proceso de involución y posterior regeneración de un nuevo tejido alveolar que garantice una próxima lactancia adecuada (Salvador, 1998).
Mayor número de días seca (>60 días) también determina reducción de la producción de la próxima lactancia, pues hay degeneración del sistema de conductos (García-Bojalil, 1992). En la Figura 2 se observa el efecto del número de días seca en la producción de la próxima lactancia, y se puede apreciar que las vacas que estuvieron secas entre 50 y 70 días, fueron las que presentaron las producciones más altas en la siguiente lactancia.
Figura 2. Efecto del número de días del periodo seco sobre la diferencia en la producción total de leche de vacas contemporáneas en la siguiente lactancia.
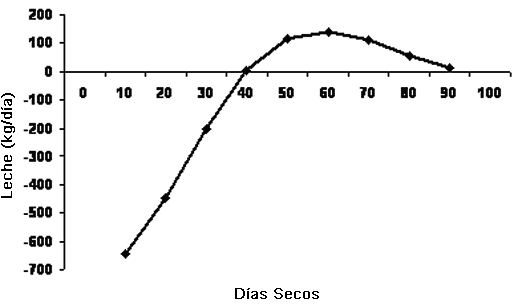
Fuente: Salama, (2005)
El calostro de las cabras con cero (0) días secas tuvo una menor concentración de IgG que el de las cabras con 27 y 56 días seca. En la lactación siguiente, las cabras con cero (0) días secas produjeron menos leche que las cabras con 27 y 56 días seca (1,73; 2,68; 2,53 L/ día, respectivamente), sin que hubiesen diferencias entre estas dos últimas. Los índices de apoptosis y proliferación del tejido mamario aumentaron en las cabras con 56 días secas durante la involución de la ubre (Salama, 2005).
1.2.5. Efecto de la talla corporal
La relación entre la talla corporal y la producción de leche ofrece resultados contradictorios. Las cabras grandes no son necesariamente más eficientes productoras de leche que las cabras pequeñas. Con base en la eficiencia energética, hay una pequeña diferencia en la producción de leche entre especies. Aunque la cabra produce más leche por unidad de peso corporal que una vaca, la eficiencia de utilización de la energía neta real es muy estrecha entre cabras y vacas. Sin embargo, las cabras de razas lecheras especializadas, tienen una tasa metabólica basal más alta, por lo cual tienden a tener una mayor eficiencia neta en producción láctea (Haenlein, 1996). En la Tabla 5 se observan algunas razas de cabras lecheras con sus tallas promedio y sus producción y composición de la leche. Se puede observar que no parece existir una relación directa entre talla y peso con la producción y el contenido de grasa y proteína de la leche.
Tabla 5. Dimensiones corporales, producción y composición de leche de razas de cabras lecheras, en 305 días de lactancia
| Raza | Altura (cm) | Peso (kg) | Producción de leche (kg) | Grasa (%) | Proteína (%) |
| Alpina | 76 | 61,2 | 903 | 3,56 | 3,06 |
| LaMancha | 71 | 59,0 | 777 | 3,80 | 3,29 |
| Nubian | 76 | 61,2 | 713 | 4,61 | 3,66 |
| Saanen | 76 | 61,2 | 942 | 3,52 | 3,02 |
| Toggenburg | 66 | 54,4 | 869 | 3,38 | 3,01 |
Fuente: Harris y Springer, (1996).
1.2.6. Ciclo estral y preñez
Aparentemente al acercarse el momento del estro, la cabra tiende a bajar la producción de leche. Esto es solo temporal y es normalmente compensado por un breve período de mayor producción después del estro. Puede haber un incremento en el porcentaje de producción de grasa durante el estro ya que baja la producción de leche y puede elevarse el CCS (Haenlein, 1996).
La preñez reduce la producción de leche durante la lactancia debido al aumento en los niveles de progesterona al final de la gestación. Salama (2005) en un ensayo con 30 cabras multíparas Murciano-Granadinas determinó el efecto negativo de la gestación sobre la producción y composición de la leche. Aunque no hubo diferencias significativas, el grupo de cabras con partos cada 24 meses, en lugar de cada 12 meses (normal), tuvo un 8% menos de leche en la producción total acumulada. Sin embargo, en la Figura 3, se observa cómo disminuye la producción de leche a partir de la semana 8 de gestación y significativamente a partir de la semana 10.
Figura 3. Comparación de las curvas de lactación de cabras lecheras Murciano – Granadinas sometidas a 1 parto al año o 1 parto cada dos años.
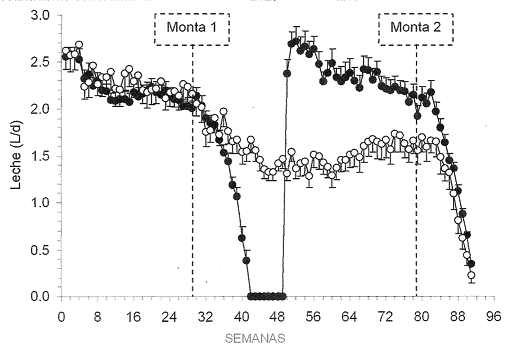
Circulos Blancos: un parto cada 2 años
Circulos Negros: un parto al año
Fuente: Salama 2005
Respecto a la composición de la leche, el mismo autor señala que las diferencias entre tratamientos solo se detectaron a partir de la semana 12 de gestación (semana 41 de lactancia) observándose que las cabras con un parto al año produjeron más grasa y proteína (4,33 vs 3,89% y 4,91 vs 3,38%, respectivamente) que las cabras con un parto cada dos años.
2. Factores extrínsecos
2.1.Efecto de la época de parto
La temperatura, la humedad, las prácticas de manejo y alimentación tienden a variar con la estación (lluviosa o seca), por lo cual se afecta la producción de leche y su contenido graso (Haenlein, 1996). Esto coincide con lo reportado por Gamarra (2006) en cabras Canarias en Venezuela, quien observó diferencias significativas tanto en la producción de leche como en la duración de la lactancia en la época de sequía y de lluvia, observándose que en la época de sequía se produce 11,54% más leche con una duración de la lactancia 16,14% menor que en la época lluviosa.
Asimismo, hay claras diferencias estacionales en la composición de la leche, con aumento o disminución de sus componentes (Renner, 1983), pero estos efectos se confunden con el efecto clima o los cambios de dieta. El clima frío (invierno) puede afectar los rendimientos en leche y la composición de la misma, y ambos están correlacionados negativamente. La alimentación en invierno provee normalmente proporciones y calidades diferentes de pasturas, ensilaje, heno o suplementos, los cuales influencian notablemente la composición de la leche.
De acuerdo con Le Jaouen (1986), en Francia, los contenidos de grasa y proteína en leche fluctúan con la época de parto o reportándose mayores niveles durante el otoño y los más bajos al principio del verano.
La temperatura ambiental, que a su vez está influenciada por la época de parto, tiene un efecto marcado sobre la producción y composición de la leche de cabra. Las cabras con altas demandas de producción son susceptibles de sufrir estrés calórico. Se observa entonces, depresión en el consumo de alimento (anorexia voluntaria) y reducción de la producción. Las temperaturas de la zona de confort para cabras europeas, en mantenimiento, están entre 25 a 30ºC, pero estos parámetros no han sido bien establecido para el crecimiento o para cabras en producción. El efecto del estrés calórico sobre el consumo, digestibilidad y tasa de pasaje en cabras no esta claro, sin embargo, balanceando raciones de acuerdo con la reducción de la producción, reduciendo la dieta de forraje, grano y grasa, suplementando con bicarbonato de sodio y otros minerales y maximizando el consumo de agua fría, puede ser beneficioso para el control del estrés calórico en cabras, además de la utilización de duchas, ventiladores y proveer sombra (Lu, 1989).
Las variaciones causadas por la cantidad de horas de luz del día (fotoperíodo) pueden afectar la secreción de leche. Las cabras que paren durante los días cortos tienden a secretar un tipo de leche con mayor contenido graso (Le Jaouen, 1986). Garcia-Hernández et al. (2007) observaron un efecto significativo del fotoperíodo sobre la producción de leche y el contenido de grasa, en cabras sometidas a períodos de luz de 20 horas al día respecto a las que tenían fotoperíodo natural de 13-14 horas de luz.
El mes de parto tiene efecto significativo (P<0,001; 651 lactancias) sobre todas las características de la leche: producción, duración de la lactancia, porcentaje y rendimiento de proteína y grasa (Pacheco et al., 1998).
En Francia, el resumen de los resultados nacionales del Dairy Control (Centre International Caprin, 2002) muestran mayores producción de leche en cabras que parieron entre octubre y noviembre que las que parieron entre marzo y abril. Estas variaciones superan en 1,4% y 1,1% para grasa y proteína, respectivamente, entre las dos épocas de parto.
Otros autores (Gipson y Grossman, 1990; Fernández et al., 2001) observaron una mayor producción de leche en cabras que parieron en otoño que aquellas que lo hicieron en primavera, debido probablemente a una mayor disponibilidad de pasto, en condiciones de clima subtropical.
De acuerdo a Oliver et al. (2001), aunque no encontraron diferencias significativas entre el rendimiento lechero en cabras que parieron en tres períodos diferentes (septiembre-diciembre, enero-abril, y mayo-agosto), si observaron que aquellas que parieron entre septiembre y diciembre tuvieron producción mayor. Estos resultados coinciden con los de Muñoz (1997), en Venezuela.
2.2. Año de parto
Browning et al. (1995), encontraron diferencias significativas por año de lactancia en el porcentaje de grasa y proteína en leche. Este efecto, a menudo, es considerado en modelos como un efecto complejo que incluye la estación. Por ejemplo, Milerski y Mare (2001) encontraron que el efecto año- estación de parto tenía una influencia significativa sobre la producción diaria de leche y en los contenidos de grasa y proteína (P<0,0001; 2762 cabras).
En un ensayo durante siete años en Venezuela, realizado por Muñoz (1997), el año de parto fue significativativo (P<0,01) sobre la producción de leche y los días en lactancia. El autor observó diferencias de 65,2 kg de leche entre los años de mayor y menor producción.
2.3. Prácticas de ordeño
Después del estímulo inicial para la bajada de la leche, se necesita un período de alrededor de 60 segundos para la respuesta de liberación de oxitocina, la cual tiene un efecto en vacas de alrededor de 5 a 8 minutos (Bruckmaier y Blum, 1998). En cabras lecheras, la liberación de oxitocina ocurre en pocos segundos, por lo que se pueden ordeñar inmediatamente después de la bajada sin tener que seguir estimulando. Esto ocurre debido a la incapacidad de las cabras, al contrario de las vacas, de cerrar completamente los conductos lácteos, por lo que no pueden retener completamente, el fluido de la leche (Haenlein, 1996) y a la disposición anatómica de la cisterna de la glándula y cisterna del pezón, que en cabras es mayor que en vacas, por lo que la porción láctea de la cisterna es muy superior al 20% de leche que tiene la vaca antes del ordeño y de la estimulación (Bruckmaier y Blum, 1998).
2.3.1. Ordeño mecánico
Lu et al. (1991) encontraron que niveles mayores de vacío incrementan el promedio y la tasa de eyección de la leche en el ordeño (P<0,001), disminuye el tiempo de ordeño (P<0,001), y eleva el CCS (P<0,05). Al respecto, Haenlein (1996) obtuvo resultados contrarios, ya que el ordeño a mano puede causar mayor incremento en el CCS que el ordeño mecánico.
En cabras ordeñadas a 52 kPa de presión de vacío, los resultados del California Mastitis Test (CMT) fueron menores cuando se incrementó la relación de pulsación de 50:50 a 70:30 y disminuyó el tiempo de ordeño (P <0,05). La más alta tasa de ordeño y los resultados más bajos de CMT y CCS, se obtuvieron en cabras ordeñadas con una relación de pulsación de 60:40. Incrementos en el ciclo de pulsación de 60 a 90 ciclos/min elevan el promedio de producción, la tasa máxima de secreción de leche, disminuye el tiempo de ordeño y los resultados de CMT. No se observó mejora en la tasa de secreción cuando el ciclo de pulsación aumentó a 120 ciclos/min. Los parámetros óptimos del equipo de ordeño para cabras son: nivel de vacío de 45 a 52 kPa, relación de pulsación de 60:40, y ciclo de pulsación de 90 ciclos/min (Lu et al., 1991).
2.3.2. Duración del intervalo de ordeño
Entre el ordeño matutino y vespertino, en el mismo día, la composición de la leche puede cambiar (Simos et al., 1991). Este aumento puede ser confundido con el nivel de rendimiento lechero. En la Tabla 6, se observa cómo aumenta el porcentaje de grasa, proteína y sólidos totales cuando disminuye el intervalo entre ordeños. Salama (2005) con cabras Murciano Granadinas, variando el intervalo entre ordeños entre 8 y 16 horas; reportó una velocidad de síntesis de 80 y 71 mL de leche/h, respectivamente. Al disminuir a un solo ordeño al día (24 h) se disminuye la velocidad de síntesis a 58 mL/h.
Tabla 6. Composición de la leche con diferentes intervalos entre ordeños
| Intervalo entre ordeños | ||
| 14 h | 10 h | |
| % grasa | 5,10 | 5,30 |
| % proteína | 3,54 | 3,58 |
| %sólidos totales | 13,94 | 14,30 |
Fuente: Haenlein, (1996).
2.3.3. Duración del ordeño
La primera leche extraída de la ubre contiene mucho menos grasa (tan bajo como 1 ó 2%) que la leche extraída al final del ordeño (tan alta como 7 a 9%). La razón para esta distribución de los glóbulos de grasa no es bien conocida. Los glóbulos se pueden agregar en el alvéolo, lo cual puede retrasar su pasaje hacia la cisterna de la glándula, al contrario que la porción más fluida que pasa más rápido. Inmediatamente del inicio del ordeño, la leche en los conductos galactóforos tiene menos grasa que en el alvéolo (Haenlein, 1996).
2.3.4. Frecuencia de ordeño
Según Peris et al. (1998), en la mayoría de los casos, la supresión de ordeños tiene influencia sobre el rendimiento lácteo y en la composición de la leche. En la Tabla 7 se observan las diferencias en producción de leche, grasa y proteína al variar el número de ordeños. Por ejemplo, cuando se pasa de dos ordeños a un ordeño al día, la producción disminuye entre 5 y 30%, dependiendo de la raza y momento de la supresión, mientras que el porcentaje de grasa aumenta y el de proteína no cambia.
Tabla 7. Efecto de la frecuencia de ordeños sobre la variación de la producción, composición de la leche y duración de la lactancia en cabras lecheras.
| N° de ordeños/día | Autor (año) | Leche Lts. [%] | Grasa kg [%] | Proteína kg [%] | Días en lactancia |
| 2x a 1x | Mocquot y Auran (1974) | –266 [45] | –7,1[43] | –8,4 [42] | –12 |
| Mocquot y Guillimin (1975) | –112 [41]1 | –3,2[41]1 | –3,4[39]1 | –16 | |
| –174 [38]2 | –4,9[37]2 | –4,6[34]2 | –16 | ||
| Mocquot (1978) | –196 [35] | –6,1[37] | –6,1[34] | –17 | |
| Capote et al. (1995) | –[5]1 | –[6]1 | –[5]1 | - | |
| Capote et al. (1999) | – [6.3]1 | – [9,97]1 | – [8,6]1 | - | |
| Salama et al. (2003) | - [18] | – [16] | – [2,43] | - | |
| 2x a 3x | Henderson y Parker (1987) | +[9] | - | - | - |
| Wilde et al. (1987) | +[20] | - | - | - | |
| Knight (1992) | +[10] | - | - | - |
X = número de ordeños. 1: cabras de primera lactancia, 2: tercera lactancia.
Fuente: Peris et al. (1998). Modificado.
Capote (1999) en un ensayo con cabras Canarias encontraron, que aunque al aumentar el número de ordeños de 1 a 2 al día incrementaba la producción alrededor de 6 - 8 % no se justificaba el doble ordeño por el aumento de trabajo e incremento en los costos de producción, por lo cual no se obtenía un mayor beneficio.
Salama et al. (2003) determinaron, en cabras Murciano-Granadinas (que se ordeñan una vez en lugar de dos veces al día) una disminución de la producción de 18% y un aumento del porcentaje de sólidos totales, grasa, y caseína. La disminución de la producción es más marcada en cabras de 1 a 4 lactancias, que en cabras con más lactancias, siendo el cambio más marcado, en la lactancia temprana (de 2 a 12 semanas).
Se ha demostrado que al aumentar la frecuencia de ordeño incrementa la migración de neutrófilos desde la sangre a la glándula mamaria, para una mayor eficiencia en la fagocitosis y defensa de la ubre contra infecciones (Paapa et al., 1992). Sin embargo, muchas cabras lecheras son ordeñadas una sola vez al día, rutinariamente.
2.3.5. Ordeño a fondo
Además del aumento en la frecuencia de ordeño, el ordeño a fondo (si se combina ordeño y mamado de los cabritos que remueven toda la leche residual) en la lactancia temprana, estimula el desarrollo de la glándula mamaria, incrementando tanto la proliferación como la diferenciación de las células alveolares, por lo cual se incrementa el rendimiento lácteo. Esta proliferación de células alveolares permanece las primeras semanas de lactancia (Peris et al., 1997).
La existencia de una proteína láctea de bajo peso molecular (6000 – 30000 Mr) denominada FIL (Feedback Inhibitor of Lactation, por sus siglas en inglés), en la leche de cabras es un factor de inhibición de la síntesis de leche. Al aumentar la cantidad de leche en la glándula aumenta el contenido del FIL, por lo cual disminuye la producción de leche. Al ordeñar a fondo, disminuye la cantidad de FIL, lo cual incrementa la producción (Salama et al., 2003).
Adicionalmente, la presión intramamaria que se produce al acumularse la leche en la glándula (cisterna, conductos y alvéolos) produce una disminución en la velocidad de síntesis de leche y disminución de la producción láctea, por lo que el ordeño a fondo disminuye la presión intramamaria y aumenta la producción de leche. Esto es muy importante en la lactancia temprana (primera mitad) durante la cual se produce mayor cantidad de leche; sin embargo, en la lactancia tardía (segunda mitad) este factor pierde importancia, pues la cisterna de la ubre parece contener bien la leche producida (Salama et al., 2003).
2.4. Salud de la ubre
Una forma rápida y eficiente de medir la salud de la ubre es la cantidad de células somáticas en leche. Sin embargo, la secreción de leche en cabras es apocrina, mientras que en vacas es merocrina (Haenlein, 1996), lo cual explica el porque la leche de cabra puede tener un alto CCS, especialmente al final de la lactancia o en las últimas porciones de leche al escurrir la ubre, sin que esto tenga alguna relación con mastitis (Haenlein, 1996). Algunos componentes de la leche como las proteínas del suero, lactosa, lipasa, sodio, y cloro, se incrementan cuando hay mastitis, mientras que la grasa, sólidos, caseína, calcio, fósforo, potasio y el rendimiento en queso, disminuyen (Haenlein, 1996). Resultados similares reportaron Zumbo et al. (2004) y Salvador et al. (2006), como se observa en la Tabla 8, al analizar las correlaciones entre los componentes y las características químicas de la leche de cabras mestizas Canarias en Venezuela. Aunque no hubo diferencias significativas (P>0,05) entre la producción de leche y sus propiedades que determinen la presencia de mastitis, estos autores reportaron una correlación negativa leve entre la producción de leche diaria y la acidez y el Wisconsin Mastitis Test de –0,17 y -0,20, respectivamente.
Tabla 8. Correlación entre los componentes y las características químicas de la leche de cabras mestizas Canarias
| Componentes (%) | Características | |||
| Crioscopia | Acidez | WMT | pH | |
| Grasa | - 0,33** | 0,41** | 0,27** | - 0,23** |
| Proteína | - 0,38** | 0,57** | 0,37** | - 0,11 |
| Caseína | - 0,31* | 0,55** | 0,39* | - 0,35** |
| Sólidos totales | - 0,34** | 0,48** | 0,38** | - 0,32** |
| Sólidos no grasos | - 0,16 | 0,29** | 0,28** | - 0,25* |
| Cloruros | - 0,04 | - 0,06 | 0,30** | - 0,05 |
| Humedad | 0,34** | - 0,48** | - 0,38** | 0,32** |
| Cenizas | - 0,25* | 0,24* | 0,28** | - 0,40** |
WMT: Wisconsin Mastitis Test. **: P<0,01, *: P<0,05
Fuente: Salvador et al. (2006)
2.5. Nutrición y alimentación
Sin tomar en cuenta la genética, existe una relación entre la cantidad y composición de la dieta diaria y los requerimientos para producción. Variaciones de la dieta pueden traer cambios importantes en la producción y composición de la leche (Moranh-Fehr, 2005; Haenlein, 1996). En términos generales, el consumo mínimo diario de materia seca es de 3% del peso vivo en la mayoría de las cabras, pero las altas productoras (por encima de 1 kg de leche/día) pueden llegar a consumos del 5% del peso vivo (Haenlein, 1996) e, incluso, pueden consumir hasta 7% de materia seca (MS) de su peso vivo, en comparación con el consumo de 3-4% MS de las vacas (Jimeno et al., 2003). Este nivel de consumo elevado muestra que la cabra lechera necesita abundantes nutrientes para la síntesis láctea. La eficiencia de producción de leche por cabras lecheras es bastante similar a la de las vacas (Harris y Springer, 1991; Jimeno et al., 2003). Las cabras son muy eficientes en la conversión alimenticia y además tienen una capacidad relativa más grande para el consumo de forraje que las vacas u ovejas (25-40% de peso vivo, en comparación al 12,5-15% para vacas, 12,5-20% para ovejas) (Botnick, 1994).
Sahlu et al. (2004) estudiaron el efecto de tres dietas de preparto para cabras Alpinas, con tres niveles de proteína bruta (8,5, 11,5 y 14,5% MS) y de EM (1,8, 2,16 y 2,53 Mcal/kg MS) sobre la variación del peso vivo (en preparto), la prolificidad y el rendimiento en producción de leche de la siguiente lactancia. La producción de leche aumentó linealmente como respuesta a la concentración energética de la dieta de preparto y cuadráticamente en respuesta al contenido proteico de la dieta (2,59; 3,26 y 3,07 kg/d para 8,5; 11,5 y 14,5% proteína bruta, respectivamente).
Con el propósito de convertir nutrientes en alta producción de leche, la densidad de las proteínas y la energía del consumo de alimento diario deben incrementarse, debido a la limitada capacidad del rumen en términos de volumen. Hay que mantener una buena calidad del forraje y una mezcla adecuada de granos para el mejor aprovechamiento en el mantenimiento de altos niveles de producción. En este aspecto, las cabras tienen ventaja sobre las vacas y ovejas ,ya que además de pastar son ramoneadoras (Hernández, 2002; Baldizán y Chacón, 2004; Preston, 2004) por lo cual pueden tener acceso a frutos, tallos y hojas de alto valor nutritivo, que aunado a sus hábitos de consumo altamente selectivos le garantizan una dieta nutricional rica. Rodríguez-Parra y Caja (2004) determinaron que las cabras Murciano Granadinas rechazaban el 30% de la ración ad libitum, mientras que ovejas Merino solo rechazaban el 20%.
Variaciones en la dieta o la composición de la misma también afectan la composición de la leche. Chilliard et al. (2003) hicieron una revisión de la literatura relacionada con el efecto de la suplementación de diferentes fuentes de grasa y en diferentes proporciones sobre la producción, contenido graso y proteína de la leche de cabras lecheras, la cual se observa en la Tabla 9. Estos autores demostraron la existencia de una alta correlación entre el balance energético y el porcentaje de ácidos grasos con más de 18 átomos de carbono en la leche.
Tabla 9. Cambios en la producción de leche, grasa y proteína con diferentes fuentes de grasa en cabras lecheras.
| Fuentes de lípidos (% del concentrado) | Producción de leche (kg/día) | Grasa (g/kg) | Proteína (g/kg) | Autor |
| Ácido Palmitico 14% | -0,06 | +11,4 |
| Astrup et al., 1985 |
| Ácido Estearico 14% | +0,10 | +6,5 |
| Astrup et al., 1985 |
| Grasa animal (5% de la ración) | +0,38 | +2,0 | -1,0 | Gelaye and Anoah, 1988 |
| Grasa animal (5% de la ración) | -0,29 | +7,0 | +1,0 | Lu, 1993 |
| Grasa animal 4% | -0,09 | +3,7 | +1,4 | Daccord, 1987 |
| Sales de Ca 15% (aceite de palma) | -0,02 | +14,2 | +1,5 | De Maria Ghionna et al., 1987 |
| Sales de Ca 3-4% ración (aceite de palma) | +0,10 | +3,7 | -0,1 | Teh et al., 1994 |
| Sales de Ca 3-4% ración (aceite de palma) | +0,21 | +5,2 | +0,3 | Rousselot et al., 1995 |
| Aceite de Canola 4% | +0,19 | +9,2 | -0,3 | Mir et al., 1999 |
| Linaza tratada 23% | -0,20 | +6,3 | +1,4 | Ferlay et al., |
| Linaza 14% | +0,27 | +5,5 | +2,5 | Rouel et al., |
| Aceite de Linaza 5% | +0,30 | +4,2 | +2,4 | Rouel et al., |
| Semilla de girasol 9% | +0,42 | +6,3 | +2,1 | Rouel et al., |
| Aceite de girasol 5% | +0,42 | +4,9 | +1,3 | Rouel et al., |
| Soya 26% | +0,49 | +3,9 | +1,4 | Rouel et al., |
| Soya 49% | -0,14 | +9,3 | +0,3 | Bernard et al., |
| Semilla de algodón 18% | -0,02 | +4,4 |
| Bertocci et al.,1988 |
| Aceite protector 7% | +0,08 | +2,9 | -0,8 | Lanzani et al.,1985 |
| Ensilaje de torta de oliva | -0,05 | +3,1 | -0,5 | Hadjipanayiotou, 1999 |
Fuente: Chilliard et al. (2003)
La adición de ácidos grasos de cadena larga incrementa el contenido de grasa de la leche y el rendimiento, sin cambio en el contenido de proteína de la leche. Los ácidos grasos C4 – C14 de la grasa de la leche disminuyeron drásticamente, mientras que los ácidos grasos de cadena larga aumentaron, incluyendo los ácidos insaturados y colesterol (Baldi et al., 1992).
Las variaciones en la cantidad y diferencia en las fuentes de proteína de la dieta pueden afectar la composición de la leche de cabra. Al respecto, Sanz Sampelayo et al. (1999) utilizando grano de habas, harina de girasol, gluten de maíz y semilla de algodón como diferentes fuentes de proteína de la dieta, observaron diferencias significativas (P<0,01) en la composición de la leche, en el porcentaje de sólidos totales (15,21; 15,52; 16,40; 16,33, respectivamente), porcentaje de proteína (3,25; 2,87; 3,50; 3,18, respectivamente), pero no obtuvieron diferencias en el porcentaje de grasa ni en la producción total de leche.
Las cabras son menos sensibles que las vacas a la longitud o tamaño de partícula procedente de los forrajes. El contenido en grasa de la leche no disminuye al reducir el tamaño de partícula del mismo, siempre que se mantenga un nivel mínimo de fibra en la dieta, ya que tanto la grasa total como la proteína total de la leche de cabra se encuentran más influenciadas por el consumo de energía que por el tipo de forraje (alfalfa henificada vs pellets) que se incorpore a la dieta (Sanz Sampelayo et al., 1999). En determinadas ocasiones y en dietas destinadas a cabras de alta producción para las cuales no se dispone de forrajes de buena calidad, una estrategia adecuada puede consistir en sustituir parte de ese forraje por subproductos de buena calidad y forrajes peletizados (granulados), en la que la relación fibra:concentrado podría estar próxima a 35:65 manteniendo los niveles de fibra dentro de las recomendaciones. Bava et al. (2001) demostraron que las cabras toleran bien dietas sin forrajes y altas en proteína, durante toda la lactación, sin ningún efecto negativo sobre la producción de leche, su composición o la salud del animal, siempre que el nivel de fibra en la dieta sea adecuado. Al contrario de estos resultados, Kawas et al. (1991) en Brasil, demostraron que sustituyendo en la ración, el forraje por alimento concentrado en una proporción de 45:55, se deprime el porcentaje de grasa en la leche, aumenta el contenido de proteína, el rendimiento lechero y la ganancia de peso, mientras decrece el tiempo de alimentación y de rumia.
Conclusiones y Recomendaciones
En esta revisión de literatura, se analizaron dos grandes grupos de factores que afectan la producción y composición de la leche de cabra: los factores intrínsecos como la raza, los polimorfismos de la fracción proteica de la leche, el número de partos, el tipo de parto, el estado de la lactancia, el período seco previo, el estro y la gestación; y los factores extrínsecos como la época y año del parto, las prácticas de ordeño (tipo, duración, intervalo, frecuencia de ordeños), la salud de la ubre y la alimentación, siendo esta última tan importante y variable que se necesitaría una revisión bibliografica aparte sólo para este tópico.
La mayoría de las investigaciones recientes, realizadas en los países desarrollados, apuntan hacia las diferencias que existen en la composición de la leche, específicamente hacia los diferentes polimorfismos de la fracción proteica y los factores que lo determinan, así como estas diferencias afectan el rendimiento quesero como principal factor de la rentabilidad de esta especie.
Sin embargo, se constató la necesidad de investigar más acerca de estos factores a nivel tropical, utilizando grupos genéticos mestizos, ya que la mayoría de los animales pertenecen a estos grupos genéticos en nuestras condiciones, y se pudo evidenciar la escasa bibliografía encontrada al respecto. La profundización de investigaciones en estos aspectos, aportaría conocimientos de esta especie con elevada capacidad productiva lechera para el desarrollo de las regiones con déficit en la producción de leche en condiciones tropicales.
Referencias
1. Antunac, N.; Samarija, D.; Havranek, J. L.; Pavic, V.; Mioc, B. 2001. Effects of stage and number of lactation on the chemical composition of goat milk. Czech. J. Anim. Sci., 46:1212-1819. [ Links ]
2. Aparicio, N.; Pérez, J. 2001. Comenzar un programa de control de la mamitis ovina y caprina. Dirección Electrónica: http://www.iespana.es/capra/index.htm. Consultado el 25/02/02. [ Links ]
3. Baldi, A.; Cheli, F.; Corino, C.; Dell`Orto, V.; Polirodi, F. 1992. Effects of feeding calcium salts of long chain fatty acids on milk yield, milk composition, and plasma parameters of lactating goats. Small Rumin. Res., 6:303-310. [ Links ]
4. Baldizán, A.; Chacón, E. 2004. Sistemas agroforestales con ovinos y caprinos. IV Congreso Nacional de Ovinos y Caprinos. UNEFM. Santa Ana de Coro. pp. 140-163. [ Links ]
5. Bava, L.; Rapetti, G.; Crovetto, A.; Tamburini, A.; Sandrucci, A. 2001. Effects of a nonforage diet on milk production, energy, and nitrogen metabolism in dairy goats throughout lactation. J. Dairy Sci., 84:2450–2459. [ Links ]
6. Blanchard, N. 2001. Avances de la explotación caprina en Venezuela y pertinencia de su desarrollo. III Congreso Nacional y I Congreso Internacional de Ovinos y Caprinos. Universidad Central de Venezuela. Facultad de Agronomía. Maracay. Venezuela. pp. 25 – 34. [ Links ]
7. Botnick, M. 1994. The Basics of Digestion and Feeding of Goats. The Homesteaders Connection, Dirección Electrónica: http://www.psmag.com/HC/. Consultado el 10/01/03. [ Links ]
8. Browning, R. Jr.; Leite-Browning, M. L.; Sahlu, T. 1995. Factors affecting standardized milk and fat yields in Alpine goats. Small Rumin. Res., 18:173-178. [ Links ]
9. Bruckmaier, R.; Blum, J. 1998. Oxytocin release and removal in ruminants. J. Dairy Sci., 81:939 - 949 [ Links ]
10. Capote, J. 1999. The effects of milking once or twice daily throughout lactation on milk production of Canarian dairy goats. Milking and milk production of dairy sheep and goats (1999). EAAP publication 45:267-273. [ Links ]
11. Capote, J.; López, J. L.; Darmanin, N.; Peris, S.; Argüello, A. 1995. Influencia del doble ordeño en la producción lechera de las cabras Canarias (Tipo Tinerfeño). XX Jornadas Científicas de la Sociedad Española de Ovinotecnia y Caprinotecnia, 25-27 de septiembre, Madrid (España). [ Links ]
12. Centre International Caprin. 2002. Contrôle Laitier. Dirección electrónica: http://www.chevres-sur-le-web.com/. Consultada el 15/05/2004. [ Links ]
13. Chilliard, Y.; Ferlay, A.; Rouel, J.; Lambert, G. 2003. A review of nutritional and physiological factors affecting goat milk lipid synthesis and lipolysis. J. Dairy Sci., 86:1751-1770. [ Links ]
14. FAO. 2005. Anuarios Estadísticos. Dirección Electrónica: http://www.fao.org. Consultado el 10/03/06. [ Links ]
15. Fernández, G. 2000. Parámetros productivos de cabras Pardo Alpinas y sus cruzas, bajo régimen de pastoreo. Producción Latina, XXV: 541-544. [ Links ]
16. Fernández, C.; Sánchez, A.; Martí, A.; Perea, C. 2001. Modelización de la curva de lactación según el número de crías nacidas en cabras Murciano-Granadinas. XXVI Jornadas Científicas y V Internacionales de la Sociedad Española de Ovinotécnia y Caprinotécnia. Ed. Junta de Andalucía. Consejería de Agricultura y pesca, España. pp. 897-902. [ Links ]
17. Gamarra, Y. 2006. Evaluación de algunos factores que afectan la producción de leche y la duración de la lactancia en cabras mestizas de la raza canaria con cabras locales. Tesis para optar al titulo de Magíster Scientiarium en Producción de Ovinos y Caprinos. Universidad Centrooccidental Lisandro Alvarado. Decanato de Ciencias Veterinarias. Cabudare. Venezuela. 36 p. [ Links ]
18. García–Bojalil, C. 1992. Manejo y alimentación de la vaca lechera durante el periodo seco y sus implicaciones durante el postparto. Manual de Asistencia Técnica. Guía Agropecuaria de Venezuela. Editorial Agropecuaria C.A. Maracaibo, Venezuela. [ Links ]
19. García-Hernández, R.; Newton, G.; Horner, S.; Nuti, L. C. 2007. Effect of photoperiod on milk yield and quality, and reproduction in dairy goats. Livestock Sci., 110:214-220. [ Links ]
20. Gipson, T. A.; Grossman, M. 1990. Lactation curves in dairy goats: a review. Small Rum. Res., 3:383. [ Links ]
21. Haenlein, G. 1996. Goat Management. Dirección electrónica: http://ag.udel.edu/extension/information/goatmgt/gm-list.htm. Consultado el 01/10/2004. [ Links ]
22. Henderson, A. J.; Peaker, M. 1987. Effects of removing milk from the mammary ducts and alveoli, or of diluting stored milk, on the rate of milk secretion in the goat. Quart. J. Exp. Physiol., 72: 13-19. [ Links ]
23. Hernández, I. 2002. Ramoneo de las cabras en un bosque seco tropical: Especies consumidas y su valor nutritivo. Dirección electrónica: http://capra.iespana.es/capra/ramoneo.htm. Consultado el 01/10/2006. [ Links ]
24. Hervé, A.; Sigwald, J. 2001. Résultats 2000 du contrôle laitier. Réussir La Chèvre, 246:33-34. [ Links ]
25. Infante, D.; Tormo, R.; Conde, M. 2003. Empleo de la leche de cabra en pacientes con alergia a las proteínas de la leche de vaca. An. Pediatr.,59:138-142. [ Links ]
25. Jimeno, V.; Rebollar, P.; Castro, T. 2003. Nutrición y alimentación del caprino de leche en sistemas intensivos de explotación. XIX Curso de Especialización FEDNA. Madrid, 23 y 24 de octubre de 2003. pp.155-178. [ Links ]
26. Jordana, J.; Amills, M.; Diaz, E.; Angulo, C.; Serradilla, J. M.; Sanchez, A. 1996. Gene frequencies of caprine alpha-s-1-casein polymorphism in Spanish goat breeds. Small Rumin. Res., 20:215-221. [ Links ]
27. Kawas, J.; Lopes, J.; Danelon, D.; Lu, C. 1991. Influence of forage-to-concentrate ratios on intake, digestibility, chewing and milk production of dairy goat. Small Rumin. Res., 4:11-18. [ Links ]
28. Knight, C. H. 1992. Milk yield responses to sequential treatments with recombinant bovine somatotrophin and frequent milking in lactating goats. J. Dairy Res., 59, 115-122. [ Links ]
29. Le Jaouen. J. C. 1986. Composition du lait : la situation actuelle. La Chèvre, 153:10-13. [ Links ]
30. Lu, C. D. 1989. Effects of heat stress on goat production. Small Rumin. Res., 2:151-162. [ Links ]
31. Lu, C. D.; Potchoiba, M. J.; Loetz, E.R. 1991. Influence of vacuum level, pulsation ratio and rate on milking performance and udder health in dairy goats. Small Rumin. Res., 5:1-8. [ Links ]
32. Martin, P. 1993. Genetic polymorphism of goat milk proteins. Lait. 73:511-532. [ Links ]
33. Milerski, M.; Mare, V. 2001. Analysis of systematic factors affecting milk production in dairy goat. Acta Univ. Agric. et silvic. Mendel. Brun (Brno).1:43-50. [ Links ]
34. Mocquot, J. C. 1978. Effets de lomission régulière et irrégulière dune traite sur la production laitière de la chèvre. II Symposium International sur la Traite Mécanique des Petits Ruminants, 22-27 mai, Alghero (Italie), pp. 175-201. [ Links ]
35. Mocquot, J. C. ; Auran, T. 1974. Effets de différentes fréquences de traite sur la production laitière des caprins. Ann . Génét. Sél. Anim., 6:463-476. [ Links ]
36. Mocquot, J. C. ; Guillimin, P. 1975. Effets de différentes fréquences de traite sur la production laitière des chévres. 1ères Journées de la Recherché e Ovine et Caprine, 2-4 Déc., pp. 108-116. [ Links ]
37. Moioli, B. ; Pilla, F. ; Tripaldi, C. 1998. Detection of milk genetic polymorphisms in order to improve dairy traits in sheep and goats: A review. Small Rumin. Res., 27:185-195. [ Links ]
38. Morand-Fehr, P. 2005. Recent developments in goat nutrition and application: A review. Small Rumin. Res., 60:25–43. [ Links ]
39. Muñoz, G. 1997. Comportamiento productivo y reproductivo en un rebaño caprino experimental en el Estado Lara, Venezuela. Tesis para optar al titulo de Magíster Scientiarium en Producción Animal. Mención Genética. Universidad Central de Venezuela. Facultades de Agronomía y Ciencias Veterinarias. Maracay. Venezuela. 134 p. [ Links ]
40. Oliver, F.; Pérez-Guzmán M.D.; Pérez E.M.; Montoro V. 2001. Estudio de la influencia de la edad al primer parto sobre la producción lechera de cabras de raza Murciano-Granadona en Castilla-La Mancha. XXVI Jornadas Científicas y V Internacionales de la Sociedad Española de Ovinotecnia y Caprinotecnia. Ed. Junta de Andalucía. Consejería de Agricultura y pesca. pp. 897-902. [ Links ]
41. Paapa, M.; Capuco, J.; Lefcourt, A.; Burvenich, C.; Miller, R.H. 1992. Physiological response of dairy cows to milking. En: Proceedings International Symposium on Prospects for Automatic Milking. A.H. Lipema ed., PUDOC Sci. Publ. Wageningen, EAAP 65:93-105. [ Links ]
42. Pacheco, F. ; Monteiro, A. ; Lopes, Z. ; Barros, M. 1998. Contrôle laitier caprin dans la région du Minho (Portugal). En: Milking and milk production of dairy sheep and goats. Proceedings International Symposium on the Milking of Small Ruminants, Athens, Greece. EAAP. 95:460-462. [ Links ]
43. Park, Y. 2007. Rheological characteristics of goat and sheep milk. Small Rumin. Res., 68:73-87. [ Links ]
44. Park, Y.; Juárez, M.; Ramos, M.; Haenlein, G. 2007. Physico-chemical characteristics of goat and sheep milk. Small Rumin. Res., 68: 88-113. [ Links ]
45. Peris, S.; Such, X.; Caja, G. 1998. Características de los sistemas de ordeño en ganado caprino y su relación con el estado sanitario de la ubre. Revista OVIS Nº 54: Mamitis caprina Tomo II. D. Capítulo I. Enero. pp. 11-23. [ Links ]
46. Pinkerton, F. 1987. Tecnología de la Producción Caprina. FAO. Oficina Regional para América y el Caribe. Programa de Cooperación Técnica. Chile. 242 p. [ Links ]
47. Preston, T. 2004. Estrategia nutricional para la producción caprina. IV Congreso Nacional de Ovinos y Caprinos. UNEFM. Santa Ana de Coro. 14 al 17 de julio. pp. 133-140. [ Links ]
48. Remeuf, F.; Lenoir, J. 1986. Relationship between the physicochemical characteristics of goat milk and its rennet ability. FIL-IDF Bulletin 202:68-72. [ Links ]
49. Renner, E. 1983. Milch und Milchprodukte in der Ernaehrung des Menschen. VV-GmbH Volkswirtsch. Verlag, Munich, 467 p. [ Links ]
50. Rodríguez – Parra, M.; Caja, G. 2004. Comparación de la ingestión voluntaria de forrajes en ovino y caprino. IV Congreso Nacional de Ovinos y Caprinos. UNEFM. Santa Ana de Coro. 14 al 17 de julio. pp. 24. (Resumen). [ Links ]
51. Salama, A.; Such, X.; Caja, G.; Rovai, M.; Casals, R.; Albanell, E.; Marin, M.; Marti, A. 2003. Effects of once versus twice daily milking throughout lactation on milk yield and milk composition in dairy goats. J. Dairy Sci., 86:1673-1680. [ Links ]
52. Salama, A. 2005. Modificación de la Curva de Lactación en Cabras Lecheras: Efectos de la frecuencia de ordeño, el periodo de secado y el intervalo entre partos. Tesis Doctoral. Departament de Ciència Animal i dels Aliments de la Universitat Autonoma de Barcelona, Bellaterra. 163 p. [ Links ]
53. Salvador, A. 1998. Mantenga sus proporciones en el rebaño. Revista Venezuela Bovina. 13:50-52. [ Links ]
54. Salvador, A.; Martinez, G.; Alvarado, C.; Hahn, M. 2006. Composición de leche de cabras mestizas Canarias en condiciones tropicales. Zootecnia Trop., 24:307-320. [ Links ]
55. Sánchez, I.; Martínez, R.; Torres, G.; Becerril, C.; Mastache, A.; Suárez, J.; Rubio, M. 2006. Producción de leche y curvas de lactancias en tres razas de cabras en el trópico seco de México. Vet. Méx., 37:493-502. [ Links ]
56. Sanz Sampelayo, M.; Pérez, M.L.; Gil Extremera, F.; Boza, J. J.; Boza, J. 1999. Use of different dietary protein sources for lactating goats: milk production and composition as functions of protein degradability and amino acid composition. J. Dairy Sci., 82:555–565. [ Links ]
57. Sahlu, T.; Goetsch, A.; Luo, J.; Nsahlai, I.; Moored, J.; Galyean, M.; Owensf, F.; Ferrell, C.; Johnson, Z. 2004. Nutrient requirements of goats: developed equations, other considerations and future research to improve them. Small Rumin. Res., 53:191–219. [ Links ]
58. Simos, E.; Voutsinas, L. P.; Pappas, C.P. 1991. Composition of milk of native Greek goats in the region of Metsovo. Small Rumin. Res., 4:47-60. [ Links ]
59. Voutsinas, L.; Pappas C.; Katsiari M. 1990. The composition of Alpine goats milk during lactation in Greece. J. Dairy Res., 57: 41-51. [ Links ]
60. Wilde, C. J.; Henderson, A. J.; Knight, C. H.; Blatchford, D. R.; Faulkner, A.; Vernon, R. G. 1987. Effects of long-term thrice-daily milking on mammary enzyme activity, cell population and milk yield in the goat. J. Anim. Sci., 64:533-539. [ Links ]
61. Zeng, S.S.; Escobar E.N.; Popham T. 1997. Daily variations in somatic cell count, composition, and production of Alpine goat milk. Small Rumin. Res., 26:253-260. [ Links ]
62. Zullo, A.; Barone, C. M.; Chianese, L.; Calatruglio, P.; Occidente, M.; Matassino, D. 2005. Protein polymorphisms and coagulation properties of Cilentana gota milk. Small Rumin. Res., 58:223- 230. [ Links ]
63. Zumbo, A.; Chiofalo, B.; Liotta, L.; Rundo Sotera, A.; Chiofalo, V. 2004. Quantitative and qualitative milk characteristic of Nebrodi goats. South African J. Anim. Sci., 34 (Suplement 1):155-157. [ Links ]

