










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkArchivos Latinoamericanos de Nutrición
versión impresa ISSN 0004-0622versión On-line ISSN 2309-5806
ALAN v.58 n.2 Caracas jun. 2008
Papel de las poliaminas en la alimentación. Importancia de las poliaminas en la alimentación infantil
Carlos Gómez Gallego, Gaspar Ros Berruezo, María José Bernal Cava, Darío Pérez Conesa, María Jesús Periago Castón
Departamento de Tecnología de los Alimentos, Nutrición y Bromatología. Facultad de Veterinaria. Universidad de Murcia. Campus de Espinardo. España. Instituto de Nutrición Infantil Hero. Hero España SA, España
Resumen.
Durante la década de los 90 del siglo pasado comenzaron los primeros estudios que señalaban a las poliaminas como importantes factores de crecimiento de la leche materna. Sin embargo, todavía se desconoce mucho sobre el papel que desempeñan en la alimentación infantil y cual sería la ingesta recomendada para este grupo poblacional. En los últimos años va siendo cada vez mayor la atención que muestra la comunidad científica internacional hacia las poliaminas, debido no sólo al importante papel que desempeñan en el metabolismo celular, sino por su posible participación en diversas patologías y durante el desarrollo del organismo. Sería recomendable, teniendo en cuenta que el contenido en poliaminas de las fórmulas infantiles es unas 10 veces menor al de la leche materna, profundizar más en este campo, con el fin de garantizar una correcta alimentación durante la etapa de lactancia.
Palabras clave: Putrescina, espermidina, espermina, poliaminas, fórmulas infantiles.
Summary.
Role of polyamines in diet. Importance of polyamines in infant nutrition. The first studies indicating polyamines as important growth factors in breast milk began during the nineties of last century. Nevertheless, it is still not well known the role they play in infant nutrition or what the recommended intake would be for this population group. In recent years, there has been increased attention of the international scientific community towards polyamines, not only due to the important role they play in the cellular metabolism, but also to their possible implication in some diseases and during the development of the human organism. Bearing in mind that the content in polyamines of the infant formula is around tenfold less than in breast milk, it would be recommended to gain insight into this theme in order to guarantee correct nutrition during lactation.
Key words: Putrescine, spermidine, spermine, polyamines, infant formula.
Recibido: 06-01-2008 Aceptado: 15-04-2008
INTRODUCCION
Con el nombre genérico de poliaminas se conoce a una familia de compuestos cuyas características comunes son poseer una estructura simple alifática de bajo peso molecular (1), ser policationes flexibles que a pH fisiológico se encuentran presentes casi exclusivamente en sus formas di, tri y tetraamonio protonadas (1,2) y ser compuestos muy estables capaces de resistir el calor y condiciones ácidas o alcalinas (3).
Los miembros más representativos de esta familia son la putrescina, la espermina y la espermidina, aunque otros autores incluyen a la cadaverina por estar presente de forma natural en muchos organismos vivos (4).
Recientemente, una nueva molécula derivada de la arginina, denominada agmatina, ha sido identificada como poliamina. Es una amina policatiónica que es sintetizada por descarboxilación de la L-arginina y que fue originalmente identificada como neurotransmisor (5).
Actualmente se siguen descubriendo otras poliaminas con estructuras químicas diferentes o en forma cíclica, con distintas funciones. Así, se ha descrito toda una gama de poliaminas macrocíclicas con funciones de defensa en insectos (4).
Las poliaminas fueron descubiertas por Antonie van Leeuwenhoek en 1678, que, al observar al microscopio muestras de semen desecadas, las vio formando cristales de espermina fosfato. Sin embargo, no fue hasta 1924 cuando se dedujo la fórmula empírica de los cristales, y dos años después se consiguieron sintetizar químicamente por primera vez. Los nombres de la espermidina y la espermina reflejan claramente su descubrimiento original. La putrescina fue aislada por primera vez a partir de Vibrio cholerae, pero su nombre común deriva de las grandes cantidades que se encuentran en la carne putrefacta. Debido a sus orígenes puede sorprender que a día de hoy las poliaminas sean consideradas como reguladores críticos del crecimiento, diferenciación y muerte celular (6).
El metabolismo de las poliaminas ha sido cuidadosamente conservado a lo largo de la evolución de los seres vivos, existiendo varios mecanismos de regulación que contribuyen a mantener la homeostasis intracelular de las poliaminas. (7).
Existen tres fuentes de poliaminas para el organismo: biosíntesis in situ a partir de aminoácidos, ingestión directa en la dieta y síntesis y liberación por la flora residente en el tracto gastrointestinal (3).
Aunque las células pueden sintetizar poliaminas, la dieta parece ser una fuente fundamental, porque al igual que en el caso de ciertos aminoácidos semiesenciales, la capacidad de síntesis de las células o los órganos no es suficiente para satisfacer los requerimientos totales (3). El contenido en las diferentes poliaminas varía de acuerdo al tipo de alimento; de forma general, en alimentos vegetales el contenido en espermidina es mayor que el de espermina. En alimentos de origen animal la relación espermidina/espermina es la contraria, siendo mayor la cantidad de espermina. (8). Con respecto a la ingesta de poliaminas, hay dos hechos que acontecen en nuestro país: hay pocos estudios de composición de poliaminas en los alimentos y sólo se han realizado para quesos, carnes, embutidos y algunos vinos mediante técnicas de cuantificación por HPLC; y no hay datos sobre su consumo poblacional (9). Por otro lado, a pesar del hecho de que nosotros ingerimos cantidades significativas de poliaminas, la biodisponibilidad en el hombre no es del todo conocida (6)
Su alta concentración en el fluido espermático y en la sangre, su posible papel en el desarrollo del cáncer y en su tratamiento; su papel en la nutrición y el desarrollo, sobre todo en etapas de crecimiento intenso; y su papel como inhibidores de la glicosilación no enzimática, hacen a estos compuestos muy interesantes desde el punto de vista de la investigación.
Función de las poliaminas
Desde las décadas de los 80 y 90 se sabe que las poliaminas están implicadas en varios procesos de crecimiento y diferenciación celular. El importante papel de las poliaminas aparece reflejado en la Figura 1, siendo fundamental en el crecimiento normal y adaptativo de varios sistemas de órganos, lo que está bien documentado. Además, las poliaminas están esencialmente implicadas en la carcinogénesis y el crecimiento de tumores (7).
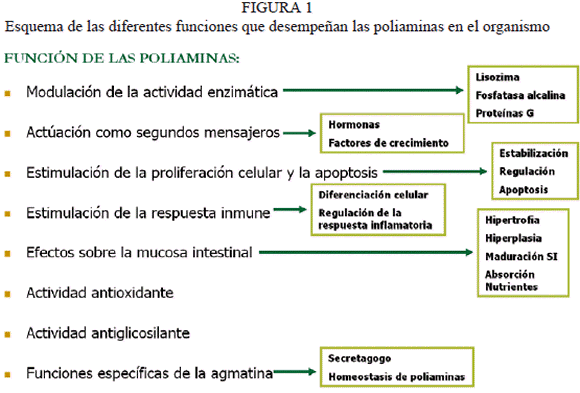
A pH fisiológico, las poliaminas poseen carga positiva en cada uno de sus átomos de nitrógeno. Sin embargo, la carga de las poliaminas se distribuye a lo largo de la cadena carbonada, lo que las convierte en unas moléculas únicas y distintas desde el punto de vista electroquímico al resto de cationes polivalentes celulares (6). Además, su función depende de las cargas eléctricas, ya que la energía de unión a moléculas disminuye conforme lo hace el número de cargas. Así, se ha visto que la espermidina es la más activa y la putrescina es la menos activa en el control de varios procesos biológicos (5), aunque esto no siempre es así.
Las poliaminas modulan la actividad de algunas enzimas, como la lisozyma, aumentando su actividad y su estabilidad, y de la fosfatasa alcalina (por inducción de modificaciones postranscripcionales) haciendo que aumente su actividad en el yeyuno y disminuya en el íleon (10). Además, las poliaminas están implicadas en la regulación de bastantes enzimas de unión a membrana, incluyendo a (6,11): adenilato ciclasa, proteínas G, transglutaminasa tisular y varios canales iónicos como: el de N-metil-D-aspartato, los KIR y los canales de calcio activados por voltaje.
Efectos sobre la proliferación celular y la apoptosis
La estimulación de la proliferación celular parece estar relacionada con la estabilización de las cargas negativas del ADN y con la estabilización de la estructura de la cromatina.
Parece ser que también regulan algunos factores transcripcionales y la síntesis de proteínas. Sin embargo, la acumulación excesiva de poliaminas induce la apoptosis, debido probablemente al incremento del estrés oxidativo asociado a la acumulación de peróxido de hidrógeno durante el catabolismo de las poliaminas por la poliamino oxidasa (5). Además, las células pueden sufrir apoptosis cuando sus reservas de poliaminas se agota (12).
Estos efectos sobre la proliferación celular son debidos a que las poliaminas neutralizan las cargas negativas de los ácidos nucleicos y favorecen que el ADN adopte una conformación apropiada (11), se ha visto que la espermidina facilita la compactación del ADN y la actividad de las topoisomerasas, favorece su resistencia a la ADNasa I y a otras nucleasas, al mismo tiempo que lo mantienen transcripcionalmente activo (4), la espermidina y la espermina pueden unir las hendiduras mayores y menores del ADN, actuando como un cepo entre dos moléculas diferentes o dos partes alejadas de la misma molécula (6) y, por otro lado, los efectos proliferativos de las poliaminas pueden ser debidos también a la estimulación directa de la síntesis de proteínas a nivel de la estimulación de la maduración de la subunidad 30S del ribosoma y a nivel de la traducción de muchas proteínas importantes para el crecimiento celular y la viabilidad. Además, la espermidina es imprescindible para la hipusilación del factor iniciador de la traducción de eucariotas 5A (eIF-5A). (13,14). La estructura del eIF-5A y su modificación están altamente conservadas desde levaduras a humanos e incluso arqueas. En levaduras, un fallo al sintetizar este factor proteico es letal (15).
Sin embargo, la función más importante de las poliaminas, en la que no pueden ser sustituidas por otras moléculas cargadas positivamente, es su acción como segundos mensajeros, mediando en la acción de todas hormonas conocidas y factores de crecimiento, jugando un papel indispensable en el metabolismo humano (16).
Efectos sobre el sistema inmune (SI).
Las poliaminas están implicadas en la diferenciación de las células del sistema inmune y en la regulación de la respuesta inflamatoria, con cambios en los niveles de IgA e incremento en el porcentaje de linfocitos intraepiteliales (5).
Se ha visto que las poliaminas pueden actuar como intermediarios en el efecto timotrófico de mediadores como la prolactina. También, en el curso de la inflamación local, la espermina es liberada por las células dañadas o muertas favoreciendo la migración y el crecimiento de linfocitos (11).
Efectos sobre la mucosa intestinal
La acción de las poliaminas sobre el crecimiento, desarrollo y la permeabilidad intestinal no es del todo conocida y suscita controversia. Al parecer pueden ejercer efectos tróficos directos sobre las células intestinales inmaduras, mejorando la permeabilidad de la mucosa y produciéndose de forma paralela una maduración del sistema inmune intestinal, afectando particularmente a la diferenciación de la población linfocitaria intraepitelial (17).
Se ha visto que en ratas lactantes las poliaminas producen un incremento de todos los parámetros de crecimiento de la mucosa, inducen prematuramente un patrón adulto de las enzimas de la mucosa (lactasa, sacarasa, maltasa y aminopeptidasa) e incrementan la concentración de inmunoglobulinas en las vellosidades y en las criptas (18).
Se ha visto que además de hipertrofia de las células intestinales producen hiperplasia, estimulando la regeneración del tejido. Inhibe la secreción ácida gástrica, promueve la curación de las úlceras y previene el desarrollo de úlceras gástricas inducidas experimentalmente. Además de estimular el desarrollo intestinal favorecen e incrementan la absorción de nutrientes (19).
De forma más concreta se ha visto en ratas que la espermina y espermidina a nivel de intestino incrementan la expresión enzimática, la síntesis de ADN y proteínas, al mismo tiempo que reducen la permeabilidad a macromoléculas (20).
La inducción de la proliferación y la maduración del tracto intestinal parece ser, al menos en parte, debida a cambios en la composición lipídica de la membrana y la fluidez (5).
Actividad antioxidante de las poliaminas. Muchos estudios han demostrado que la actividad antioxidante de las poliaminas y de algunas aminas biógenas fenólicas puede ser incluso más fuerte que las de los tocoferoles, el ácido ascórbico, el hidroxianisol butilado (BHA) y el hidroxitolueno butilado (BHT) (21).
Se ha visto que las células que tienen sus reservas de poliaminas agotadas son altamente sensibles al ataque oxidativo, lo que sugiere que las poliaminas son importantes antioxidantes fisiológicos. Además, se ha visto también que las células responden al estrés oxidativo incrementando la biosíntesis de poliaminas y su captación (22).
Fisiológicamente se viene pensando que la capacidad de las poliaminas de captar las diferentes especies de oxígeno reactivas parece tener una importancia menor porque la constante de esa reacción es baja, y se asume que sus propiedades antioxidantes podrían depender sobre todo de su habilidad para quelar metales (6). De forma general, se ha visto que la putrescina no reacciona con los radicales hidroxilo y la reactividad de la espermidina es menor que la del resto de poliaminas con dos grupos aminopropilo terminales (22). Así, las poliaminas naturales tienen menor capacidad para captar radicales peroxilo y anión superóxido que otros antioxidantes. Sin embargo no debemos despreciar esta capacidad debido a la alta concentración local de poliaminas alrededor de las dianas aniónicas de las especies de oxígeno reactivas como el ADN y las membranas celulares (6).
Actividad antiglicosilante de las poliaminas La glicosilación no enzimática de proteínas y ácidos nucleicos es la base de las complicaciones de la diabetes debidas a la hiperglucemia. Esta glicosilación no enzimática forma, en una primera etapa, productos de glicosilación tipo Amadori o fructosaminas. En una segunda etapa se forman los productos de glicosilación avanzada (AGEs), que son los responsables de las retinopatías, neuropatías y nefropatías diabéticas, y están implicados también en patologías como la uremia y la aterosclerosis (12).
Existían bastantes indicios que hacían de las poliaminas buenos candidatos para inhibir esta glicosilación como: su naturaleza policatiónica, su ubicuidad, su papel esencial en el crecimiento, su relativamente alta concentración en algunos tejidos (2 nmol/l en la piel) y su alta concentración en el semen (10 mmol/l), un fluido que contiene alrededor de 10 mmol/l de fructosa (un muy agresivo agente glicosilante) (12).
Se ha demostrado que las poliaminas inhiben la glicosilación ya que se ha podido comprobar que la espermina inhibe el avance de la glicosilación en histonas y en la ubiquitina, y que inhibe la glicosilación inducida experimentalmente por el metilglioxal. Además, se ha visto que la espermina, la espermidina, la carnosina y la aminoguanidina actúan como inhibidores de los AGEs (12).
Estos efectos han sido demostrados a las concentraciones fisiológicas in vitro. Otros argumentos que apoyan esta hipótesis son: la alta concentración de espermina en el núcleo, lo que permite proteger al ADN y a las histonas, la inducción de la ornitina descarboxilasa por la radiación UVB y el estrés oxidativo, y la disminución de los niveles de poliaminas y el aumento de la glicosilación con la edad (12).
Funciones específicas de la agmatina
Debido a sus peculiaridades la agmatina tiene funciones diferentes al resto de poliaminas (11): actúa como un secretagogo, modulando la liberación de insulina y el metabolismo de la glucosa. Estimula la liberación de insulina en función de la dosis. Además, se ha visto que, in vitro, estimula la secreción de adrenalina y noradrenalina en el sistema nervioso central estimula la liberación de la hormona liberadora de hormona luteinizante. Además, en función de la dosis, incrementa el efecto analgésico de la morfina. Se une al receptor á-2 adrenérgico, al receptor de imidazolina y al receptor NMDA, y activa la óxido nítrico sintasa de células endoteliales. La agmatina juega un importante papel en la homeostasis de las poliaminas por inhibición de la ornitina descarboxilasa e incremento de la actividad y la síntesis de la espermidina/espermina acetil transferasa.
Poliaminas y cáncer
Desde los años 50 se conoce la relación entre la enfermedad de cáncer y las poliaminas, estando bien demostrado que la putrescina se acumula en hematomas de rápido desarrollo, y que la tasa de crecimiento depende de la relación entre las concentraciones de espermidina y espermina. Oncogenes como ras, jun y fos inducen la transformación de células normales a células tumorales por una vía relacionada con la biosíntesis de poliaminas (23). Debido a esto muchos investigadores han tratado de buscar en las poliaminas un marcador temprano de la enfermedad.
Se ha demostrado un incremento en tejido tumoral tanto de la actividad enzimática de la ornitina descarboxilasa como de la expresión del ARNm de la ornitina descarboxilasa, incremento que está directamente correlacionado con el grado de malignidad del tumor. También cabe destacar la capacidad de las poliaminas para regular la expresión de protooncogenes. Tienen un interés especial los estudios que atribuyen un comportamiento protooncogénico al gen de la ornitina descarboxilasa, estando demostrado que una vez inducida su sobreexpresión, es capaz de provocar un desarrollo tumoral (2).
Se ha visto que entre el 70%-90% de los pacientes con cáncer excretan poliaminas conjugadas (acetilespermidina ) en la orina. El problema es que no se pueden usar como indicador, ya que algunas personas de manera normal o en determinados estados fisiológicos no patológico (como puede ser el caso de las embarazadas) excretan poliaminas en la orina, pudiendo aparecer entorno a un 20% de falsos positivos. Por otro lado, sí que podrían utilizarse como buenos marcadores para hacer el seguimiento de la enfermedad ya que los niveles de espermidina y espermina eritrocitaria aumentan conforme progresa el tumor. También serían un buen marcador en el caso de los cánceres recurrentes, ya que un aumento en los niveles de poliaminas se observa 1-6 meses antes de que aparezcan los primeros signos clínicos y radiológicos (23).
Respuesta al estrés mediada por poliaminas
La respuesta general al estrés se activa por la vía de los segundos mensajeros y responde al denominado stress program intracelular. En ella se observa, entre otras alteraciones metabólicas, el incremento transitorio del metabolismo de las poliaminas denominado por Gilard en 1992 como poliamyne-stress-response (PSR). Esta respuesta cerebral lleva implícita la persistente acumulación de putrescina y una eventual reducción de espermidina y espermina. Se ha postulado que una correcta regulación del metabolismo cerebral de las poliaminas es la mejor respuesta a los agentes estresantes. Situaciones de estrés durante los primeros periodos de vida alteran el normal desarrollo cerebral que posteriormente provocan alteraciones que afectan al sujeto adulto. Especial interés despierta la inhibición de la actividad cerebral de la ornitina descarboxilasa en respuesta al estrés que supone la separación maternal en el recién nacido (2).
Contenido en poliaminas de los alimentos
En muchas situaciones, en las cuales se requiere un crecimiento intensivo, los requerimientos de poliaminas podrían ser grandes, siendo necesaria una alta ingesta de poliaminas en la dieta. Por tanto, es importante establecer los niveles de estos compuestos en los alimentos y sus requerimientos dietéticos, para así, poder definir dietas restrictivas o ricas en poliaminas cuando se requiera.
Las poliaminas se encuentran en los alimentos en dos formas: libres y conjugadas. Mayoritariamente se encuentran conjugadas en alimentos de origen vegetal, principalmente unidas a compuestos fenólicos, y pueden ser hidrolizadas con un ácido fuerte. Hasta ahora, en la mayoría de artículos publicados no se diferencia entre las diferentes formas de las poliaminas (8, 24).
Durante el procesado de los alimentos se pueden dar cambios en el contenido de las diferentes poliaminas. Así, el contenido en poliaminas aumenta durante los procesos fermentativos o la maduración (24).
Se han observado cambios en las poliaminas durante el almacenamiento de productos frescos a 5ºC. En ellos, aumenta el contenido en putrescina durante este periodo de 3 a 8 veces, de forma paralela al aumento de los recuentos de microorganismos aeróbicos; las otras poliaminas no cambian significativamente, aunque algunos autores citan disminuciones en los niveles de espermina a partir de los 36 días de almacenamiento. En cuanto al tratamiento térmico no debería variar el contenido en poliaminas, aunque se ha visto que en el tostado del café se da una completa pérdida de la putrescina y la espermidina, y se observa una considerable disminución del contenido en espermina (8), quizás no debido al tratamiento térmico en sí, si no a que se vean favorecidas determinadas reacciones enzimáticas de degradación.
El contenido en poliaminas en los alimentos es muy variado, siendo los alimentos con mayores niveles de poliaminas el germen de trigo, arroz integral, arroz salvaje, mango filipino, pimienta verde, calabaza japonesa, nueces, encurtidos fermentados y quesos, eperlano de charca, buccino, huevas de salmón saladas, anguila, la carne, granos de soja fermentados o no, champiñones, té verde y las naranjas (14).
Como se mencionó previamente, el contenido en las diferentes poliaminas varía de acuerdo al tipo de alimento; de forma general, en alimentos vegetales el contenido en espermidina es mayor que el de espermina. En alimentos de origen animal la relación espermidina/espermina es la contraria, siendo mayor la cantidad de espermina. (8). Las setas contienen gran cantidad de espermidina, las naranjas más cantidad de putrescina y las hojas de té contienen de las tres poliaminas en gran cantidad. La carne roja contiene mayoritariamente espermina. La leche y los huevos contienen proporcionalmente menos cantidades. La predominante en la leche es la espermidina mientras que los derivados como el queso o provenientes de la fermentación microbiana contienen mayor cantidad de putrescina y espermidina. El vinagre y varias clases de bebida alcohólicas contienen también cantidades elevadas. El sake contiene bastante agmatina que proviene de la descarboxilación de la arginina que, seguramente, se produce en el proceso de fermentación por las bacterias acidolácticas o reductoras del ácido nítrico (2).
Importancia dietética de las poliaminas Como se ha visto, las poliaminas son esenciales para el metabolismo celular, el crecimiento y la renovación de tejidos. Este requerimiento, como es lógico, está incrementado en tejidos con una alta tasa de crecimiento. Aunque las células pueden sintetizar poliaminas, la dieta parece ser una fuente esencial, porque al igual que en el caso de ciertos aminoácidos semiesenciales, la capacidad de síntesis de las células o los órganos no es suficiente para satisfacer los requerimientos totales (3).
Con respecto a la ingesta de poliaminas, hay dos hechos que acontecen en nuestro país: hay pocos estudios de composición de poliaminas en los alimentos y sólo se han realizado para quesos, carnes, embutidos y algunos vinos mediante técnicas de cuantificación por HPLC; y no hay datos sobre su consumo poblacional (9). Se asume que la dieta occidental suple cantidades del orden de cientos de nanomoles de poliaminas diarias. Así el aporte de poliaminas en el Reino Unido se considera del orden de 388 nmol/día repartido del siguiente modo: 57% de putrescina, 26% de espermidina y 18% de espermina. (2).
Esta disminución se ha comprobado, no sólo en roedores sino también en suero y orina de humanos saludables (12). A causaLos requerimientos de poliaminas son altos en las etapas de crecimiento intenso, por lo que las cantidades suplidas en la dieta juegan un papel importante en la infancia y adolescencia. En los adultos sanos los requerimientos son inferiores ya que sólo se necesitan para el reemplazamiento celular o bien como mediadores de la respuesta hormonal o de factores de crecimiento (2,19).
Dietas deficientes en poliaminas producen en roedores pérdida de peso, infertilidad en hembras, pancreatitis y deterioro del aprendizaje espacial; mientras que dietas ricas en poliaminas se correlacionan con la prevención de la aparición de alergias alimentarias, prevención de las úlceras gástricas y estimulación de la maduración intestinal. Se han demostrado también los efectos antitumorales de los inhibidores de la síntesis de poliaminas (14).
El cálculo de los niveles de poliaminas en los alimentos podría facilitar el cálculo de la ingesta dietética, siendo esencial a la hora de diseñar dietas ricas o pobres en poliaminas. Por otro lado, durante el envejecimiento se ha visto que la inducción hormonal y la actividad de la ornitina descarboxilasa disminuyen, disminuyendo la biosíntesis de poliaminas.
Por otro lado, durante el envejecimiento se ha visto que la inducción hormonal y la actividad de la ornitina descarboxilasa disminuyen, disminuyendo la biosíntesis de poliaminas. Esta disminución se ha comprobado, no sólo en roedores sino también en suero y orina de humanos saludables (12). A causa de la disminución de la actividad de la ornitina descarboxilasa se produce una disminución de la proliferación y del recambio tisular, por lo que el aporte exógeno de poliaminas podría tener un papel importante en las últimas etapas de la vida (2). Estudios realizados con ratas ponen de manifiesto que la disminución en la concentración de poliaminas con la edad no afecta a todos los órganos por igual. El nivel de poliaminas en útero, cerebro y páncreas se mantiene constante; disminuye un poco en el intestino y disminuye significativamente en timo, bazo, ovarios, hígado, estómago, pulmón, riñón, corazón y músculo. El tejido en el que más disminuyen los niveles de poliaminas es el músculo, seguido de riñón, hígado, bazo y timo. Por tanto, para mantener un nivel de poliaminas óptimo es importante ingerir una cantidad suficiente con la dieta, especialmente en personas mayores (14).
Dietas con un alto contenido en poliaminas pueden jugar un papel importante en la recuperación de la microflora y la mucosa intestinal, y la administración de espermina puede mejorar el tratamiento de los traumatismos (5). Como ya hemos señalado antes, la espermidina y la espermina protegen del daño estructural y funcional causado por los productos de glicosilación avanzada en muchas patologías. En experimentos con ratas, unas reservas hepáticas bajas de espermidina y espermina impiden que se inicie el proceso de regeneración tras una hepatoectomia parcial. Un incremento en la ingesta dietética de poliaminas parece tener efectos favorables durante el postoperatorio o durante la curación de heridas (8).
Organos como el tracto gastrointestinal, páncreas y bazo, los cuales tienen una alta tasa de renovación celular, son especialmente dependientes de las poliaminas de la dieta. Además, factores antinutritivos, factores que incrementan la actividad metabólica del estómago por dañar los tejidos (saponinas, taninos ), factores que mimetizan la acción de las hormonas o factores de crecimiento (algunas lectinas), o factores que interfieren con la digestión (como los inhibidores de las enzimas digestivas) van a incrementar la demanda de poliaminas (3).
Por otro lado, se ha visto que altas concentraciones de poliaminas en la dieta pueden incrementar la proliferación de células cancerosas en tumores ya existentes. En estos casos, la combinación de tratamientos farmacológicos con α- difluorometil ornitina (un inhibidor de la ODC), neomicina y dietas con un bajo contenido en poliaminas podría ser crucial en la reducción del crecimiento tumoral (5). Por el contrario, recientemente ha sido demostrado el efecto antiproliferativo de la agmatina en células tumorales. Esto no es debido a la inducción de necrosis celular o apoptosis, sino mediante la inhibición de la síntesis de poliaminas y el transporte por la inducción de la antizyma (11).
A pesar de los efectos beneficiosos de las poliaminas presentes en la dieta es importante tener en cuenta que las poliaminas son precursores potenciales de nitrosaminas, compuestos con acción carcinogénica, al reaccionar con nitritos; y que la putrescina puede potenciar los efectos tóxicos de las aminas biógenas porque interfiere con las enzimas que las metabolizan, inhibiendo o compitiendo a nivel de la monoamino oxidasa, diamino oxidasa e hidroximetil transferasa (24, 25).
Poliaminas en la alimentación infantil Después del periodo de vida intrauterino y por falta de maduración, el recién nacido tiene ciertas funciones biológicas y digestivas limitadas que progresivamente irá desarrollando.
La maduración intestinal ocurre entre la lactancia y el destete, sufriendo el epitelio intestinal de mamíferos cambios como: incremento en la producción de mucus, adaptación inmunológica a nuevos agentes microbianos y nutritivos, adaptación digestiva a nuevos nutrientes, producción de las enzimas necesarias para poder afrontar la alimentación adulta (26).
La naturaleza de las señales que regula la maduración del intestino delgado no es del todo conocida, pero se cree controlada por factores: hormonales, de crecimiento y nutricionales (26).
Está demostrado que las poliaminas son factores de maduración para el intestino delgado y desempeñan funciones relacionadas con la diferenciación celular y la proliferación.
Varios estudios demuestran considerables variaciones individuales en la concentración de poliaminas en la leche humana (Tabla 1). Más allá de las variaciones interespecíficas e individuales, han sido establecidos factores que influyen sobre la concentración de poliaminas, como: factores genéticos, fase de la lactancia, duración de la lactación y cantidad de leche en la glándula mamaria, factores ambientales, estado nutricional y dieta ingerida (sobre todo la ingesta de poliaminas), edad de la madre, origen étnico, ritmo circadiano de la secreción de poliaminas y contaminaciones bacterianas entre otros (7, 17).
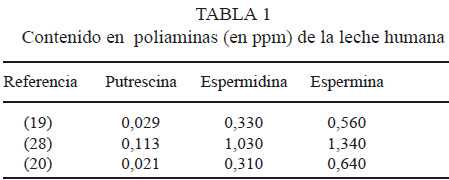
De todos estos factores, uno muy importante es la fase de lactancia. Está comprobado que la leche humana y de rata contiene más espermidina y espermina al final que al principio de la lactación, siendo estables los niveles de putrescina a lo largo de todo el periodo (26). Otros autores han observado que la concentración de poliaminas en la leche humana aumenta durante las primeras semanas tras el parto, alcanzando un máximo después de la primera o segunda semana y declinando progresivamente después (7).
Como ya hemos visto, debido a su alta tasa de proliferación, la mucosa intestinal tiene una alta demanda de poliaminas. Además, la maduración del intestino delgado es un proceso sensible a la nutrición, siendo necesarios una transición consecutiva de diferentes concentraciones de las distintas poliaminas (y de otros factores de la leche) para la completa maduración. La administración oral de espermidina y espermina exógenas tiene como consecuencia una marcada aceleración de la maduración y proliferación de la mucosa (7). Se ha visto que en ratas lactantes producen un incremento de todos los parámetros de crecimiento de la mucosa, inducen prematuramente un patrón adulto de las enzimas de la mucosa (lactasa, sacarasa, maltasa y aminopeptidasa) e incrementan la concentración de inmunoglobulinas en las vellosidades y en las criptas (18)
En la zona apical del enterocito, en las membranas de las microvellosidades, aparecen muchas glicoproteínas que están implicadas en la digestión, el transporte, la detoxificación o en procesos infecciosos. La estructura de las cadenas de glicanos juega un papel importante a la hora de la integración de las proteínas en la membrana, pero el efecto de la estructura del glicano sobre la actividad biológica de la proteína es totalmente desconocido. Se ha visto que en el intestino de ratas lactantes se encuentra una forma activa de la enzima lactasa, mientras que en adultos aparece una forma inactiva en la membrana intracelular con un alto contenido en manosa y mucosa. El tratamiento de ratas lactantes con tiroxina (factor hormonal que regula la maduración) induce la precoz aparición de la forma inactiva fucosilada (26). La forma activa de la lactasa está sialilada mientras que la forma inactiva está fucosilada. La estructura de la cadena de glicano tanto en la lactasa como en la sacarasa podría ser responsable de su integración en la membrana apical del enterocito (27).
La espermidina y la espermina son factores de maduración para muchas glicoproteínas de la membrana del borde en cepillo, tanto en términos de glicosilación como de actividad enzimática. Por tanto, cambios postnatales en la fucosilación de glicoproteínas intestinales podrían ser, al menos parcialmente, consecuencia de un aumento en la ingesta de poliaminas (especialmente espermidina y espermina) durante el periodo de destete (27). Aunque se desconoce en humanos, los cambios en la glicosilación de proteínas parecen ser similares a los de roedores y cerdos, siendo particularmente marcados los cambios en el periodo de destete, un periodo clave para la aparición de las funciones intestinales del adulto (26).
Debido a las variaciones paralelas entre el aumento en los niveles de espermidina y espermina y la galactosilación y la fucosilación de glicoproteínas, es necesario tener en cuenta el papel de las poliaminas en el proceso de maduración intestinal. A parte de los probables efectos directos, las poliaminas podrían también estimular indirectamente la secreción de hormonas o de otros efectores secundarios. Así, la actividad glicosiltransferasa en ratas lactantes es sensible a los glucocorticoides, y se ha sugerido que la espermina puede afectar a la secreción de ACTH (hormona adrenocorticotropa) y corticosterona por la estimulación de la liberación de hormonas esteroideas o a través de mecanismos dependientes de citoquinas. La liberación de corticosteroides va a producir un aumento en la actividad fucosiltransferasa y en la incorporación de fucosa en las membranas de la microvellosidades, lo que va a aumentar la fucosilación de proteínas (26).
Aunque estos estudios se han llevado a cabo en roedores, el papel de las poliaminas de la leche humana en la maduración intestinal de bebés debe tenerse en cuenta, más aún cuando en las fórmulas infantiles en polvo la concentración de poliaminas es aproximadamente 10 veces menor que en la leche humana (26), tal y como se muestra en la Tabla 2.
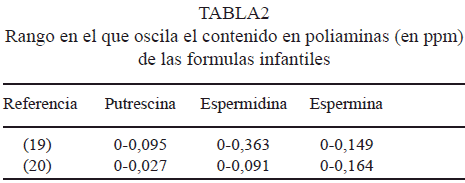
Por otro lado se ha demostrado que la ingesta insuficiente de poliaminas induce sensibilicación a los alérgenos de la dieta. La probabilidad de desarrollar una alergia puede alcanzar el 80% si el contenido medio de espermina es inferior a 0,4 mg/ l y prácticamente no existe si el contenido medio de espermina supera los 2,6 mg/l. La espermina incrementa la proliferación y la diferenciación de linfocitos aislados de niños (8), por lo que sería muy importante tener estos datos en cuenta a la hora de elaborar dichas fórmulas y asegurar una correcta alimentación durante la etapa de lactancia.
Se ha demostrado también, que la administración oral de espermina y espermidina en ratas prematuras aumenta la maduración intestinal, con cambios en la expresión enzimática y en los componentes secretados (como la IgA). El efecto es dependiente de la dosis y específico (ya que la administración de ornitina y arginina no tiene el mismo efecto). El problema es que en estos estudios la dosis administrada era mucho mayor que la que las ratas ingieren por la leche materna. Aun así, asumiendo que los recién nacidos consumen unos 600-700 ml de leche al día (unos 3,5 μmol de poliaminas), el contenido en poliaminas podría, teóricamente, tener un efecto bioactivo (29). También se ha visto que la espermidina y la espermina presentes en la leche materna disminuyen la permeabilidad intestinal a macromoléculas y quizás sea este el mecanismo por el que reducen la frecuencia de alergias alimentarias en niños (18).
CONCLUSIONES
En vista de todo lo dicho anteriormente, sería recomendable hacer algún estudio con el fin de establecer cual es el rango en el que oscila el contenido en poliaminas en las fórmulas infantiles a día de hoy, ya que los datos que están disponibles en la bibliografía son de hace más de quince años, pudiendo haber cambiado desde entonces las técnicas de elaboración de dichas fórmulas y, por tanto, el contenido en poliaminas.
Aunque no se han establecido recomendaciones en cuanto al nivel de poliaminas en la alimentación infantil, es importante conseguir que el aporte de poliaminas en las fórmulas infantiles sea lo más parecido posible al de la leche materna, ya que, como hemos visto, se han demostrado tanto los beneficios que tiene un adecuado aporte como los inconvenientes que podrían derivar de su carencia.
En este sentido, y en relación a la variabilidad encontrada en cuanto al contenido en poliaminas de la leche materna, sería recomendable hacer un estudio dentro de la población española, para poder comparar con los resultados que sí han sido publicados en otros países.
AGRADECIMIENTOS
A la Fundación Séneca, Agencia Regional de Ciencia y Tecnología, por la beca concedida a Carlos Gómez Gallego para la realización de la tesis doctoral en la que queda recogida la presente revisión bibliográfica.
REFERENCIAS
1. Morrys D and Harada JJ. The participation of polyamines in the proliferation of bacterial and animal cells. Polyamines in Biomedical Research, Gaugas, J. M. (ed.), Wiley, New York, 1988, p 1-16. [ Links ]
2. Farriol M, Segovia-Silvestre T, Venereo Y, Orta X. Importancia de las poliaminas: revisión de la literatura. Nutr Hosp 1999; 14: 101-113. [ Links ]
3. Bardocz S. Polyamines in food and their consequences for food quality and human health. Trends Food Sci Tech 1995; 6l, 341- 346. [ Links ]
4. Reynoso Orozco R, Delgado Saucedo JL, Puebla Pérez AM, Calvo Méndez C, Arteaga Nieto P, Santerre A. Biología celular y molecular de las poliaminas: monografía de los conocimientos básicos. Scientia-CUCBA 2005; 7 (2): 161-170. [ Links ]
5. Larqué E, Sabater-Molina M and Zamora S. Biological significance of dietary polyamines. Nutrition 2007; 23: 87–95. [ Links ]
6. Wallace HM, Fraser AV and Hughes A. A perspective of polyamine metabolism. Biochem J 2003; 376: 1-14. [ Links ]
7. Löser C. Polyamines in human and animal milk. Br J Nut 2000; 84: (Suppl.1): S55-S58. [ Links ]
8. Kalac P, Krausova P. A review of dietary polyamines: Formation, implications for growth and health and occurrence in foods. Food Chem 2005; 90:219–230. [ Links ]
9. Farriol M, Venereo Y, Orta X, Company C, Gómez P, Delgado G and Rodríguez R. Ingesta de antioxidantes y poliaminas en pacientes con quemaduras graves. Nutr Hosp 2004; 19: 300- 304. [ Links ]
10. Dandrifosse G, Peulen O, El Khefif N, Deloyer P, Dandrifosse AC and Grandfils C. Are milk polyamines preventive agents against food allergy. Proc Nutr Soc 2000; 59:81–6. [ Links ]
11. Moinard C, Cynober L, De Bandt JP. Polyamines: metabolism and implications in human diseases. Clinical Nut 2005; 24: 184-197. [ Links ]
12. Gugliucci A. and Menini T. The polyamines spermine and spermidine protect proteins from structural and functional damage by AGE precursors: a new role for old molecules? Life Sci 2003;72/23:2603–2616. [ Links ]
13. Nishimura K, Murozumi K, Shirahata A, Park MH, Kashiwagi K and Igarashi K. Independent roles of eIF5A and polyamines in cell proliferation. Biochem J 2005; 385: 779-785. [ Links ]
14. Nishimura K, Shiina R, Kashiwagi K and Igarashi K. Decrease in polyamines with aging and their ingestion from food and drink. J Biochem 2006; 139: 81-90. [ Links ]
15. Coffino P. Regulation of cellular polyamines by antizyme. Nature Reviews, Mol Cell Biol 2001; 2: 188- 194. [ Links ]
16. Casal S, Oliveira MBPP and Ferreira MA. Determination of biogenic amines in coffee by an optimized liquid chromatographic method. J Liq Chromatogr & Rel Technol 2002; 25 (16): 2535- 2549. [ Links ]
17. Baró L, Jiménez J, Martínez-Férez A, Boza JJ. Bioactive compounds derived from human milk. Ars Pharmaceutic 2001; 42:1; 21-38. [ Links ]
18. Motyl T, Ptoszaj T, Wojtasik A, Kukulska W and Podgurniak M. Polyamines in cows and sows milk. Comp Biochem Physiol. 1995; 111: 427-133. [ Links ]
19. Pollack PF, Koldousky O, Nishioka F. Polyamines in human and rat milk and in infant formulas. Am J Clin Nutr 1992; 56:371-375. [ Links ]
20. Buts JP, Dekeyser N, Deraedemaeker L, Collete E, Sokal EM. Polyamine profiles in human-milk, infant artificial formulas, and semielemental diets. J Ped Gast Nut 1995; 21 (1): 44-49. [ Links ]
21. Lavizzari T, Veciana-Nogues MT, Bover-Cid S, Marine-Font A, Vidal-Carou MC. Improved method for the determination of biogenic amines and polyamines in vegetable products by ion-pair high-performance liquid chromatography. J Chromatogr 2006; 1129: 67–72. [ Links ]
22. Gaboriau F, Vaultier M, Moulinoux JP, Delcros JG. Antioxidative properties of natural polyamines and dimethylsilane analogues. Redox Rep 2005; 1: 9-18. [ Links ]
23. Bachrach U. Polyamines and cancer: Minireview article. Amino Acids 2004; 26: 307-309. [ Links ]
24. Novella-Rodríguez S, Veciana-Nogués MT, Izquierdo-Pulido M and Vidal Carou MC. Distribution of Biogenic Amines and Polyamines in Cheese. J Food Sci 2003; 68: 750-755. [ Links ]
25. Soufleros EH, Bouloumpasi E, Zotou A, Loukou Z. Determination of biogenic amines in Greek wines by HPLC and ultraviolet detection after dansylation and examination of factors affecting their presence and concentration. Food Chem 2007; 101: 704–716. [ Links ]
26. Biol-Ngaragba MC and Louisot P. Regulation of the intestinal glycoprotein glycosylation during postnatal development: role of hormonal and nutritional factors. Biochimie 2003; 85, 331- 352. [ Links ]
27. Biol-Ngaragba MC, Greco S, George P, Hugueny I and Louisot P. Polyamine participation in the maturation of glycoprotein fucosylation, but not sialylation, in rat small intestine. Ped Res 2002; 51: 625- 634. [ Links ]
28. Romain N, Dandrifosse G, Jeuselte F, Forget G. Polyamine concentration in rat milk and food, human milk and infant formulae. Ped Res 1992; 32: 58-63. [ Links ]
29. Buts JP, De Keyser N, De Raedemaeker L. Saccharomyces boulardii enhances rat intestinal enzyme expression by endoluminal release of polyamines. Ped Res 1994; 36:522-527. [ Links ]

