











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkActa Botánica Venezuelica
versión impresa ISSN 0084-5906
Acta Bot. Venez. v.24 n.2 Caracas jun. 2001
CONTRIBUTIONS TO SOUTH AMERICAN CAESALPINIACEAE. VI. CAMPSIANDRA FELIPEANA (CAESALPINIEAE) A DWARF SWAMP-FOREST ENDEMIC OF THE LOWER ORINOCO WATERSHED
Basil STERGIOS D.
BioCentro, UNELLEZ, Mesa de Cavacas, Estado Portuguesa, Venezuela 3323 basilven@cantv.net
RESUMEN
Se describe e ilustra Campsiandra felipeana, una nueva especie de la cuenca baja del Río Cinaruco en el Estado Apure de Venezuela. Se encuentra principalmente en las cuencas de agua semi-negra del drenaje fluvial Cinaruco-Capanaparo de la llanura eólica, y muestra afinidades con las especies amazónicas del género. Esta especie está más relacionada con las especies C. emonensis, C. nutans y C. aymardii de los hábitats de agua negra o semi-negra de la región de la Guayana venezolana. Se discuten las relaciones entre estas especies y sus hábitats.
Palabras clave: agua negra, Campsiandra, chiga, Cinaruco, delta interno, Orinoco.
ABSTRACT
Campsiandra felipeana, a new species from the lower Cinaruco River drainage in the state of Apure, Venezuela is described and illustrated. It is primarily confined to the semiblack-water Cinaruco-Capanaparo watersheds of the aeolian llanura, and shows affinities to black-water Amazonian species of the genus. It is most closely related to C. emonensis, C. nutans and C. aymardii of the Venezuelan Guayana region of black-water or semiblack-water habitats. Relationships to these species and their habitats are discussed.
Key words: black-water, Campsiandra, chiga, Cinaruco, internal delta, Orinoco.
The Cinaruco River, which flows lazily from the western border of Venezuela with Colombia eastwards across the open plains or llanos of southern Apure state in east-central Venezuela, is characterised as a semi-black water (darkly tea-coloured), aeolian plains river habitat. This distinguishes it from most of the other major rivers in the region (PDVSA 1995). As one approaches the final drainage into the Orinoco in the east, the river expands into a permanently flooded internal delta, characterised by a labyrinth of extended branchings, filled with rather shallowly submerged sand-bars and bordered by low, saturated embankments or semiblack-water swamp-forests. Scattered within the river delta itself, are patches of spindly, low-treelet or shrubby emergent vegetation, which can be readily sampled by meandering from one patch to another in a small, paddled dug-out. Given the previously described particulars, the Cinaruco River, especially as it approaches the Orinoco, has been shown to harbour a number of plant genera and species most typically found in the Venezuelan Amazonian black-water habitats (Schargel & Aymard 1993; Stergios 1997). Such flooded, open-water emergents as Malouetia pubescens (R. Gómez et al. 564; Davidse & González 15629) help to underscore this tendency, as regarding the genus Malouetia in general ("palo de boya").
Inhabiting, sometimes exclusively, many of these delta sand-bars is a unique, spindly-trunked, low-emergent treelet of the genus Campsiandra, hitherto unknown to science. This new species has coriaceous leaflets typical of vegetation found in black-water Amazonian habitats.
Campsiandra felipeana Stergios, sp. nov.
TIPO: VENEZUELA. APURE: rebalse del Río Cinaruco, entre su boca y el campamento CORPOVEN, January 1987, B. Stergios, P. Stergios & D. Taphorn 9682 (Holotype, PORT; Isotypes, BM, GUYN, K, MO, NY, TFAV, US, VEN. Fig. 1.
Arbor gracilis, parum emergens, arenicola et reparina, 4-6 m alta. Folia (7)9-foliata, foliolis coriaceis, oblongo-lanceolatis, 11-15 cm longis, 2.5-3 cm latis; supra lucidis, glabris vel fere glabris, excepta costa complanata; subtus adpresso-puberulentis, copiose rubro-ferrugineo indutis; venis utrinque elevato-reticulatis, praesertim subtus. Sepala dense ferrugineo-puberulenta, fascia angusta, saturate rubra nuda secus apicalem marginem donata; calycis tubo sparse puberulento, cyathiformi; pedicello 0.5-1 mm sub calycis tubo articulato.
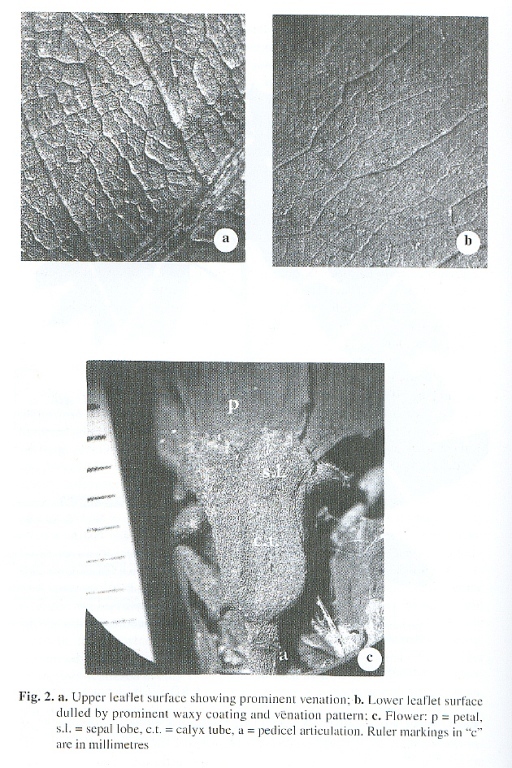
Spindly, low-emergent, tree, 4-6 m tall. Branchlets glabrous, glebulaterugosulose with circumferentially distributed, longitudinal, slightly darkened (in dried specimen), rounded ridges. Leaves imparipinnate, (7)9-foliate, 16-18 cm long; petioles briefly winged, 4-4.5 cm to the first pair of leaflets, canaliculate dorsally for 2-2.5 cm, then abruptly revolute to 1-1.5 cm; densely and maculately adpressed ferrugineous-puberulent amidst a copious, superficial, somewhat glaucous, waxy coating; rachis somewhat to obviously angular, semisulcate distally nearing each union of leaflets; petiolules terete, crassulate, circumferentially rugose, 2.5-3 mm long, 1.5 mm wide, ferrugineous-puberulent, sometimes with annularly transverse splitting in dried specimens. Leaflets opposite, most nearly perpendicular to the rachis, coriaceous, oblong-lanceolate, 11-11.5 cm long, 2.5-3 cm wide, the more distal leaflets slightly falcate except the terminal one, apex rounded-mucronate; upper surface lustrous, glabrous, but adpressed-puberulent along a flattened, non-raised, dark-red, glaucous-waxy midvein; lower surface with a thickened, reddish-ferrugineous waxy coating, adpressed puberulent with bubalinous-brunneolate hairs, both of these latter characters scattered evenly over blade surface as well as along a triquetrousraised, dark-red midvein with irregular blotches of waxy deposits; adaxial venation notably reticulately raised and readily visible to the quinary veinlets (Fig. 2-a), all reddish-coloured; abaxial venation also raised, reticulate, but much less prominent (Fig. 2-b), quarternary and quinary veinlets hardly visible. Inflorescence a terminal, somewhat loose, panicle of racemes 12-15 cm long, 8-10 cm wide, finely but densely and evenly ferrugineous-puberulent along all branches; branched racemes ascending, straight or slightly incurved, the lower ones 6-7 cm long, shortening to 2.5-3 cm long at apex; pedicels straight to recurving, minutely adpressed-puberulent, 1.5-1.8 cm long, arising from the distal third of the raceme branch, setaceous, early-caducous below the distal third to about mid-point on the fertile branch, articulating at about 5 mm below the base of the calyx tube. Flowers rather small but showy, 1-1.5 cm long at anthesis, more densely packed towards the distal portions of the raceme branchlets; bracts early-caducous, cymbiform, minute, 1 mm long, 0.5 mm wide at base, straminousminute- velutinous without, nitid, lustrous, glabrous and dark red-black within; calyx tube cyathiform, scarcely adpressed-puberulent to nearly glabrous (Fig. 2-c), 5 mm long, 4 mm wide at base of sepal lobes; sepals widely deltoid-obtuse, auriculate-imbricate at the basal corners, 2 mm long, 3 mm wide at the base, densely adpressed-ferrugineous-puberulent, but with a narrow naked strip around the outer margin, 0.25 mm wide, very dark red-coloured when dried; petals usually white, sometimes with a faint rose-coloured tinge, 10 mm long, 4 mm wide, slightly cucullate at margins, borders ciliate 3/4 -way to the base to nearly at the base; stamens 15, filaments filamentous, tortuous, 3.5-4 cm long, dark red-coloured, glabrous; anthers copiously ferrugineous-lanulose-pilose below, only sparsely pilosulose above, 1.5 mm long, 1.2 mm wide; ovary flattened, slightly falcate, 4 mm long, 1 mm wide, stipe 4 mm long at anthesis, style 2.5 cm long, glabrous, dark red-coloured, stigma truncate, slightly infundibuliform, ovules 10, obloid. Pods lustrous, flattened, slightly falcate, 18-22 cm long, 4.5-5 cm wide. Seeds compressed-ovoid to compressed-obloid, 4 cm x 5.5 cm; testa chartaceous, fragile, extending from seed-body into a circumnexate, thickened, spongy wing, 5-7 mm wide.

Distribution and Ecology. Restricted to semi-black water, llanuran or eolic waterways of southern Apure State in Venezuela, most notably the riveredge swamp-forests and exposed sand bars of the lower Cinaruco River and less frequently in the nearby watershed of the La Pica River (Caño La Pica) and the Cunivichito River, the latter two southern-bank tributaries of the Capanaparo River. As with the more common Campsiandra implexicaulis (Stergios 1996), this species flowers from December-January, setting fruit from March-April, or thereabouts.
Paratypes. VENEZUELA: APURE: Río Cinaruco, selva de rebalse, margen derecha de la boca, diciembre 1971, B. Trujillo 11742 (MY); Río Cinaruco, rebalse ribereño entre su boca y campamento de CORPOVEN, dicembre 1986-enero 1987, B. Stergios et al. 9681 (PORT,VEN), 9683 (K,PORT,VEN), 9684 (BM, K, MY, NY, PORT, US, VEN); Río Cinaruco, entre el campamento de CORPOVEN y boca del Juriepe, diciembre 1986-enero 1987, B. Stergios et al. 9688 (BM, PORT, US, VEN), 9698 (NY, PORT, US); Caño Cunivichito, trocha La Soledad-Capanaparo, diciembre 1986-enero 1987, B. Stergios et al. 9752 (BM, GUYN, K,MY, MYF, NY, PORT, TFAV, US, VEN); Caño La Pica, 1,5 km aguas arriba del paso chalana, costilla sur, junio 1994, B. Stergios 16134 (K,PORT, NY).
Relationships. Outstanding amongst the most unique features of Campsiandra felipeana is the spindly, single-trunked low-growth, emergent aquatic or semi-acuatic habit, coupled with a particular, open sand-bar or low swamp-forest habitat. Although other species of Campsiandra known to exist in the region (C. implexicaulis, C. angustifolia, C. taphornii; Stergios 1993, 1996) typically are found in similar habitats, the trees are much larger and more robust, inhabit only seasonally inundated swamp-forests, or are restricted to gallery forest embankments. Nearly all these other species have multiple trunks. Campsiandra felipeana belongs to the adpressed-pubescent group of the genus (see Stergios 1996), and can be most closely allied to the C. emonensis, C. nutans, C. aymardii subgroup, having the proliferate, dull waxy coating on the underside of the leaflets, a more or less rounded-mucronate leaflet apex, and a distinctly trigonous lower midvein with a narrowing, raised ridge. Campsiandra felipeana can be distinguished from all three by the notably coriaceous leaflets (a characteristic primarily prevalent in the other known primarily black-water, erect-pubescent group of the genus), the reddish-ferrugineous aspect of the waxy coating, and the distinctly dark-red colouring of the midvein on both leaflet surfaces, and the notable prevalence of a waxy residue on both upper and lower midveins. The dark-red, narrow naked band along the apex of the sepals is narrow, but sharply defined from the rather dense, ferrugineous sepal pubescence, the pedicels generally erect, not drooping, and the articulation closer to (but not at) the base of the calyx tube.
The previously known spindly-trunked growth habit in Campsiandra pertained only to the Amazonian black-water C. macrocarpa, which is confined primarily to the black-water swamp-forests of the Casiquiare-Rio Negro watersheds of southern Venezuelan Amazonas (Cowan 1958; Stergios 1996). This habit is generally typical of many other black-water, swamp-forest species, together with the coriaceous leaves (or leaflets), low growth habit, and high tolerance of consistent inundation. However, these trees may reach much larger diameters and be as tall as 12-15 m, a trait not found with C. felipeana. Also, the erectpubescent character has shown to be a much stronger and consistent tie to the black-water Campsiandra group than trunk and texture characters. Nevertheless, it is indeed notable that these more typical black-water Amazonian traits can also be present in more northern Venezuelan black to semiblack-water habitats.
Campsiandra felipeana is dedicated to an outstanding Venezuelan researcher, educator, colleague and founding President of the UNELLEZ university in the Venezuelan llanos, Dr. Felipe Gómez Alvarez (1923-2001), who pioneered interest in the ethnobotany of Campsiandra amongst the Indian groups in southern Apure State of Venezuela (see Stergios 1993), and its possible potential as a viable cultivar.
ACKNOWLEDGEMENTS
I am indebted to CONICIT Research Grant S1-1806 for providing the funds for the initial field-work phases of the study. I am grateful to Laurence J. Dorr (US) for his review of the manuscript, and to Bruno Manara (VEN) for his expertise with the Latin diagnosis.
BIBLIOGRAPHY
1.- Cowan, R. 1958. Campsiandra macrocarpa. In: Maguire, B. et al. Botany of the Guayana Highland-Part III. Mem. New York Bot. Gard. 10(1): 1-166. [ Links ]
2.- Petróleos de Venezuela, S.A. (PDVSA). 1955. Imagen de Venezuela, una visión espacial. PDVSA, Caracas. [ Links ]
3.- Schargel, R. & G. Aymard. 1993. Observaciones sobre suelos y vegetación en la llanura eólica limosa situada entre los ríos Capanaparo y Riecito, Estado Apure,Venezuela. BioLlania 9: 110-147. [ Links ]
4.- Stergios, B. 1993. La etnobotánica del árbol "chiga" (Campsiandra, Caesalpiniaceae) en la región llanera de la cuenca del medio río Orinoco en el suroeste de Venezuela. BioLlania 9: 71-90. [ Links ]
5.- Stergios, B. 1996. Contributions to South American Caesalpiniaceae. II. A taxonomic update of Campsiandra (Caesalpinieae). Novon 6: 434-459. [ Links ]
6.- Stergios, B. 1997. Relación biodiversidad versus fisiografía y suelos en bosques de galería de los llanos occidentales. Sección Vegetación. Informe final del proyecto de investigación Convenio UNELLEZ-Fundación Polar. Guanare,Venezuela. [ Links ]
