











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkActa Botánica Venezuelica
versión impresa ISSN 0084-5906
Acta Bot. Venez. v.28 n.1 Caracas ene. 2005
Patrón Fenológico de una población de Gelidium Serrulatum J. Agardh (Rhodophyta, Gelidiales) en la localidad de Taguao, Estado Vargas, Venezuela
Sonia ARDITO1 y Santiago GÓMEZ2
1Fundación Instituto Botánico de Venezuela. Apartado 2156. Caracas 1010-A. e-mail: ardito@cantv.net.
2
Centro de Botánica Tropical, Instituto de Biología Experimental, Facultad de Ciencias. Universidad Central de Venezuela. e-mail: sagomez@strix.ciens.ucv.veRESUMEN
Se aporta información acerca de la fenología reproductiva de la especie Gelidium serrulatum J.Agardh, un alga roja de importancia económica presente en las costas venezolanas. En la población estudiada se encuentran presentes, durante todo el año, las diferentes fases que conforman el ciclo de vida de esta especie: tetrasporófitos, gametófitos femeninos y masculinos, siendo los talos tetraspóricos más abundantes que los gametofíticos, lo cual implica un desbalance en la proporción 1:1 esperada entre ellos; las proporciones entre los gametófitos (femeninos y masculinos) son similares. Esto indica que la reproducción sexual es un evento que debe ocurrir en la población estudiada, sin embargo el desbalance observado entre fases tetraspóricas y gametofíticas sugiere que mecanismos adicionales a la reproducción sexual pudieran estar interactuando en el mantenimiento de la población, uno de estos podría ser la baja viabilidad reportada para las tetrásporas de esta especie.
Palabras clave: Fenología, gametófito, Gelidiales, Gelidium, tetrasporófito
Phenology pattern of a population of Gelidium serrulatum J. Agardh (Rhodophyta, Gelidiales), Vargas State, Venezuela
ABSTRACT
Information on the reproductive phenology of Gelidium serrulatum J.Agardh, a red algal species with economical importance is given. In the studied population, during the whole year, the different phases of the life cycle of this species are presented: tetrasporophytes, male and female gametophytes, being tetrasporophytes more abundant than gametophytes, which implies an unbalance in the proportion 1:1 between them; on the other hand, the proportions among male and female gametophytes are similar. This indicates that the sexual reproduction is a probable event in the studied population; however, the unbalance observed between tetraspoprophytes and gametophytes suggest that additional mechanisms of reproduction could be interacting in the studied populations maintenance, one of these may be the reported low viability for tetraspores in this species.
Key words: Gametophyte, Gelidiales, Gelidium, phenology, tetrasporophyte
INTRODUCCIÓN
Uno de los aspectos más interesantes de las especies del género Gelidium y en general de todas las especies pertenecientes al orden Gelidiales, es el ciclo reproductivo, el cual es como el que ocurre en el género Polysiphonia caracterizado por presentar una alternancia de fases: una gametofítica haploide y dioica, una tetrasporofítica diploide y una tercera fase, la carposporofítica diploide que se desarrolla sobre el gametófito femenino. De acuerdo con este ciclo de vida se esperaría una alternancia de fases haplodiplónticas, de forma tal que la proporción de gametófitos y terasporófitos quedara equilibrada (1:1) (Santelices 1988; Salinas 1991; Melo & Neushul 1993); sin embargo, diversos estudios realizados en distintas latitudes señalan una marcada desproporción entre fases, siendo la fase tetrasporofítica (diploide) más frecuente en varios órdenes de magnitud que la gametofítica (haploide), e incluso esta última puede permanecer ausente en las poblaciones naturales (Santelices 1988a; Melo & Neushul 1993; Santos & Duarte 1996; Rico & Guiry 1997; Rueness & Frediksen 1998).
Estas desviaciones del patrón reproductivo esperado se han tratado de explicar en función de distintas hipótesis, tales como: 1- La influencia de los factores ambientales sobre el desarrollo y crecimiento de las distintas fases, 2- Posibles anomalías en los procesos reproductivos, 3- Mecanismos de propagación vegetativa eficientes; todas estas teorías se basan en una mayor fecundidad o supervivencia de una fase con respecto a otra (Santelices 1990b; Maggs & Rico 1991; Salinas 1991b; Juanes & Puente 1993; Rico & Guiry 1997; Rueness & Frediksen 1998); no obstante, el aporte que tenga cada uno de estos aspectos sobre el desarrollo de una determinada fase parece depender de la especie estudiada y del hábitat donde ésta se desarrolla.
En este trabajo se realiza un estudio acerca del patrón fenológico de la especie Gelidium serrulatum J.Agardh, determinando para ello la frecuencia de aparición y proporción de las distintas fases del ciclo de vida; además, se discute acerca de las distintas hipótesis propuestas que podrían estar afectando a la población estudiada.
ÁREA DE ESTUDIO
El área donde se realizaron las colecciones se encuentra al norte de Venezuela, en el estado Vargas, en la localidad de Taguao, a 10° 34 45" Lat. N y 67° 06 10" Long. O (Fig. 1) y consiste de plataformas coralinas en las cuales se distinguen tres niveles (superior, medio e inferior) correspondientes a la zona intermareal (Ardito et al. 1995). La población de Gelidium serrulatum se ubica en el borde de la plataforma, es decir, en la zona intermareal inferior, donde el grado de exposición al oleaje es fuerte.
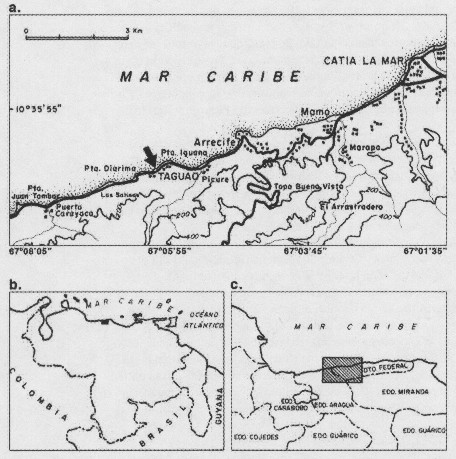
Ubicación geográfica del área de estudio. b. Ubicación relativa nacional. c. Ubicación relativa regional.Fig. 1. a.
MATERIALES Y MÉTODOS
Se realizaron salidas mensuales al área de estudio durante 13 meses (octubre 1998-octubre 1999) para colectar las algas, las cuales se colocaron en bolsas plásticas (16 x 15 cm) con cierre hermético, cada una de éstas representó una muestra; se colectó un total de 10 muestras por mes, en diferentes puntos de la plataforma, seleccionados al azar. Se transportaron a baja temperatura hasta el laboratorio, manteniéndose bajo estas condiciones hasta su procesamiento.
En el laboratorio se procedió a identificar y separar las fases reproductivas maduras presentes en cada muestra, siguiendo la metodología empleada por Huérfano (1998), la cual consiste en extender la muestra en una bandeja de vidrio colocada sobre una tabla cuadriculada en 100 recuadros numerados de área igual a 2 cm; se separaron 10 frondas seleccionadas al azar, mediante el uso de una tabla de números aleatorios, dando un total de 100 frondas por mes (se consideró fronda al eje erecto, incluyendo ramas y pínnulas de cualquier orden, de acuerdo con Akatsuka 1986). Cada fronda fue revisada al microscopio estereoscópico para determinar el tipo y el número de frondas correspondientes a cada fase, para así establecer tanto la frecuencia relativa como la proporción de las distintas fases.
La clasificación de las frondas se realizó en función de la presencia de las estructuras reproductivas: tetrasporangios en los tetrasporófitos, espermatangios en los gametófitos masculinos y cistocarpos con carposporófitos que crecen sobre los gametófitos femeninos y por lo tanto su presencia permite el reconocimiento de estos últimos; aquellas frondas donde no se observaron estructuras reproductivas se clasificaron como vegetativas.
La frecuencia relativa de cada fase fue determinada por su presencia y ausencia en las muestras mensuales y las proporciones fueron estimadas por el cociente: Número de frondas de cada fase/Total de frondas analizadas.
Se realizó la prueba estadística G con corrección de continuidad (Sokal & Rolhf 1986) para determinar si existían diferencias significativas entre las proporciones observadas y esperadas de los tetrasporófitos y gametófitos (de acuerdo al ciclo de vida de la especie en estudio). Para el análisis de las frecuencias de los gametófitos femeninos y masculinos, se calcularon las probabilidades binomiales con el objeto de determinar si existían diferencias significativas entre éstas; no se empleó la prueba de G para tal fin, debido a que ésta no se recomienda cuando el número poblacional es menor de 25 (Sokal & Rolhf 1986).
RESULTADOS
En la población estudiada de G. serrulatum las distintas fases que conforman el ciclo de vida estuvieron presentes durante todo el período de estudio, sin mostrar un patrón evidente de estacionalidad; sin embargo, se observó una clara dominancia de los tetrasporófitos sobre los gametófitos tanto femeninos como masculinos (Fig. 2).
Las fluctuaciones que presentaron cada una de las fases en el patrón de frecuencias de aparición mensual indican que los tetrasporófitos y estadios vegetativos son los más frecuentes en la población, seguidos por los gametófitos femeninos y por último los gametófitos masculinos (Fig. 2). La frecuencia de aparición de los tetrasporófitos varió entre 32 y 68%, presentando un máximo en el mes de octubre (1999) y un mínimo en el mes de abril; en el caso de los talos vegetativos la frecuencia fluctuó entre 11 y 48%, registrando el valor máximo en el mes de febrero y su mínimo en los meses de octubre y noviembre (1998). Los gametófitos femeninos presentaron un descenso en las frecuencias de aparición, a principios del estudio, durante los meses de octubre a marzo (de 19 a 1%), para luego fluctuar entre 9 y 17%, en el caso de los masculinos también se observó este descenso en sus frecuencias de aparición, pero fue de octubre a enero (de 14 a 2%), oscilando posteriormente entre 5 y 14% (Fig. 2).
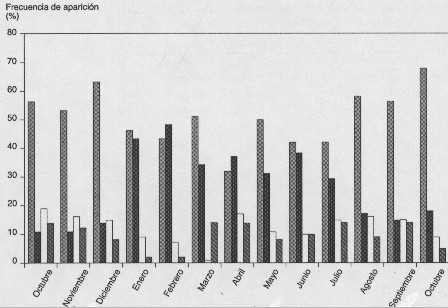
Tetrasporófito Vegetativo Gametófito femenino Gametófito masculino
Fig. 2. Distribución de frecuencias de aparición mensual de cada una de las fases de vida de G. serrulatum.
Las proporciones mensuales observadas de cada una de las fases señalaron un claro desbalance en la proporción 1:1 esperada entre los tetrasporófitos y los gametófitos, lo cual se ratificó con las pruebas de bondad de ajuste (pruebas de G) (Tabla 1). Los resultados indicaron que existen diferencias significativas entre las proporciones esperadas y observadas de los tetrasporófitos y gametófitos a lo largo del año, a excepción de los meses de abril y julio. Con respecto a las proporciones observadas y esperadas (1:1) entre los gametófitos femeninos y masculinos el cálculo de las probabilidades binomiales mostró la dominancia de los gametófitos femeninos únicamente en el mes de enero y de los gametófitos masculinos en el mes de marzo (p<0,05), en el resto de los meses la proporción entre ambos fue la esperada.
Tabla 1. Resultados de las pruebas de G realizadas para determinar diferencias entre las proporciones observadas y esperadas de los tetrasporófitos y gametófitos por mes y por año.

DISCUSIÓN
Los resultados de este estudio indican que tanto las fases gametofíticas (masculinas y femeninas) y tetrasporofíticas que conforman el ciclo de vida de Gelidium serrulatum están presentes en la población estudiada y que se distribuyen de forma desigual en el tiempo.
Patrones similares de abundancia de tetrasporófitos vs. gametófitos han sido observados para otras especies del género Gelidium, donde la escasez de talos sexuales es común (Montalva & Santelices 1981; Stewart & Norris 1981; Akatsuka 1986; Anderson et al. 1991; Salinas 1991a, b; Melo & Neushul 1993; Sosa et al. 1993). Resultados similares fueron señalados para poblaciones de Gelidium serrulatum en el Oriente del país (Loayza 1994). La proporción de talos femeninos y masculinos ha sido menos estudiada y pocos trabajos refieren la frecuencia de aparición de los talos masculinos (Hoyle 1978; Steward & Norris 1981; Santos & Duarte 1996); además, son escasos los estudios que establecen relaciones de significancia estadística entre las proporciones de los gametófitos, lo cual no permite establecer con certeza cuan alejadas pudieran estar las proporciones observadas de las esperadas.
La dominancia de los tetrasporófitos en la población estudiada indica un desbalance en la proporción 1:1 esperada entre éstos y los gametófitos; por otro lado, la similitud en las proporciones de los gametófitos (femeninos y masculinos) en la mayor parte del año, señalan que la reproducción sexual es un evento que debe ocurrir en la población, por lo que es posible que las diferencias observadas en el patrón de distribución temporal de las proporciones de cada una de las fases del ciclo de vida de G. serrulatum se deban a que otras estrategias reproductivas, además de la reproducción sexual, pudieran estar actuando en el mantenimiento de la población.
Las diferentes hipótesis propuestas para explicar las posibles razones del desbalance en el patrón esperado de abundancia entre los tetrasporófitos y gametófitos en las poblaciones naturales se basan principalmente en una mayor fecundidad o supervivencia de una fase con respecto a otra o a mecanismos de reproducción como la propagación vegetativa y/o apomeiosis (Akatsuka 1986; Santelices 1988a; Hawkes 1990; Anderson et al. 1991; Destombe et al. 1993; Salinas 1991a, b; Juanes & Puente 1993; Sosa et al. 1993; Huérfano 1998).
También se ha intentado explicar esta aparente anomalía en función de los límites de distribución geográfica de cada especie. Dixon (1965, citado por Santelices 1978), hipotetizó que este fenómeno puede ser explicado en términos de inhibición ambiental de la expresión del potencial reproductivo, suponiendo una igualdad de talos gametofíticos y tetraspóricos en el centro de distribución geográfica (condiciones óptimas de crecimiento), iniciándose un desbalance en las proporciones esperadas a medida que se acerca a sus límites de distribución, pudiendo llegar a inhibirse completamente la expresión reproductiva de ambos talos en los extremos de la distribución geográfica. Esta hipótesis se ha verificado en algunas especies de Gelidium y Pterocladia (Irving & Dixon 1977, citado por Akatsuka 1986; Santelices 1988a, 1990a, b). El desbalance encontrado en las proporciones esperadas de G. serrulatum no podría ser explicado totalmente por la hipótesis antes planteada, dado que esta especie es esencialmente tropical y sólo ha sido descrita para Trinidad, Colombia y Venezuela (Ganesan 1989), por lo que es posible que aún los límites de distribución geográfica no hayan sido completamente demarcados.
Akatsuka (1986) y Santelices (1988a, 1990a) han interpretado la hipótesis propuesta por Dixon (1965, citado por Santelices 1978) de la forma siguiente: una determinada fase del ciclo de vida puede ser incapaz de expresarse bajo ciertas condiciones ambientales no óptimas, por lo tanto, se esperaría que la abundancia relativa de la reproducción sexual o asexual presente patrones de cambio latitudinales, debido a que los factores ambientales varían con la latitud, así como estacional o localmente, por lo cual es probable que los cambios temporales de factores ambientales jueguen un papel importante en la distribución de fases encontrada en la población estudiada.
Una hipótesis alternativa plantea diferencias marcadas en la capacidad de propagación vegetativa entre los tetrasporófitos y gametófitos. Se ha demostrado que existe una capacidad de propagación diferencial entre fases isomórficas (gametófito femenino y tetrasporófito) del ciclo de vida de especies de Gelidium y Pterocladia, siendo ésta mucho menor o en ocasiones inexistentes en las frondas gametofíticas (Salinas 1991a, b; Juanes & Puente 1993; Huérfano 1998); de esta forma este mecanismo de reproducción asexual podría contribuir a una distribución desigual de estos estadios reproductivos con una clara dominancia de los tetrasporofitos.
En las algas rojas se han observado procesos de apomeiosis (Hawkes 1990), entendiéndose por ésta la producción de esporas diploides por falta de meiosis (Huérfano 1998). Estudios de laboratorio sobre ciclos de vida y cariotipos en Gelidiales (Dixon 1963; Santelices 1988a, 1990b) sugieren que probablemente la meiosis no ocurre en todos los esporangios de un tetrasporófito dado; de esta forma, las esporas resultantes podrían ser haploides y/o diploides, adicionándose estas últimas a las carpósporas diploides y contribuyendo así a la dominancia del tetrasporófito; sin embargo, Carter (1985, citado por Anderson et al. 1991) señala que esta anomalía es un fenómeno inusual en las especies del género Gelidium; particularmente estudios cariológicos realizados en Gelidium serrulatum de la isla de Margarita (Kapraun et al. 1993), no revelan falta de meiosis en los tetrasporangios que pudiera ayudar a explicar la distribución desigual observada.
La falta de viabilidad de las tetrásporas es otra de la hipótesis que intenta explicar el desbalance en las proporciones observadas entre fases tetraspóricas y gametofíticas (Salinas 1991b). Distintos estudios de cultivo señalan diferencias marcadas entre tetrásporas y carpósporas, sugiriendo, bien sea en términos de porcentajes de germinación o de reclutamiento de juveniles a partir de una u otra espora, una mayor viabilidad de las carpósporas (Maggs & Rico 1991; Santos & Duarte 1996; Rueness & Frediksen 1998). En el caso particular de la población estudiada de G. serrulatum, se realizaron investigaciones sobre viabilidad y germinación de ambos tipos de esporas, y estos revelaron un mayor porcentaje de germinación de carpósporas con relación a las tetrásporas, a pesar de que éstas presentaron una mayor densidad de descarga, la viabilidad de las mismas fue menor. Estos resultados sugieren que la pobre viabilidad de las tetrásporas podría ser una de las causas que contribuyen a la escasez de gametófitos en la población estudiada, sin embargo, otros factores pudieran estar interactuando y contribuyendo al desbalance observado en esta especie, dado que la germinación es un proceso indispensable para el desarrollo de una plántula, pero no garantiza su supervivencia en el tiempo (Ardito 2001).
Otra de las hipótesis que se han propuesto para explicar este desbalance entre fases isomórficas está relacionada con el nivel de ploidía de éstas. Algunos estudios en distintas especies de algas rojas señalan diferencias en las respuestas fisiológicas y ecológicas entre fases isomórficas, que facilitarían una mayor supervivencia de la fase diploide en los mismos ambientes (Santelices 1988a, 1991b; Destombe et al. 1993).
En la determinación de la presencia de las fases reproductivas, otro de los aspectos que se ha discutido es el efecto del tipo de muestreo empleado (Akatsuka 1986; Huérfano 1998). Algunos estudios han omitido las frondas inmaduras o masculinas, lo cual no permite una adecuada interpretación del patrón de distribución de frecuencias de cada fase; además, otra limitante en los estudios de distribución de frecuencias en las agarofitas a diferencia de las carragenofitas (Avila et al. 1997) es que no existe ninguna prueba para determinar si las frondas inmaduras corresponderán a la fase gametofítica o tetrasporofítica. En este estudio las frondas vegetativas o inmaduras tienden a incrementar cuando disminuyen los tetrasporofitos, y en menor grado cuando disminuyen los gametófitos, lo cual lleva a pensar que posiblemente la mayoría de las frondas inmaduras se desarrollarán en frondas tetrasporofíticas.
El planteamiento de todas las hipótesis anteriormente explicadas está indicando que el patrón de frecuencia de cada una de las fases del ciclo de vida de una determinada especie puede depender de muchos factores, lo que señala la complejidad de este fenómeno. De acuerdo con los resultados de esta investigación no es posible argumentar cuál hipótesis tendría más importancia en la población estudiada; sin embargo, los resultados sugieren mecanismos adicionales a la reproducción sexual en el mantenimiento de la población.
AGRADECIMIENTOS
Los autores de este trabajo agradecen al Consejo de Desarrollo Científico y Humanístico de la Universidad Central de Venezuela por el financiamiento parcial de esta investigación, a través del Proyecto No. PI- 03-33-4329-1999.
BIBLIOGRAFÍA
1. Akatsuka, I. 1986. Japanese Gelidiales (Rhodophyta), specially Gelidium. Oceanogr. Mar. Biol. Ann. Rev. 24: 171-263. [ Links ]
2. Anderson, R., R. Simons, N. Simons, N. Jarman & G. Levitt. 1991. Gelidium pristoides in South Africa. Hydrobiologia 221: 55-66. [ Links ]
3. Ardito, S., S. Gómez & B. Vera. 1995. Estudio sistemático de las macroalgas marinas bentónicas en la localidad de Taguao, Distrito Federal, Litoral Central, Venezuela. Acta. Bot. Venez. 18: 53-66. [ Links ]
4. Ardito, S. 2001. Estudio del potencial reproductivo de Gelidium serrulatum J. Agardh (Rhodophyta, Gelidiales) y de la calidad del agar extraído de las diferentes fases del ciclo de vida. Tesis Doctoral. Facultad de Ciencias. Universidad Central de Venezuela. Caracas, Venezuela. [ Links ]
5. Avila, M., M. Núñez, A. Candia & R. Norambuena. 1997. Patrones fenológicos reproductivos de una población de Gigartina skottsbergii (Gigartinaceae, Rhodophyta), en Ancud, Chile. Gayana Oceanol. 5: 21-32. [ Links ]
6. Destombe, Ch., J. Godin, M. Nocher, S. Richerd & M. Valero. 1993. Differences in responses between haploid and diploid isomorphic phases of Gracilaria verrucosa (Rhodophyta: Gigartinales) exposed to artificial environmental conditions. Hydrobiologia 260/261: 131-137. [ Links ]
7. Dixon, P.S. 1963. The Rhodophyta: some aspects of their biology. Oceanogr. Mar. Biol. Ann. Rev. 1: 177-196. [ Links ]
8. Ganesan, E. 1989. A catalog of benthic marine algae and seagrasses of Venezuela. Fondo Editorial CONICIT. Caracas. [ Links ]
9. Juanes, P. & A. Puente. 1993. Differential reattachment capacity of isomorphic life history phases of Gelidium sesquipedale. Hydrobiologia 260/261: 139-114. [ Links ]
10. Hawkes, M. 1990. Reproductive strategies. In: Biology of the red algae (Cole, K.M. & R.G. Sheath, eds.), pp 475-475. Cambridge University Press, Cambridge. [ Links ]
11. Hoyle, M. 1978. Agar studies in two Gracilaria species (G. Bursapastoris (Gmelin) Silva and G. coronopifolia J.Agardh) from Hawai. I. Yield and gel strength in the gametophyte and tetrasporophyte generations. Bot. Mar. 21: 343-345. [ Links ]
12. Huérfano, A. 1998. Aspectos sobre la biología poblacional, citogenética experimental y cultivo bajo condiciones controladas de la agarofita Pterocladia capillacea (Gelidiales, Rhodophyta) presente en la zona litoral central de Venezuela. Tesis Doctoral. Facultad de Ciencias. Universidad Central de Venezuela. Caracas, Venezuela. [ Links ]
13. Kapraun, D.F., J.A. Dutcher & D. Freshwater. 1993. Quantification and characterization of nuclear genomes in commercially red seaweeds: Gracilariales and Gelidiales. Hydrobiologia 260/261: 679-688. [ Links ]
14. Loayza, R. 1994. Aspectos taxonómicos, fases de desarrollo "in vitro", fenología y caracteres reológicos del agar de Gelidium serrulatum J. Agardh (Gelidiales, Rhodophyta). Tesis de Maestría. Facultad de Ciencias. Universidad de Oriente. Cumaná, Venezuela. [ Links ]
15. Maggs, C.A. & J.M. Rico. 1991. A kariological demonstration of meiosis in Gelidium latifolium (Gelidiaceae, Rhodophyta) from Ireland. Phycologia 30: 487-494. [ Links ]
16. Melo, R. & M. Neushul. 1993. Life history and reproductive potential of the agarophyte Gelidium robustum in California. Hydrobiologia 260/261: 223-229. [ Links ]
17. Montalva, S. & B. Santelices. 1981. Interspecific interference among species of Gelidium from Central Chile. J. Exp. Mar. Biol. Ecol. 53: 77-88. [ Links ]
18. Rico, J.M. & M. Guiry. 1997. Life history and reproduction of Gelidium maggsiae sp. nov. (Rhodophyta, Gelidiales) from Ireland. Eur. J. Phycol. 32: 267-277. [ Links ]
19. Rueness, J. & S. Frediksen. 1998. Intraspecific reproductive variation in Gelidium pusillum (Stackh.) Le Jol. (Gelidiales, Rhodophyta) from Europe. J. App. Phycol. 10: 253-260. [ Links ]
20. Salinas, J. 1991a. Spray system for re-attachment of Gelidium sesquipedale (Clem.) Born et Thur. (Gelidiales, Rhodophyta). Hydrobiologia 2-1: 107-117. [ Links ]
21. Salinas, J. 1991b. El proceso de refijación en Gelidium sesquipedale (Clem.) Born. et Thur. (Gelidiales, Rhodophyta). Bol. Inst. Esp. Oceanogr. 7(2): 3-58. [ Links ]
22. Santelices, B. 1978. Multiple interaction of factors in the distribution of some Hawaiian Gelidiales (Rhodophyta). Pacific Sci. 32: 119-147. [ Links ]
23. Santelices, B. 1988a. Synopsis of biological data on the seaweed genera Gelidium and Pterocladia (Rhodophya). FAO Fisheries Synopsis 145: 1-45. [ Links ]
24. Santelices, B. 1988b. Algas Marinas de Chile. Ediciones Universidad Católica de Chile. Santiago de Chile. [ Links ]
25. Santelices, B. 1990a. Patterns of reproduction, dispersal and recruitment in seaweeds. Oceanogr. Mar. Biol. Ann. Rev. 28: 177-276. [ Links ]
26. Santelices, B. 1990b. New and old problems in the taxonomy of the Gelidiales (Rhodophyta). Hydrobiologia 204/205: 125-135. [ Links ]
27. Santos, R. & P. Duarte. 1996. Fecundity, spore recruitment and size in Gelidium sesquipedale. Mar. Ecol. Prog. Ser. 119: 253-263. [ Links ]
28. Sokal, R. & F. Rohlf. 1986. Introducción a la Bioestadística. Editorial Reverté, S. A. Barcelona. [ Links ]
29. Sosa, P.A., M. Jiménez del Río & G. García-Reina. 1993. Physiological comparison between gamethophytes and tetrasporophytes de Gelidium canariensis (Gelidiaceae: Rhodophyta). Hydrobiologia 260/261: 445-449. [ Links ]
30. Stewart, J.G. & J.N. Norris. 1981. Gelidiaceae (Rhodophyta) from the northern Gulf of California, México. Phycologia 20: 273-284. [ Links ]
