











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkActa Botánica Venezuelica
versión impresa ISSN 0084-5906
Acta Bot. Venez. v.28 n.2 Caracas 2005
ANATOMÍA DE COMMELINACEAE PRESENTES EN UN CULTIVO DE PAPAS (SOLANUM TUBEROSUML.) EN MÉRIDA, VENEZUELA
Anatomy of Commelinaceae present in a culture of potatoes (Solanum tuberosum L.) in Merida, Venezuela
Rebeca LUQUE ARIAS1 y Javier ESTRADA SÁNCHEZ2
1-2
Centro Jardín Botánico, Facultad de Ciencias ULA, Mérida, Venezuela1 rebecal@ula.ve
2 jestr@ula.v
RESUMEN
Commelinaceae, después de Poaceae y Cyperaceae, es la familia de monocotiledóneas con mayor número de malezas de cultivo; entre ellas, algunas especies de Commelina y Tinantia son hospederas de ácaros, royas y virus, como Potato Y potyvirus que afecta a Solanum tuberosum. Se realizó un estudio de la anatomía de los órganos vegetativos de representantes de las especies Commelina obliquay Tinantia erecta que crecen en un cultivo de papas en los Andes de Mérida, para tratar de determinar la existencia de caracteres anatómicos comunes y diferenciales para las especies, así como cuales de estos favorecen su establecimiento en el cultivo y la existencia o no de caracteres que indiquen la presencia de posibles patógenos asociados a ellas. Se encontró que ambas especies poseen hojas anfiestomáticas, con epidermis de células altas, cutícula delgada, mesofilo compacto bien diferenciado, tallos con abundantes raíces adventicias, caracteres éstos que permiten el comportamiento de malezas. Además se notó un contenido aparentemente plasmodial en las células de la vaina vascular que podría estar asociado a la presencia de patógenos.
Palabras clave: Commelinaceae, Commelina obliqua, Tinantia erecta, Solanum tuberosum, Anatomía, Malezas de cultivo
ABSTRACT
Commelinaceae is, after Poaceae and Cyperaceae, the family of monocotiledons with the greatest number of weeds associated to cultures; among them, some species of Commelina and Tinantia are host for acarus, rust and virus, like the Potato Y potyvirus that affects Solanum tuberosum. In the Andes of Mérida, Commelina obliqua and Tinantia erecta grow like weeds in cultivation of potato. To try to determine the existence of common and differential anatomical characters among them, as well as which anatomical characters favor its establishment in the cultivation, and the existence or not of characters that indicate the presence of possible pathogenic agents associated to them, the anatomy of its vegetative organs was carried out. Both species have amphistomatic leaves, tall epidermical cells , thin cuticle; compact and well defined mesophyll; stem showing abundant adventitious roots; all of those being characters that permit the weed condition. In addition, the parenchyma cells of the leaves sheath seem to be filled with a plasmodium, suggesting infection by pathogens.
Key words: Commelinaceae, Commelina obliqua, Tinantia erecta, Solanum tuberosum, Anatomy,Weeds
INTRODUCCIÓN
Commelinaceae es una familia con alrededor de 40 géneros y 1.000 especies, de distribución principalmente pantropical con algunos representantes de zonas de climas templados; consiste de hierbas perennes comúnmente rizomatosas y suculentas aunque algunas especies de Tinantia Scheidw. y Commelina L. son consideradas anuales (Tomlinson 1969). Anatómicamente la familia presenta caracteres propios que hacen que se considere como un grupo natural y aislado, siendo difícil sugerir relaciones entre Commelinaceae y cualquier otra familia de monocotiledóneas. Después de Poaceae y Cyperaceae es considerada la familia de monocotiledóneas con el mayor número de malezas de cultivo; representantes de los géneros Commelina y Tinantia se han caracterizado como malezas ruderales de hoja ancha y levemente nocivas, siendo consideradas como organismos "r" (Alán et al. 1995) que ocupan estados tempranos de la sucesión. A pesar de ser plantas medianamente invasoras, algunas características las hacen nocivas a los cultivos. Así, Commelina diffusa Burm.f. es hospedera del ácaro Tetranychus urticae Koch que ataca cultivos de ajo, apio, frijol, cebolla, tomate, plantas ornamentales, etc. También esta especie es hospedera de enfermedades causadas por royas y virus. Tinantia erecta (Jacq.) Schlecht. es susceptible a numerosos virus entre los que se encuentra el Potato Y potyvirus que afecta a Solanum tuberosum L. (Brunt et al. 1996).
En los Andes de Mérida, representantes de los géneros Commelina y Tinantia crecen en terrenos limpios que son destinados a la siembra de cultivos de papas (Solanum tuberosum) como es el caso de una parcela en la Estación Experimental de Santa Rosa del Instituto de Investigaciones Agropecuarias de la Universidad de Los Andes (IIAP-ULA), Mérida.
El presente trabajo tiene como objetivo principal realizar un estudio de la anatomía de los órganos vegetativos de representantes de las especies Commelina obliquaVahl y Tinantia erecta (Jacq.) Schldl. que crecen en un cultivo de papas (Solanum tuberosum) para tratar de determinar la existencia de caracteres anatómicos comunes y diferenciales para las especies, así como caracteres anatómicos que favorecen su establecimiento en el cultivo y tratar de determinar la existencia o no de caracteres que indiquen la presencia de posibles agentes patógenos asociados a ellas.
MATERIALES Y MÉTODOS
En la Estación Experimental de Santa Rosa del Instituto de Investigaciones Agropecuarias de la Universidad de los Andes (IIA-ULA) se colectó material de la parte aérea así como de la subterránea de plantas adultas de Commelina oblicua Vahl (Estrada J. & R. Luque 1022) y Tinantia erecta (Jacq.) Schldl (Estrada, J. & R. Luque 1021).
VENEZUELA. MÉRIDA: Estación Experimental de Santa Rosa (IIAPULA) en un cultivo de papas (Solanum tuberosum L.). 1.920 m snm, 8°35Lat. N, 71°8 Long. O. 18-VII-2001; Estrada, J. & R. Luque 1021 y 1022 [MERC]. Las muestras están depositadas en el Herbario MERC.
El área en que se emplaza el cultivo pertenece a la zona de bosque húmedo altimontano, aunque se encuentra fuertemente intervenida por la actividad agropecuaria. El suelo alrededor de la estación está clasificado como Humitropepts típico, franco grueso con estructura franco-arenosa; con pH entre 6,45-5,60. Los promedios de temperaturas máximas y mínimas en el período de 1985-2000 son de 22,2°C y 12,2°C respectivamente, lo que supone un rango de variación de 10°C y una temperatura media anual de 17,1°C; la precipitación media anual se sitúa en torno a los 2.000 mm en un patrón bimodal con dos períodos de altas precipitaciones, uno de abril a junio y el otro de septiembre a noviembre (Jaimez et al. 2001).
Para el estudio anatómico el material se fijó en FAA según Johansen (1940). Del material fijado se prepararon cortes histológicos a mano alzada y con micrótomo. Los cortes a mano fueron aclarados en solución de cloro comercial al 10% por 1 min, lavados en agua, colocados en ácido acético 1% y teñidos con la doble tinción azul de astra-fucsina básica (Kraus et al. 1998). Para la obtención de los cortes con micrótomo se deshidrató el material en una serie butílica (Johansen 1940) con inclusión en parafina (Sass 1951) y posteriormente se cortó con micrótomo rotativo. Los cortes fueron teñidos con la doble tinción azul de astra-fucsina básica (Luque et al. 1996).
Se examinó y fotografió el material con un microscopio Zeiss Axioscop 20.
RESULTADOS
Hoja
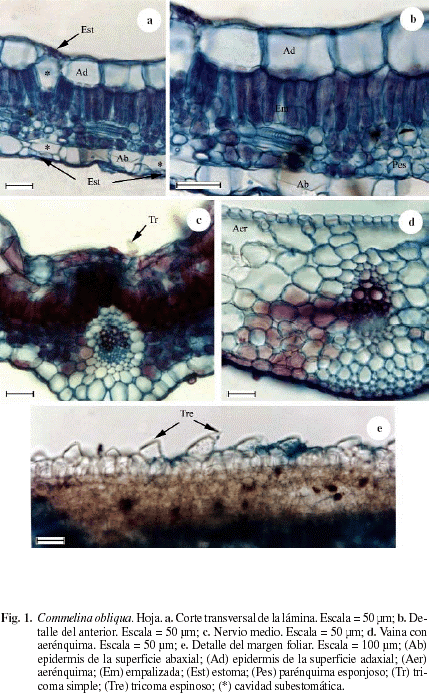
La figura 1a,b muestra un corte transversal de la hoja de Commelina oblicua en las que se observa la epidermis de las superficies adaxial y abaxial de una capa de células. Epidermis de la superficie adaxial formada por células cuadradas a rectangulares cuya altura es equivalente a 1/2 del espesor del mesofilo; en vista paradermal las células son poliédricas con las paredes delgadas, rectas a ligeramente onduladas (Fig. 3b), tricomas simples (Fig. 1c); cutícula y estratos cuniculares finos. Epidermis abaxial formada por células rectangulares cuya altura es equivalente de 1/4 a 1/3 del espesor del mesofilo; en vista paradermal las células son poliédricas con las paredes delgadas, rectas a ligeramente onduladas, distinguiéndose tricomas bicelulares espinosos y tricomas simples (Fig. 3a), en el borde de la lámina se observan tricomas bicelulares espinosos (Fig. 1e); ambas superficies son muy similares. Hoja anfiestomática, en ambas superficies estomas con distribución difusa y con cuatro células subsidiarias; cámara subestomática profunda (Fig. 1a). Mesofilo bifacial. Empalizada de una capa de células que ocupa aproximadamente 1/2 del espesor del mesofilo. Parénquima esponjoso compacto de tres a cuatro capas de células (Fig. 1b). Haces colaterales rodeados por una vaina parenquimática simple sin extensiones epidérmicas. Nervio medio poco diferenciado formado por un solo haz vascular (Fig. 1c). Vaina foliar con epidermis de una sola capa de células hacia ambas superficies, con estomas. Hacia la superficie adaxial se observa un aerénquima hipodérmico (Fig. 1d). Haces vasculares colaterales con una vaina vascular parenquimática simple.
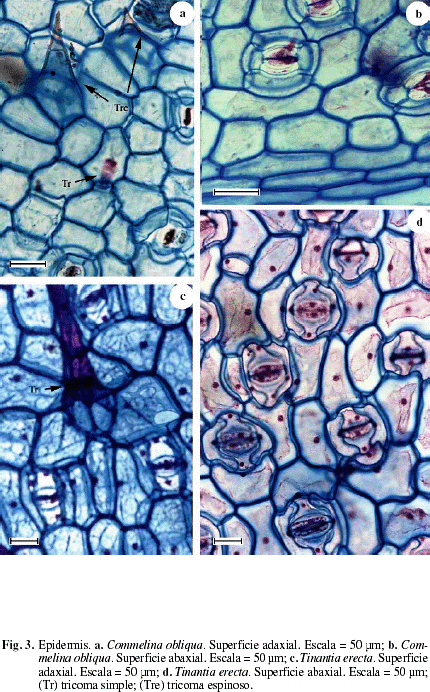
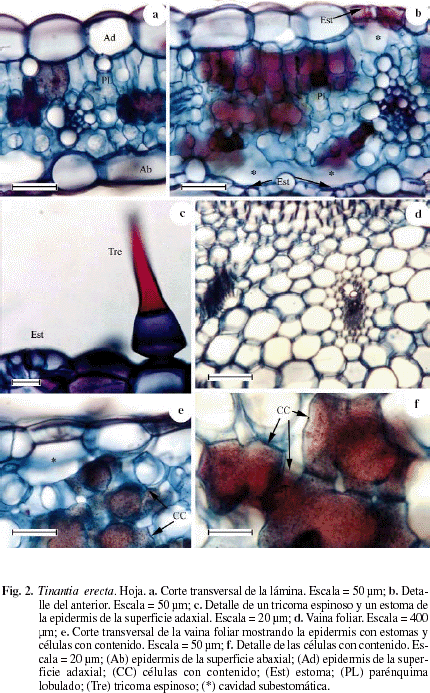
La Figura 2 representa el corte transversal de la hoja de Tinantia erecta en las que se observa la epidermis de las superficies adaxial y abaxial de una capa de células. Epidermis de la superficie adaxial formada por células cuadradas a rectangulares en corte transversal, con las paredes periclinales externas e internas un poco convexas (Fig. 2a, b) y cuya altura corresponde aproximadamente a 1/2 del espesor del mesofilo; en vista paradermal las células son poliédricas con las paredes ligeramente onduladas (Fig. 3c), tricomas simples y tricomas bicelulares espinosos (Fig. 2c). Epidermis de la superficie abaxial formada por células cuadradas a rectangulares en corte transversal y cuya altura corresponde a aproximadamente 1/3 del espesor del mesofilo; en vista paradermal las células son poliédricas con las paredes onduladas (Fig. 3d). Hojas anfiestomáticas con una mayor densidad de estomas en la superficie abaxial. Estomas de la superficie adaxial rodeados de 4 células subsidiarias (Fig. 3c) y los de la superficie abaxial rodeados por 6 células subsidiarias (Fig. 3d). Cavidad subestomática amplia (Fig. 2b). Mesofilo bifacial. Empalizada de 1-2 capas de células parenquimáticas lobuladas en forma de H (Fig. 2a, b), se observan células que se tiñen más intensamente posiblemente por la presencia de sustancias fenólicas. Parénquima esponjoso de 2-3 capas de células lobuladas. Haces vasculares colaterales rodeados por una vaina parenquimática simple, en el haz vascular se observa que la vaina vascular se prolonga hasta el límite de la cámara subestomática adaxial y abaxial (Fig. 2b). Aunque no se encuentra representado, el nervio medio es poco diferenciado y está formado por un solo haz vascular. Vaina foliar (Fig. 2d-f) con epidermis de una capa de células con estomas, con la cavidad subestomática amplia (Fig. 2e), en su parte central parenquimática con los haces colaterales; hacia la periferia se observan células lobuladas y en el interior de algunas de ellas un contenido denso que podrían corresponder a plasmodios (Fig. 2e, f).
Tallo
En la sección transversal del tallo de Commelina obliqua (Fig. 4a, b) se observa la epidermis de una capa de células más o menos cuadradas a elípticas con tricomas simples y glandulosos (Fig. 4d) y estomas (Fig. 4b). La corteza está formada por una región externa de 1-2 capas de células colenquimáticas, seguida por 6-8 capas de células parenquimáticas. El límite interno de la corteza está formado por una endodermis de células hialinas grandes (Fig. 4a, b). Haces vasculares periféricos (Fig. 4a, b) y perimedulares (Fig. 4e) con una vaina vascular esclerenquimática simple. En la Figura 4a se observa un haz vascular periférico con dos grandes vasos del metaxilema y el floema primario. La Figura 4c representa el corte longitudinal del tallo mostrando elementos de vaso anillados y espiralados.

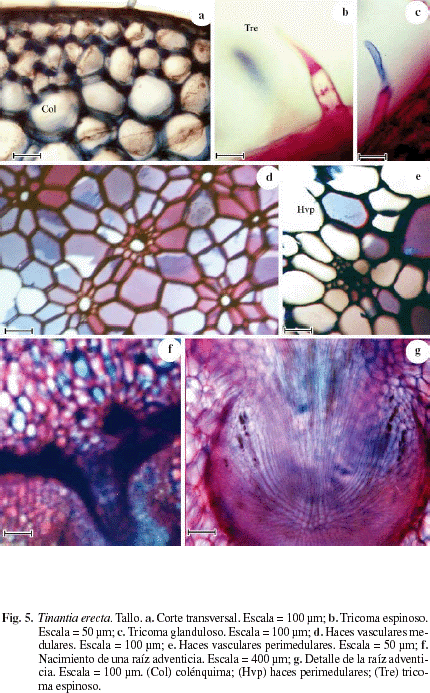
La Figura 5 representa el corte transversal del tallo de Tinantia erecta donde se observa una epidermis de una capa de células de paredes gruesas, con estomas y tricomas espinosos (Fig. 5b) y glandulosos (Fig. 5c). La corteza está formada por una hipodermis colenquimática de 3-4 capas de células (Fig. 5a), corteza parenquimática muy amplia. Haces vasculares medulares (Fig. 5d) y perimedulares (Fig. 5e). Se observan células con rafidios. Las figuras 5f, g representan la salida de una raíz adventicia.
Raíz
Las Figuras 6a, c representan la sección transversal de la raíz de Commelina obliqua mostrando la epidermis de células pequeñas, persistentes, internamente se diferencia la exodermis de 1-2 capas de células con paredes engrosadas, más grandes las externas (Fig. 6a); la corteza parenquimática es amplia (Fig. 6a) e internamente se diferencia la endodermis con bandas de Caspary (Fig. 6c); aunque no se encuentra representado, cuando la raíz alcanza un mayor desarrollo, las células de la endodermis pueden presentar engrosamientos en forma de U en sus paredes. El cilindro vascular está formado por un periciclo de células parenquimáticas (Fig. 6c), una estela que puede variar en el número de polos de protoxilema; en la Figura 6c se representa una poliarca con grandes vasos del metaxilema y los polos del floema primario alternando. La Figura 6b muestra la salida de una raíz lateral.
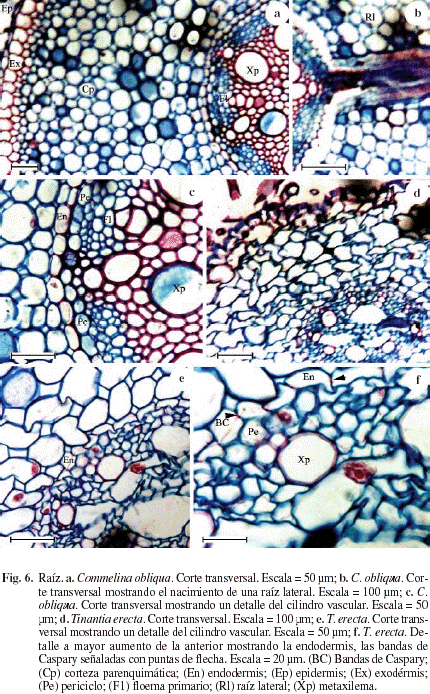
Las Figuras 6d, f representan la sección transversal de la raíz de Tinantia erecta mostrando la epidermis de una capa de células persistente con pelos radicales (Fig. 6d); internamente se diferencia la exodermis con células de paredes algo engrosadas, corteza parenquimática amplia e internamente diferenciada la endodermis (Fig. 6e, f) con bandas de Caspary (Fig. 6f); aunque no se encuentra representado, cuando la raíz alcanza un mayor desarrollo, las células de la endodermos pueden presentar engrosamientos en forma de U en sus paredes. El cilindro vascular está formado por un periciclo de células parenquimáticas (Fig. 6f), una estela poliarca donde se observan grandes vasos del metaxilema y los polos del floema primario alternando.
DISCUSIÓN
Las plantas estudiadas son herbáceas con los ejes construidos en nudos y entrenudos. Commelina obliqua presenta tallos decumbentes con entrenudos largos, mientras que Tinantia erecta presenta largos tallos erectos. Para Commelinaceae los representantes más especializados son aquellos que poseen una dorsiventralidad fija. El reemplazo de la simetría radial por la dorsiventral parece haber tenido un origen filogenético pero también puede ocurrir ontogenéticamente (Clark en Tomlinson 1969).
Anatómicamente los órganos vegetativos de las especies estudiadas presentan ciertos caracteres comunes:
1. Hojas de consistencia membranácea, dorsiventrales; cutícula delgada, epidermis de una capa de células en ambas superficies, formadas por células altas, carácter éste considerado como una adaptación a los ambientes abiertos con alta radiación solar; según Solereder & Meyer (en Tomlinson 1969), la epidermis de células altas grandes puede funcionar como un tejido reservante de agua, sin embargo, en este trabajo se consideró que estas células epidérmicas altas podrían estar funcionando como un filtro para el exceso de radiación.
2. Hojas anfiestomáticas, carácter que probablemente puede tener relación con la alta radiación que produce un cierto tipo de xeromorfismo. Fahn & Cutler (1992) muestran que hojas anfiestomáticas se presentan en 51% de las especies con caracteres xeromórficos estudiadas por ellos. Parkhust (1978), utilizando modelos para correlacionar hojas anfi e hipostomáticas con factores de ambientes xéricos, mésicos e hidrofíticos, observó que las hojas anfiestomáticas tendrían un mejor desempeño fotosintético. Mott et al. (1982) y Mott & Michaelson (1991) comprobaron que la presencia de estomas en ambas superficies está relacionada con altas intensidades de luz, lo cual podría explicar la presencia de hojas anfiestomáticas en las especies estudiadas que crecen en las primeras etapas de sucesión de cultivo con alta radiación solar.
3. Estomas con distribución difusa, carácter variable para la familia.
4. Cavidad subestomática amplia, es un carácter común para representantes de la familia debido principalmente a la situación de los estomas con respecto a las restantes células epidérmicas.
5. Tricomas tectores espinosos y glandulosos típicos, de representantes de la familia.
6. Mesofilo compacto, empalizada de 1-2 capas de células, también podría considerarse como una adaptación a la alta radiación a la cual están sometidas las plantas estudiadas.
7. Haces vasculares colaterales rodeados por una vaina parenquimática simple. Nervio medio poco pronunciado formado por un haz vascular colateral, caracteres típicos de la familia.
8. Tallos de contorno redondeado. Epidermis de una capa de células con estomas y tricomas tectores y glandulosos. Corteza externa colenquimática, y la media parenquimática, diferenciándose internamente la endodermis de células grandes hialinas.
La presencia de una endodermis diferenciada en los tallos y otros órganos de la planta es un carácter que ha despertado polémica entre los investigadores; así, según Van Fleet (1961), la endodermis puede presentarse en todos los órganos de la planta y el desarrollo de bandas de Caspary está sujeto a factores ambientales tales como la radiación. Dicho autor demuestra que tallos sometidos a períodos de oscuridad desarrollan una endodermis en la que se observa bandas de Caspary, mientras que tallos de la misma especie que crecen en ambientes iluminados no desarrollan endodermis conspicua con bandas de Caspary. Sin embargo, según Beck et al. (1982) la mayoría de las veces la endodermis y el periciclo están ausentes del tallo de las plantas con semilla. Lo encontrado en las especies estudiadas concuerda con las ideas de Van Fleet en el sentido de que no se observan bandas de Caspary ya que las plantas estudiadas crecen en condiciones de alta radiación, por lo que los autores del presente trabajo consideran que no es conveniente denominar a esta capa como endodermoide (como es llamada por Tomlinson 1969) ya que podría tratarse de una endodermis propiamente dicha.
9. Raíces adventicias que parten del tallo. Raíz con epidermis persistente, exodermis bien diferenciada con paredes engrosadas, corteza parenquimática amplia e internamente se diferencia la endodermis, que en sus estados juveniles muestra claramente las bandas de Caspary y en estados posteriores del desarrollo el engrosamiento en forma de U, el periciclo es parenquimático y la estela con un número variable de polos de protoxilema. En algunas raíces la porción central puede estar ocupada por un elemento de vaso xilemático grande. La mayoría de estos caracteres son comunes para la familia.
Presentan también caracteres diferenciales para cada especie:
1. En Commelina obliqua la hoja presenta la epidermis de ambas superficies formada por células con paredes rectas a ligeramente onduladas; en Tinantia erecta las células de la epidermis de ambas superficies son onduladas, carácter que podría darle a la hoja una mayor resistencia ya que esta especie crece más erguida mientras que C. obliqua crece postrada.
2. Estomas en C. obliqua presentan 4 células subsidiarias en ambas superficies, en T. erecta los estomas de la epidermis de la superficie adaxial poseen 4 células subsidiarias mientras que en la superficie abaxial 6 células subsidiarias. El desarrollo de los estomas en estas especies debe ser estudiado con un mayor detalle ya que según Tomlinson (1969) el número de células subsidiarias es un carácter fijo para muchos géneros de Commelinaceae, entre éstos Commelina y Tinantia y las observaciones presentadas en este trabajo no coinciden con lo expuesto con ese autor, mucho menos la diferencia entre la epidermis de las superficies adaxial y abaxial de T. erecta.
3. Parénquima de empalizada con células de forma alargada en C. oblicua y lobadas en forma de H en T. erecta; según Haberlandt (1914), la presencia de parénquima clorofílico en forma de H aparece esporádicamente en los diferentes grupos de plantas vasculares, en dicotiledóneas aparece principalmente en Ranunculaceae pero también en otros grupos, en monocotiledóneas en Bambusa, Calamagrostis, etc. Posiblemente la forma lobada puede ser considerada como una estrategia para la translocación de sustancias en el mesofilo.
La relación del tejido epidérmico con el mesofilo debe ser estudiada con mayor detalle en estas especies. Según Lee & Bowling (1992), la apertura y cierre de los estomas en Commelina communis está influenciada por una sustancia que se produce en el mesofilo de la hoja en presencia de luz pero no en la oscuridad y esta sustancia es producto de la fotosíntesis. Es posible que relaciones similares puedan presentarse en las especies aquí estudiadas y que la diferencia en la morfología celular pueda ser explicada con base en procesos fisiológicos.
La presencia de células con sustancias fenólicas es un carácter común en Commelinaceae y puede estar representando un mecanismo de protección contra herbivoría.
Un carácter importante observado en la vaina foliar de T. erecta es la presencia de células con un contenido en su citoplasma que tiene la apariencia de ser una infección causada por un plasmodio u otro organismo. Para diferentes especies de la familia se ha indicado su carácter como hospederas de enfermedades e infecciones de otros cultivos, entre ellos papa, como es el caso registrado por Brunt et al.(1996), quienes indican la susceptibilidad de T. erecta al ataque de diferentes virus. Debido a ello es probable que estas células puedan estar relacionadas con una posible infección que afecte al cultivo en el que se encuentren.
En el tallo la principal diferencia entre las dos especies es el tipo de arreglo de los haces vasculares. C. obliqua presenta haces vasculares periféricos y perimedulares mientras que T. erecta presenta haces perimedulares y medulares.
CONCLUSIONES
Las especies estudiadas presentan, en sus órganos vegetativos, caracteres anatómicos que les permiten mostrar un comportamiento como malezas, esto es, hojas con epidermis de células altas cubiertas por una cutícula delgada y con tricomas, estomas en ambas superficies, mesofilo compacto con clorénquima bien diferenciado, características éstas que les permiten la colonización en ambientes abiertos como son los estados tempranos de la sucesión de cultivos.
La formación de abundantes raíces adventicias a partir de los tallos permite una rápida propagación vegetativa, lo que favorece su establecimiento en los cultivos.
La presencia de células con un contenido, aparentemente producto de una infección, puede estar indicando el papel de hospedero de enfermedades de cultivo que numerosos autores registran para diferentes representantes de la familia.
Se recomienda continuar con estudios fisiológicos en estas especies para determinar las relaciones mesofilo-epidermis foliar y determinar que tipo de organismos está ocupando las células aparentemente infectadas.
AGRADECIMIENTOS
Este trabajo se realizó en el Laboratorio de Anatomía Vegetal del Centro Jardín Botánico de la Facultad de Ciencias de la Universidad de Los Andes. Los autores quieren agradecer a la Lic. Josefina Peraza por la elaboración de los cortes histológicos, al IIAP-ULA por facilitar la colecta en el cultivo de papas, al Prof. Ramón Jaimez por la colaboración con los datos de campo.
BIBLIOGRAFÍA
1. Alán, E., U. Barrantes, A. Soto & R. Agüero. 1995. Elementos para el manejo de malezas en agrosistemas tropicales. Editorial Tecnológica de Costa Rica. [ Links ]
2. Beck, C.H., R. Schmid & G.W. Rothwell. 1982. Stelar morphology and the primary vascular system of seed plants. Bot. Rev. 48: 691-815. [ Links ]
3. Brunt, A.A., K. Crabtree, M.J. Dallwitz, A.J. Gibbs, L. Watson & E.J. Zurcher (eds.). 1996. Plant Viruses Online: Descriptions and Lists from the VIDE Database. Version: 20th August 1996. URL http://biology.anu. edu.au/Groups/MES/vide/ [ Links ]
4. Fahn, A. & D.F. Cutler. 1992. Xerophytes. Encyclopedia of plant anatomy. Vol 13, Part 3, X. Gebrüder Borntraeger. Berlin, Stuttgart. [ Links ]
5. Haberlandt, G. 1914. Physiological Plant Anatomy. Translated from the fourth German edition by Montagu Drummond. London. [ Links ]
6. Jaimez, R.E., F. Castro & P. Alizo. 2001. Promedios mensuales de precipitación, temperatura máxima y mínima y evaporación registrados en la estación climatológica Santa Rosa desde 1967. IIAP-ULA. 6 pp. Mecanografiado. [ Links ]
7. Johansen, D.A. 1940. Plant microtechnique. Mc Graw- Hill Book. New York-London. [ Links ]
8. Kraus, J.E., H.C. Souza, M.H. Rezende, N.M. Castro, C. Vecchi & R. Luque. 1998. Astra blue and basic fuchsin double staining of plants materials. Biotech. Histochem. 73(5): 235-243. [ Links ]
9. Lee, J. & D.J.F. Bowling. 1992. Effect of the mesophyll on stomatal opening in Commelina communis. J. Exp. Bot. 43(252): 951-957. [ Links ]
10. Luque, R., H.C. Souza & J. Krauss. 1996. Estudio comparativo dos métodos de coloração de Röeser (1972) modificado e Kropp (1972) visando a subtitução do Azul de Astra por Azul de Alcião 8 GS ou 8 GX. Acta Bot. Brasil.10(2): 199-212. [ Links ]
11. Mott, K.A. & O. Michaelson. 1991. Amphistomy as an adaptation to high light intensity in Ambrosia cordifolia (Compositae). Amer. J. Bot. 78: 76-79. [ Links ]
12. Mott, K.A., A.C. Gibson & J.W. OLeary. 1982. The adaptative significance of amphistomatic leaves. Pl. Cell Environm. 5: 455-460. [ Links ]
13. Parkhust, D.F. 1978. The adaptative significance of stomatal occurrence of one or both surfaces of leaves. J. Ecol. 66: 367-383. [ Links ]
14. Sass, J. E. 1951. Botanical microtechnique, 3 Ed. Iowa State College Press. [ Links ]
15. Tomlinson, P.B. 1969. Anatomy of the monocotyledons. III Commelinales-Zingiberales (Metcalfe, C.R. ed.). Clarendon Press. Oxford. [ Links ]
16. Van Flett, D.S. 1961. Hystochemistry and function of the endodermis. Bot. Rev. 27: 165-219. [ Links ]
