











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkActa Botánica Venezuelica
versión impresa ISSN 0084-5906
Acta Bot. Venez. v.28 n.2 Caracas 2005
ANATOMÍA ECOLÓGICA DEL XILEMA SECUNDARIO DE UN BOSQUE SECO TROPICAL DE VENEZUELA
Ecological anatomy of secondary xylem in a tropical dry forest of Venezuela
Williams J. LEÓN H.
Universidad de Los Andes. Facultad de Ciencias Forestales y Ambientales. Departamento de Botánica. Laboratorio de Anatomía de Maderas. wleon@ula.ve
RESUMEN
Se estudió la anatomía de la madera de 51 individuos de especies arbóreas colectados en el bosque El Caimital, que corresponde a un bosque seco tropical, y se correlacionó la estructura anatómica con las condiciones del medio ambiente. Las características estudiadas fueron número de poros por mm2, diámetro de poros, diámetro de punteaduras, longitud de elementos vasculares, tipo de platinas de perforación, presencia de cristales y sílice. Se determinaron los índices de vulnerabilidad, mesomorfía y agrupamiento de vasos. En general se observó un predominio de caracteres que se orientan hacia la eficiencia de la conducción. Según el tamaño de los poros, número de vasos y los índices de vulnerabilidad y mesomorfía, el leño de los individuos estudiados puede designarse como mesomórfico.
Palabras clave: Ecoanatomía, Índice de vulnerabilidad, Mesomorfía, Bosque seco tropical, Disponibilidad de agua, Barinas,Venezuela
ABSTRACT
The wood anatomy of 51 trees from El Caimital forest were studied from an ecological perspective and were compared with data from other habitats. The features studied were vessel frequency, vessel diameter, vessel element length, perforation plates, intervessel pit size, presence of crystals and silica. Vulnerability, mesomorphy and vessels grouping index were calculated. A predominance of characters towards hydraulic efficiency was observed. In accordance with vessel size, vessel number, mesomorphy and vulnerability index, the wood of the studied individuals can be characterized as mesomorphic.
Key words: Ecological wood anatomy, Vulnerability index, Mesomorphy,Water availability, Dry tropical forest, Barinas,Venezuela
INTRODUCCIÓN
La relación entre estructura de la madera y factores climáticos ha sido reconocida desde hace muchos años. Se han realizado diferentes estudios con énfasis en el análisis del efecto de algunos parámetros ecológicos como humedad, temperatura, estacionalidad, altitud y distribución geográfica sobre caracteres anatómicos cuantitativos como diámetro de poros, frecuencia de vasos, grado de agrupamiento de los vasos, longitud de elementos vasculares y tamaño de punteaduras intervasculares. También se ha hecho énfasis en la presencia de engrosamientos espiralados y platinas de perforación escalariformes, así como de un sistema de conducción subsidiario representado por elementos traqueales imperforados (Lindorf 1994). Las tendencias ecológicas han sido interpretadas como el resultado de adaptaciones funcionales a factores ambientales y al considerar el transporte de agua como una de las principales funciones del xilema (Zhang et al. 1992), es lógico esperar que los estudios ecoanatómicos estén orientados hacia aspectos relacionados con los elementos de conducción.
La realización de estudios ecoanatómicos ha tomado en consideración el análisis de la estructura de la madera de diferentes ambientes, desde condiciones de aridez hasta de selva nublada. Las tendencias ecológicas y evolutivas en cuanto al diámetro de los poros, tipo de platinas de perforación, frecuencia de vasos, longitud de elementos vasculares, longitud total de vasos y tipos de fibras han sido discutidas en términos de seguridad y eficiencia en el transporte de agua. Tanto la eficiencia o máxima conductividad como la seguridad están fuertemente relacionadas con el diámetro y frecuencia de vasos, observándose que un aumento del diámetro de los poros incrementa notablemente la eficiencia en cuanto a conducción, pero al mismo tiempo disminuye la seguridad (Sidiyasa & Baas 1998). Los estudios ecoanatómicos han buscado conocer cómo varían algunos elementos xilemáticos ante cambios en las condiciones del medio donde crece la planta (Baas 1973; Dickinson et al. 1978; Oever et al. 1981; Baas et al. 1983; Baas & Carlquist 1985; Carlquist & Hoekman 1985; Zhang et al. 1992; Villagra & Roig 1997; Sidiyasa & Baas 1998; Moglia & López 2001). En Venezuela son pocos los trabajos que se han realizado en este aspecto, destacando los de Pérez (1989), Silva (1991), Lindorf (1994, 1997), León (2001) y Araque & Gámez (2004), donde tomaron como elemento básico de análisis los índices de vulnerabilidad y mesomorfía propuestos por Carlquist (1977), con los cuales se busca determinar el grado de seguridad o eficiencia que presentan los elementos de conducción que se desarrollan en un medio determinado. Estos índices consideran aspectos fundamentales del tejido de conducción como diámetro, frecuencia y longitud del elemento vascular. Algunos autores han argumentado la carencia de significado predictivo o funcional de estos índices, basándose en el hecho de que en algunos casos los valores obtenidos muestran un amplio rango de variación, a pesar de la uniformidad del hábitat (Lindorf 1994). Sin embargo, a pesar de esas amplias variaciones, los índices de Carlquist son los únicos que se han propuesto para determinar relaciones de elementos de conducción con el medio donde crece la planta, y las razones por las cuales son objetados se deben a que parte de esas características tienen que ver con aspectos genéticos y evolutivos propios de cada especie.
En el país se han realizado algunos estudios ecoanatómicos de la madera para diferentes zonas de vida, pero no se tienen reportes para una de las de mayor amplitud y en la cual se encuentra el área de estudio: el bosque seco tropical. En este sentido, el presente trabajo tiene como objetivo establecer la relación entre la estructura de la madera y las características del medio ambiente de individuos de porte arbóreo de diferentes especies que crecen en la zona conocida como bosque El Caimital, Barinas.
MATERIALES Y MÉTODOS
El material de estudio está representado por muestras de madera colectadas en la zona conocida con el nombre de El Caimital, estado Barinas. Esta localidad pertenece a la zona de vida del Bosque Seco Tropical (BST). Según Veillon (1985), las características climáticas generales del área de estudio son las siguientes: precipitación media anual de 1400-1500 mm con 4-5 meses de sequía, temperatura media anual de 26°C, 100-150 m snm, terrenos planos, suelos generalmente arcillosos y pertenecientes a la categoría de Vertisoles. Las muestras estudiadas (Tabla 1) pertenecen a la colección de la Xiloteca MERv del Laboratorio de Anatomía de Maderas y cuentan con el material botánico de respaldo en el herbario MER, ambos adscritos a la Universidad de Los Andes. Para la realización del estudio microscópico se hicieron cortes con un micrótomo de deslizamiento para obtener secciones transversales, radiales y tangenciales de 15-25 µm de espesor y como medio de tinción se utilizó la solución de safranina (Corothie 1967). La preparación de macerados se realizó sumergiendo astillas en una solución de peróxido de hidrógeno y ácido acético glacial (2:1) e introduciéndolas en estufa a 60°C durante 12 horas (Franklin 1937). Las características tomadas en consideración corresponden a diámetro de poros, número de poros por mm2, agrupación de poros, longitud de elementos vasculares, diámetro de punteaduras intervasculares, presencia de traqueidas vasculares o vasicéntricas, tipo de platinas de perforación y presencia de cristales. Para determinar el número de mediciones realizadas en cada uno de los caracteres mencionados y categorizarlos se utilizó lo establecido por IAWA Committee (1989). Adicionalmente se calcularon los índices de vulnerabilidad (IV) y mesomorfía (IM) propuestos por Carlquist (1977), así como el índice de agrupamiento de vasos (IVg). Las fórmulas utilizadas para cada uno de estos índices fueron las siguientes:
IV = Diámetro de poros/poros por mm2
IM = IV x Longitud de elementos vasculares
IVg = Número total de poros / Número de grupos de poros
Se establecieron comparaciones con los valores obtenidos para la selva nublada andina (Pérez 1989), para un bosque muy seco tropical (Lindorf 1994), en la zona de vida de bosque húmedo premontano (Araque & Gámez 2004) y para el bosque higrófilo o manglar (León 2001).
RESULTADOS
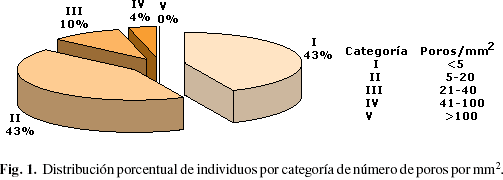
En la Tabla 2 se presentan los valores correspondientes a poros por mm2, diámetro de poros, diámetro de punteaduras, longitud de elementos vasculares, índices de vulnerabilidad, mesomorfía y de agrupamiento de vasos para cada individuo estudiado. Para la frecuencia de poros se observó que más del 80% de los individuos poseen menos de 20 poros por mm2 y dentro de este rango, 43,14% de los individuos presenta un número de poros inferior a 5 por mm2 (Fig. 1).
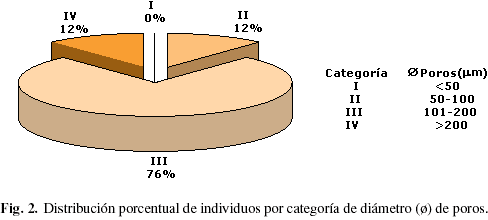
Con relación al diámetro promedio de los poros (Tabla 2), el promedio mínimo fue 71 µm para Sloanea sp. y Trichilia sp. y el máximo fue 240 µm en Ceiba pentandra. La mayor parte de los individuos estudiados poseen poros medianos (Fig. 2), observándose que 76,48% del material de estudio se ubicó en esta categoría.
Con respecto al tamaño de las punteaduras, 9,8% de los individuos presentó punteaduras exclusivamente diminutas, 11,77% punteaduras de diminutas a pequeñas, 1,96% con punteaduras exclusivamente pequeñas, 29,41% con punteaduras pequeñas a medianas, 11,77% de punteaduras exclusivamente medianas, 11,76% con punteaduras medianas a grandes y 21,57% de los individuos presentó punteaduras exclusivamente grandes. En el caso de Hirtella triandra no se observaron punteaduras y este individuo representa 1,96% del total estudiado.
Las platinas de perforación son exclusivamente simples en 94,23% del material estudiado mientras que 5,77% presentó una combinación de platinas simples y múltiples. El desarrollo de platinas múltiples se observó en forma de platinas foraminadas y radiadas en Cytharexyllum sp., escalariformes en Nectandra sp., y reticuladas en Sloanea sp.
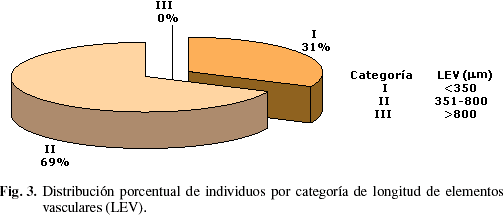
La longitud promedio de los elementos vasculares osciló entre un mínimo de 205 µm en Albizzia guachapele y un máximo de 786 µm en Sapium sp. En la distribución de individuos por categoría de longitud de elementos vasculares se observa que 68,63% de los individuos se ubica en la categoría de elementos vasculares medianos (Fig. 3).
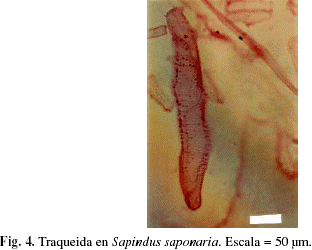
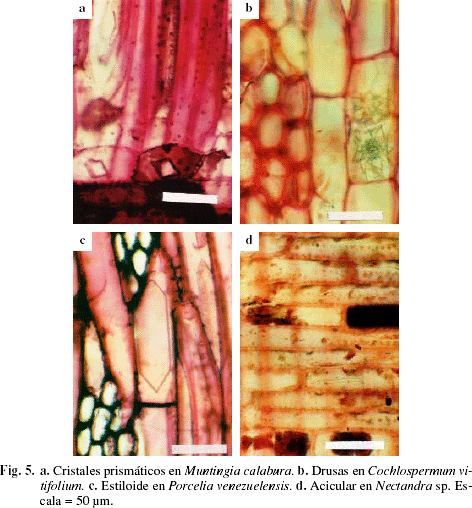
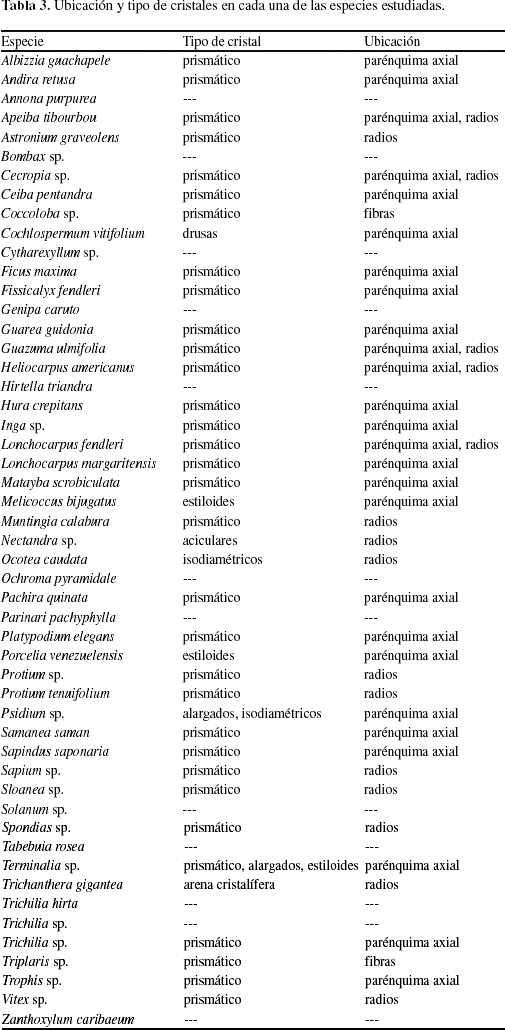
La presencia de traqueidas sólo se observó en la especie Sapindus saponaria (Fig. 4). Los cristales se presentaron en 76,92% de los individuos estudiados y el principal tipo de cristal encontrado fue el prismático (Fig. 5a), aunque también se observaron drusas en Cochlospermum vitifolium (Fig. 5b), estiloides en Melicoccus bijugatus, Porcelia venezuelensis (Fig. 5c) y Terminalia sp., aciculares en Nectandra sp. (Fig. 5d), isodiamétricos en Ocotea sp., Psidium sp., alargados en Psidium sp., Terminalia sp. y arena cristalífera en Trichanthera gigantea. Los cristales comúnmente se presentaron en células parenquimáticas axiales y/o radiales, exceptuando los dos representantes de la familia Polygonaceae (Coccoloba sp., Triplaris sp.), donde los cristales se ubicaron en las fibras (Tabla 3). La presencia de sílice se observó sólo en cuatro especies (Hirtella triandra, Parinari pachyphyllum, Protium tenuifolium, Trichilia sp.), lo cual representa 7,69% del material estudiado.
El valor correspondiente a los índices de vulnerabilidad y mesomorfía (Tabla 2) indican que todos los individuos presentan un leño con comportamiento mesomórfico, sin embargo se observa un amplio rango de variación entre individuos, oscilando el índice de vulnerabilidad entre 1,51 (Sloanea sp.) y 240 (Ceiba pentandra). Se presenta la distribución porcentual de individuos por categorías de índice de vulnerabilidad, observándose que el mayor porcentaje se encuentra en la categoría que incluye individuos con índice de vulnerabilidad menor de diez (Fig. 6). Con relación al índice de mesomorfía (Tabla 2), los valores obtenidos oscilaron entre un mínimo de 343 en Platypodium elegans y un máximo de 187.030 en Ochroma pyramidale. La combinación de poros solitarios y múltiples (Fig. 7, 8), expresada a través del índice de agrupamiento de vasos, alcanzó su valor máximo en Lonchocarpus fendleri con 3,17, mientras que el mínimo se observó en Hirtella triandra, Muntingia calabura y Parinari pachyphylla, en las cuales dicho índice alcanza valores cercanos a la unidad.

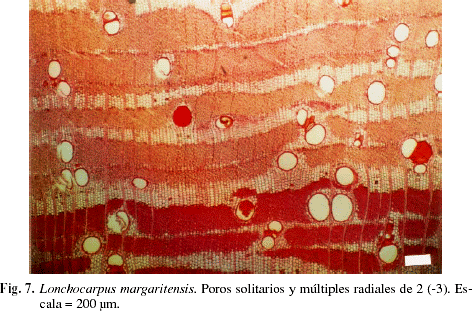
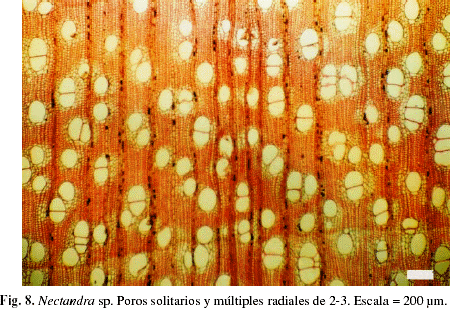
DISCUSIÓN
Según Carlquist & Hoekman (1985), los principales indicadores de mesomorfía son número reducido de vasos por mm2 y por grupo, amplio diámetro de poros, elementos vasculares largos y ausencia tanto de traqueidas como de engrosamientos espiralados. Lindorf (1994) señala que las especies con rasgos mesomórficos desarrollan una estructura conectada con una alta eficiencia, pero una baja seguridad de conducción, lo cual se manifiesta con vasos o poros de mayor tamaño y en menor número por unidad de superficie. Estas características coinciden con lo observado en el presente estudio. Mientras más se aleja el índice de vulnerabilidad del valor correspondiente a la unidad significa una mayor orientación hacia condiciones que garanticen la eficiencia de la conductividad en sacrificio de la seguridad. Cerca de 26% de los individuos estudiados mostraron índices de vulnerabilidad menores de 10, lo que indica eficiencia en conducción combinada con cierto grado de seguridad. El 74% restante se ubicó en valores superiores y aproximadamente 12% de los individuos presentaron índices de vulnerabilidad mayores de 100 lo que indica su mayor orientación hacia la eficiencia en la conducción hidráulica.
Desde el punto de vista de frecuencia de poros, la zona de bosque seco tropical se caracteriza porque los individuos arbóreos que allí se desarrollan tienden a tener un número de poros por mm2 bajo, mientras que en condiciones extremas de aridez (bosque muy seco tropical) o excesiva humedad y salinidad (manglar), la tendencia es a desarrollar mayor número de elementos de conducción. Lindorf (1994) reporta para un bosque muy seco tropical un número de poros por mm2 entre 24 y 208, mientras que para una selva nublada Pérez (1989) menciona valores que van desde 3 hasta 64 poros por mm2 y 50% de los individuos estudiados corresponden a diez o menos poros por mm2, León (2001) reporta valores que varían entre 10 y 40 para una zona de manglar y Araque & Gámez (2004) señalan que en la especie Ochotereneae colombiana que crece en zona de bosque húmedo premontano se encontraron entre 9 y 14 poros por mm2. En la zona de estudio se observó que más de 80% de los individuos estudiados poseen 20 o menos poros por mm2, valores que son notablemente inferiores a los encontrados en condiciones de bosque muy seco tropical o de manglar.
El comportamiento encontrado al comparar los resultados obtenidos con lo reportado para el bosque muy seco tropical y el bosque húmedo montano coinciden con lo mencionado por Zhang et al. (1992) quienes, estudiando la familia Rosaceae, indican que la frecuencia de vasos aumenta a medida que disminuye la disponibilidad de humedad. Igualmente,Moglia & López (2001) señalan que los elementos de los vasos disminuyen su diámetro y longitud, aumentan su frecuencia y espesor de pared celular, así como también tienden a agruparse en mayor proporción para prevenir el colapso bajo condiciones de presiones negativas a medida que aumenta el grado de aridez.
El diámetro de los vasos es probablemente la variable más importante para garantizar la eficiencia en la conducción de agua, observándose que ésta aumenta a medida que se incrementa el diámetro de los vasos; pero al mismo tiempo disminuye la seguridad debido a la relación inversa que mantiene con la frecuencia de poros (Zhang et al. 1992); mientras que los vasos estrechos son más seguros debido a que al encontrarse en mayor número, los embolismos locales no afectan una parte considerable del tejido de conducción (Baas et al. 1983). El hecho que 88,2% de los individuos estudiados se ubiquen en la categoría de poros medianos a grandes señala que para la zona de estudio existe una orientación hacia la eficiencia de la conducción, al igual que ocurre en la selva nublada donde, según Pérez (1989), 65,8% de los individuos poseen poros medianos. Por el contrario, para el bosque muy seco tropical 5,3% de los individuos posee poros medianos y 94,7% poseen poros de muy pequeños a pequeños (Lindorf 1994), mientras que en la zona de manglar 91,3% poseen poros pequeños (León 2001), lo que indica que en estas dos últimas zonas de vida existe mayor tendencia hacia el desarrollo de elementos de conducción con características orientadas hacia la seguridad.
A medida que disminuye la cantidad de humedad disponible se produce un acortamiento en la longitud de los elementos vasculares, esto coincide con lo obtenido en el presente estudio y concuerda con lo que ocurre en otras zonas de vida. Lindorf (1994) señala, para el bosque muy seco tropical, que 84,2% de los individuos desarrolla elementos vasculares cortos, mientras que en la selva nublada andina Pérez (1989) reporta que 55% se ubican en la categoría de elementos vasculares medianos, 13% cortos y 32% largos. Con relación al manglar, León (2001) señala que 50% de los individuos posee elementos vasculares cortos y 50% medianos. Los valores obtenidos en la zona de estudio indican que aproximadamente 69% de los individuos poseen elementos vasculares medianos, lo cual es mayor a lo encontrado en las zonas de vida antes mencionadas. Dentro de las tres zonas de vida comparadas, la menor disponibilidad de humedad ocurre en la zona correspondiente al bosque muy seco tropical y allí es donde se desarrolla la mayor proporción de individuos con elementos vasculares cortos; en la zona de bosque tropical se observa una reducción del número de individuos con elementos vasculares cortos y un notable incremento en la categoría de medianos, pero sin desarrollarse individuos con elementos vasculares largos. En el extremo opuesto, representado por la zona de vida correspondiente a la selva nublada o bosque húmedo montano, disminuye aún más la proporción de individuos con elementos vasculares cortos y se observa un notable porcentaje con elementos vasculares largos.
Al establecer comparaciones con otras zonas de vida, especialmente con la selva nublada o bosque húmedo montano y el bosque muy seco tropical se observa que el comportamiento en la zona de estudio se manifiesta como intermedio entre lo reportado por Lindorf (1994) para el bosque muy seco tropical y lo indicado por Pérez (1989) para el bosque húmedo montano. Esto es una respuesta ante las diferentes condiciones representadas por cada una de estas zonas de vida. Mientras que el área del bosque El Caimital presenta precipitación promedio de 1.400-1.500 mm/año con 4-5 meses de sequía (Veillon 1985), la zona de bosque muy seco tropical tiene precipitación promedio anual de 558 mm, siete meses de sequía y una tasa de evapotranspiración que, en muchos casos, sobrepasa a la precipitación (Lindorf 1994). La zona de selva nublada o bosque húmedo montano reportada por Pérez (1989) corresponde a áreas con más de 2.000 mm de precipitación promedio anual y cerca de 10 meses de lluvia. Tanto en el bosque húmedo montano como en el bosque seco tropical, todos los individuos tienen un comportamiento de tipo mesomórfico, mientras que en el bosque muy seco tropical la tendencia es hacia un comportamiento xeromórfico.
El índice de agrupamiento de vasos presenta valores que están, en la mayoría de los casos, por encima de la unidad. Solamente en las especies Hirtella triandra, Muntingia calabura y Parinari pachyphylla se observaron valores que indican la presencia de poros exclusivamente solitarios. El grado de contacto entre los poros es un elemento importante tanto en la eficiencia como en seguridad de conducción debido a que garantiza la posibilidad de mayores longitudes de desplazamiento de la columna de agua, o sea que representan rutas alternas en caso que ocurran embolismos. En las especies con índice de agrupamiento de vasos cercanos a la unidad, se observó que tienen un diámetro promedio de poros mayor de 150 µm; esto permite inferir que a pesar de existir un número reducido de áreas de contacto entre vasos adyacentes, desarrollan elementos de conducción con diámetros grandes para movilizar la mayor cantidad de humedad posible y aprovechar al máximo las pocas áreas de contacto existentes. Villagra & Roig (1997) señalan que el agrupamiento de los vasos, así como su frecuencia, permiten maximizar la seguridad del sistema hidráulico al proporcionar vías alternativas para la conducción de agua, especialmente en las especies que carecen de traqueidas. Aunque Pérez (1989) y Lindorf (1994) no calcularon el índice de agrupamiento de vasos, es importante notar que para el bosque muy seco tropical 90% de las especies estudiadas poseen vasos múltiples, con un promedio de 3,6 vasos por grupo. Este valor es mayor al observado en el presente estudio donde la mayoría de las especies se caracterizó por poseer una combinación de vasos solitarios y múltiples predominantemente de 2-3 vasos por agrupación. Según Carlquist (1984), citado por Lindorf (1997), el agrupamiento de los vasos garantiza una mayor seguridad en la conducción ya que permite que la movilización de agua que se efectúa a través de un vaso pueda continuar a través de los vasos adyacentes. Tomando en consideración este criterio, es lógico esperar que el índice de agrupamiento de vasos sea mayor en la zona correspondiente al bosque muy seco tropical.
Las punteaduras intervasculares observadas fueron exclusivamente alternas y este tipo de punteaduras es considerado como el más evolucionado, en comparación con los tipos escalariforme y opuesto, y la más eficiente para la conducción. El amplio rango de variación encontrado en el tamaño de las punteaduras está directamente relacionado con los grupos taxonómicos presentes en el área de estudio ya que representantes de familias como Meliaceae se caracterizan por el desarrollo de punteaduras diminutas o pequeñas, mientras que los de la familia Lauraceae tienden a desarrollar punteaduras grandes, independientemente del lugar donde se encuentren.
El valor correspondiente al índice de vulnerabilidad indica que 100% de los individuos presentan un leño con comportamiento de tipo mesomórfico. Este índice trata de expresar la seguridad de conducción ante la diferencia de condiciones ambientales en donde se pueda desarrollar un individuo. Los bajos valores indican una alta redundancia de los vasos o, en otras palabras, una menor vulnerabilidad y los valores menores de uno representan maderas xeromórficas y valores mayores a la unidad corresponden a maderas mesomórficas.
Los valores del índice de mesomorfía tienen un rango de variación mucho mayor que el índice de vulnerabilidad. Esto se debe a que en el cálculo de los valores de mesomorfía se incluye la longitud de los elementos vasculares, característica que está relacionada directamente con el grado de especialización, por lo que especies que crecen en una misma área tendrán tendencias distintas en cuanto al índice de mesomorfía de acuerdo a su grado evolutivo. Por esta razón se consideran más confiables los valores correspondientes al índice de vulnerabilidad, aunque es un hecho aceptado que la longitud de los elementos vasculares tiene una influencia notable sobre la eficiencia de la conducción; sin embargo, la longitud total del vaso es más importante que la de sus elementos individuales y el índice de mesomorfía no toma en consideración la longitud total de los vasos. Oever et al. (1981) indican que además del diámetro y frecuencia de poros, parámetros como la longitud total de los vasos y la proporción de tejido con actividad fisiológica de conducción pueden ejercer una notable influencia sobre la eficiencia del movimiento de líquidos en el xilema.
BIBLIOGRAFÍA
1. Araque, O. & L. Gámez. 2004. Anatomía foliar y xilemática de Ochoterenaea colombiana Barkley. Revista Forest. Venez. 49 (En imprenta). [ Links ]
2. Baas, P. 1973. The wood anatomical range in Ilex (Aquifoliaceae) and its ecological and phylogenetic significance. Blumea 21: 193-258. [ Links ]
3. Baas, P. & S. Carlquist. 1985. A comparison of ecological wood anatomy of the floras of southern California and Israel. IAWA Bull. n.s. 8: 245-274. [ Links ]
4. Baas, P., E. Werker & A. Fahn. 1983. Some ecological trends in vessel characters. IAWA Bull. n.s. 4: 141-159. [ Links ]
5. Carlquist, S. 1977. Ecological factors in wood evolution: a floristic approach. Amer. J. Bot. 64: 887-896. [ Links ]
6. Carlquist, S. & D. Hoekman. 1985. Ecological wood anatomy of the woody southern flora. IAWA Bull. n.s. 6: 319-347. [ Links ]
7. Corothie, H. 1967. Estructura anatómica de 47 maderas de la Guayana venezolana. Laboratorio Nacional de Productos Forestales. Universidad de Los Andes-Ministerio de Agricultura y Cría. Mérida,Venezuela. [ Links ]
8. Dickinson,W., P. Rury & G. Stebbins. 1978. Xylem anatomy of Hibbertia (Dilleniaceae) in relation to ecology and evolution. J. Arnold Arbor. 59: 32-49. [ Links ]
9. Franklin, G. 1937. Permanent preparations of macerated wood fibres. Trop. Woods 49: 21-22. [ Links ]
10. IAWA Committee. 1989. IAWA list of microscopic features for hardwood identification. IAWA Bull. n.s. 10: 219-332. [ Links ]
11. León H.,W. 2001. Anatomía del leño, aspectos ecológicos y filogenia en mangles de Venezuela. Revista Forest. Venez. 45: 191-203. [ Links ]
12. Lindorf, H. 1994. Eco-anatomical wood features of species from a very dry tropical forest. IAWA J. 15: 361-376. [ Links ]
13. Lindorf, H. 1997. Wood and leaf anatomy in Sessea corymbiflora from an ecological perspective. IAWA J. 18: 157-168. [ Links ]
14. Moglia, J. & C. López. 2001. Estrategia adaptativa del leño de Aspidosperma quebracho-blanco. Madera y Bosques 7(2): 13-25. [ Links ]
15. Oever van den, L., P. Baas & M. Zandee. 1981. Comparative wood anatomy of Symplocos and latitude and altitude of provenance. IAWA Bull. n.s. 2: 3-24. [ Links ]
16. Pérez, A. 1989. Caracterización ecoanatómica del leño de 40 especies del bosque La Mucuy, Estado Mérida,Venezuela. Revista Forest. Venez. 33: 43-51. [ Links ]
17. Sidiyasa, K. & P. Baas. 1998. Ecological and systematic wood anatomy of Alstonia (Apocynaceae). IAWA J. 19: 207-229. [ Links ]
18. Silva, A. 1991. Anatomía de la madera de ocho especies propias del bosque de galería (vertiente sur) del Parque Nacional El Ávila. Bol. Soc. Venez. Ci. Nat. 147: 85-136. [ Links ]
19. Veillon, J. 1985. El crecimiento de algunos bosques naturales de Venezuela en relación con los parámetros del medio ambiente. Revista Forest. Venez. 29: 5-122. [ Links ]
20. Villagra, P. & F. Roig. 1997. Wood structure of Prosopis alpataco y P. argentina growing under different edaphic conditions. IAWA J. 18: 37-51. [ Links ]
21. Zhang, S., P. Baas & M. Zandee. 1992. Wood structure of the Rosaceae in relation to ecology, habit and phenology. IAWA Bull. n.s. 13: 307-349. [ Links ]
