










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Facultad de Ciencias Veterinarias
Print version ISSN 0258-6576
Rev. Fac. Cienc. Vet. vol.49 no.1 Maracay June 2008
Caracterización Morfométrica del Híbrido Yaque Pintado (Pseudoplatystoma fasciatum x Leiarius marmoratus) y sus Progenitores (Siluriformes: Pimelodidae)
Francisco J. Mateo *,1 , Nereida Delgado * y Héctor López **
* Instituto de Zoología Agrícola, Facultad de Agronomía, Universidad Central de Venezuela. Apartado 4563, Maracay 2101, estado Aragua, Venezuela. ** Instituto de Zoología Tropical, Facultad de Ciencias. Universidad Central de Venezuela. Correo-E:mateofran@hotmail.com
Resumen
Para conocer las divergencias y/o convergencias en la forma del cuerpo entre el híbrido yaque pintado y sus progenitores, se evaluó por medio de un análisis exploratorio, el contorno corporal total y distintas regiones del cuerpo. Para ello se utilizó un análisis de componentes principales (ACP) con las matrices de covarianza y correlación, valiéndose del método jackknife, con el propósito de verificar cuándo un coeficiente dentro del componente es estadísticamente diferente de cero. Los análisis demuestran que el híbrido exhibe la forma corporal general de la especie progenitora P. fasciatum. Se confirma además, que el ACP con la matriz de correlación, si bien logró extraer en la mayoría de los casos un mejor vector de tamaño isométrico, no permitió verificar las relaciones dentro de los grupos de estudio, ya que sus gráficas por lo general ubicaron a los híbridos en posición intermedia en relación a sus progenitores, dando la impresión de que éstos heredan los caracteres de uno y otro padre prácticamente en la misma proporción. Mientras que al aplicar el ACP con matriz de covarianza, las gráficas sí permitieron visualizar las relaciones entre los grupos.
(Palabras clave: Pseudoplatystoma fasciatum, Leiarius marmoratus, híbridos, fenotipos, heredabilidad, medición del cuerpo)
Morphometric Characterization of the Yaque Pintado Hybrid (Pseudoplatystoma fasciatum x Leiarius marmoratus) and its Parents (Siluriformes: Pimelodidae)
Abstract
In order to elucidate both the divergences and the convergences in body shape between the yaque pintado hybrid and its parents, an exploratory analysis of total body shape and of different body regions was carried out. To test if a coefficient within the component was statistically different from zero, a principal component analysis (PCA) with covariance matrixes and correlation was done, with the aid of the jackknife method. Results of the analyses show that the hybrid exhibits the general body shape of the parental species P. fasciatum. Furthermore, it was also corroborated that the PCA with the correlation matrix allowed to extract a better isometric size vector in most of the cases, but it did not allow to verify the relationships within the groups studied, due to the fact that their graphs in general, placed hybrids in an intermediate position relative to their parents, giving the impression that hybrids practically inherit the characters from each parent in equal proportions. On the other hand, when the PCA, along with the application of the covariance matrix was applied, the resulting graphs did allow the visualization of the relationship between groups.
(Key words: Pseudoplatystoma fasciatum, Leiarius marmoratus, hybrids, phenotypes, heritability, body parts, body measurements)
Recibido: 16/07/07 - Aprobado: 14/05/08.
Introducción
Dentro de la familia Pimelodidae, hay especies con gran potencial para la actividad acuícola. Kossowski y Madrid (1991) y Kossowski (1991,1996, 1999) consideran que, entre las limitantes para su producción, el levante de alevines (debido a la alta tasa de canibalismo en las etapas de larva y postlarva y a los hábitos alimentarios piscívoros) es lo que soporta el mayor peso. Como alternativa, dichos autores sugirieron que la hibridación intergenérica, mediante el cruce de especies carnívoras de alto valor comercial como Pseudoplatystoma spp. y P. hemiliopterus, con especies omnívoras tales como Leiarius marmoratus (bagre yaque) (Gill, 1870), Pimelodus blochi (bagre chorrosco) (Valenciennes, 1840), P. pirinampu (Kossowski, 1999), podría arrojar resultados favorables.
Es así, como
Este estudio exploró las relaciones morfométricas entre las especies parentales y el híbrido yaque pintado mediante un análisis de componentes principales (ACP). Para ello, se utilizaron dos metodologías, ambas a partir del ACP. La primera con una Matriz de Covarianza, recomendada cuando todos los datos tienen la misma escala de medida y la segunda con
El trabajo en general, desarrolla una estrategia sencilla para explorar las semejanzas y/o diferencias de caracteres multidimensionales entre híbridos y sus progenitores. La idea en general no es nueva, puesto que ya por varias décadas ha sido aplicada en los estudios de genética evolutiva (Smith, 1973; Neff y Smith, 1979; Thorpe y Leamy, 1983; Strauss y Fuiman, 1985; Shea, 1985; Strauss, 1986, entre otros).
Materiales y Métodos
Para el estudio se diseñó un único grupo de evaluación, conformado por cuarenta y siete (47) ejemplares, diecisiete (17) individuos (entre 133,80 mm-
Los híbridos fueron suministrados por
Para su estudio, los ejemplares se preservaron en una solución de formol al 10%, primero inyectándolos y luego sumergiéndolos en la misma solución durante dos semanas. Finalmente, se transfirieron a una solución de alcohol etílico al 70% para su conservación definitiva.
Para cada individuo dentro de cada grupo, se consideraron veinte (20) medidas obtenidas por el método de las cerchas propuesto por Strauss y Bookstein (1982), tomadas a partir de diez hitos anatómicos (Zelditch et al., 1995; Dujardin, 1999; Mateo y López, 2005) (Figura 1). Las mediciones se hicieron en el lado izquierdo de cada individuo con un vernier cuya precisión es de - 0,05 mm
De esta manera, el modelo utilizado quedó representado por cinco regiones que describen la cabeza, la parte media del cuerpo, la parte anterior del tronco, la parte posterior del tronco y la cola, como se plantea a continuación(Figura 1):
-Región cabeza (puntos 1, 2 y 10).
-Región media (puntos 2, 3, 9 y 10).
-Región tronco anterior (puntos 3, 4, 8 y 9).
-Región tronco posterior (puntos 4, 5, 7 y 8).
-Región cola (puntos 5, 6, 7).
Aplicación de los componentes principales (ACP) para el contorno corporal total y las regiones corporales
Se construyeron seis (6) matrices: una correspondiente a la región corporal total (con las veinte medidas obtenidas con el método de las cerchas) y las cinco restantes referentes a la región cabeza (V1-2, V1-10 y V2-10), la región media (V2-3, V10-9, V2-10, V3-9, V2-9 y V3-10), la región tronco anterior (V3-9, V4-8, V3-4, V9-8, V3-8 y V4-9), la región tronco posterior (V4-8, V5-7, V4-5, V8-7, V4-7 y V5-8) y la región de la cola (V5-7, V5-6 y V7-6), (Figura 1). Dichas variables conforman las columnas en las distintas matrices, mientras que los individuos (47 ejemplares) constituyen las filas. A continuación, con el programa Palaentological Statistics (PAST) (Hammer et al., 2003) se practicaron los ACP con la matriz de covarianza y correlación y los datos previamente trasformados a logaritmos neperianos (Bookstein et al., 1985; Klingenberg, 1996).
Interpretación de los componentes principales El ACP calcula nuevas variables independientes a partir de las variables originales (Pla, 1986; Peña, 2002), las cuales pueden interpretarse en un estudio morfométrico como factor de tamaño general y factores de forma. La variación debida al tamaño usualmente es asignada por el ACP al primer componente principal, mientras que la variación de figura, definida como aquella que es independiente de la variación de tamaño isométrico se expresa en los componentes sucesivos (Jolicoeur y Mosimann, 1960; Corruccini, 1983; Bookstein et al., 1985; Bookstein, 1989). En este sentido, cuando se aplica un ACP con matrices que contienen muestras distintas, el primer componente principal (1CP) resume la variación de tamaño, donde las cargas dentro del vector reflejarán un promedio de cambios relacionados con el tamaño entre los grupos involucrados, mientras que el segundo componente principal (2CP) y los sucesivos, describen contrastes de figura entre los grupos. Las cargas de los componentes indican el grado en que las distintas variables medidas contribuyen con las diferencias de figura (Bookstein et al., 1985; Bookstein, 1989).
Para las interpretaciones se utilizaron las gráficas bivariadas de los componentes principales (1CP vs. 2CP y 2CP vs. 3CP), y se seleccionó la opción elipses de confiabilidad al 95% de probabilidad que proporciona el programa PAST, con la intención de obtener información de cada grupo en particular y apreciar las relaciones entre ellos (Owen y Chmielewski, 1985). Previamente, se comprobó la normalidad de cada uno de los componentes principales con el supuesto requerido para una correcta interpretación de las elipses de confianza, entendiendo que la distribución en las gráficas será normal bivariada si y sólo si es normal univariante la distribución de cada uno de los componentes (Owen y Chmielewski, 1985; Cuadras, 1991).
Las gráficas empleadas fueron 1CP vs. 2CP para verificar las relaciones de tamaño y figura y el 2CP vs. 3CP para verificar los componentes de figura capaces de establecer diferencias y/o semejanzas entre las tres muestras que componen los grupos. Asimismo, las relaciones entre las muestras para cada grupo de estudio, se interpretaron con la inspección de los signos y magnitudes de las cargas (o coeficientes) para cada carácter en los vectores propios. Los vectores propios que expresan las figuras (2CP y 3CP) tienen coeficientes de diferentes magnitudes y/o signos, mientras que el vector propio que expresa tamaño (1CP), tendrá coeficientes del mismo signo y similar magnitud (Strauss, 1985; Bookstein et al., 1985; Cuadras, 1991; Gallo-Da-Silva et al., 1998).
Debido a que las muestras contienen individuos que varían con respecto al tamaño, edad, sexo e incluso el hábitat, se espera exista tanto variación ambiental como genética en los análisis; tales aspectos interactúan con la figura (Thorpe y Leamy, 1983; Bookstein et al., 1985), por lo que se utilizaron algunas estrategias para minimizar el efecto de éstos dentro de los estudios. Por ello, se consideró además del ACP con la matriz de covarianza, el de correlación, ambos con los datos previamente transformados a logaritmo (Somers, 1986). Además se caracterizó por separado el tamaño (1CP) de la figura (2CP y 3CP), práctica comúnmente empleada en los estudios morfométricos, con el objeto de considerar sólo la contribución de los componentes de figura, para determinar las diferencias o semejanzas (híbridos y progenitores) (Smith, 1973; Neff y Smith, 1979; Thorpe y Leamy, 1983; Strauss, 1986).
Aplicación de la técnica de remuestreo jackknife
Al ACP, se le aplicó la técnica de remuestreo jackknife, con la intención de verificar si existía diferencia estadística en las cargas dentro de los componentes (Gibson et al., 1984; Marcus, 1990; Peres y Bizerril, 1994; Monteiro, 1997). La técnica consiste en calcular n veces un estadístico cualquiera (n= tamaño de la muestra ) en cada oportunidad, excluyendo un ejemplar en la muestra, hasta eliminar (una sola vez) el total de los ejemplares dentro de la muestra. De este modo se crearon para cada evaluación (contorno corporal total, región de la cabeza, región media, región anterior del tronco, región posterior del tronco y región de la cola) tantas matrices como individuos tuvieran los grupos de estudio. En total, se elaboraron doscientas ochenta y dos (282) matrices. Los pasos seguidos fueron los siguientes:
1. Se estimaron los coeficientes de los vectores propios para el total de individuos (Coef. totales).
2. Se obtuvieron n series de Coef. (n = tamaño de la muestra); cada serie con un individuo diferente excluido, esto genera los estimadores jackknife = Coef.jk.
3. Se convirtió cada estimador (Coef.jk) en pseudovalores (Coef.pv) por medio de la ecuación:
Coef. pvi = (n x Coef. total) - [(n-1) x Coef. jki]
4. El valor Jackknife para cada Coef. es el promedio de sus pseudovalores y éste es el mejor estimador del Coef .
Coef. v = ( ∑ Coef. pvi)/n.
5. Los pasos anteriores se repitieron para cada coeficiente dentro de cada grupo de estudio por evaluación.
6. Los pseudovalores fueron utilizados luego en las pruebas de t (previamente confirmado el supuesto de normalidad) para determinar si el coeficiente es estadísticamente diferente de cero (0) y por tanto con una contribución significativa a un componente dado (Gibson et al., 1984).
Resultados
Comparación del contorno corporal total
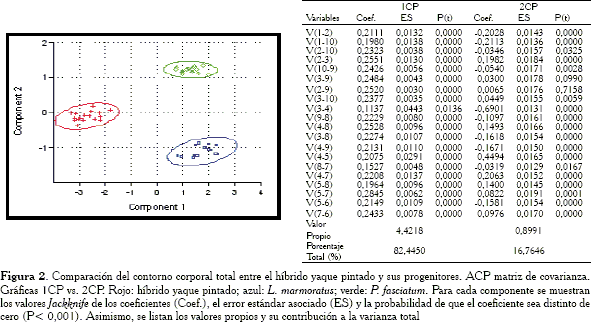
En la gráfica 1CP vs. 2CP (Figura 2) se observan tres distribuciones. Como es obvio en los análisis de componentes principales, el primer componente separa los grupos por tamaño; es así como se percibe a las especies P. fasciatum y L. marmoratus hacia la derecha, mientras que los individuos híbridos yaque pintado se encuentran hacia la izquierda. En el segundo componente, la posición del híbrido, aunque intermedia, se acerca más a L. marmoratus en comparación con la otra especie progenitora. Debido a que en las elipses de confianza los ejes mayores son aproximadamente paralelos al primer componente, entonces es de esperar que buena parte de la información contenida en el componente esté relacionada con el tamaño isométrico. Asimismo, como los ejes menores son aproximadamente paralelos al segundo componente, éste explica buena parte de la información debida a la figura.
Al analizar
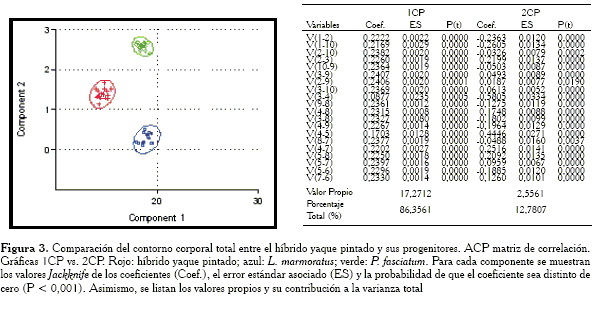
En la gráfica obtenida a partir de la matriz de correlación (Figura 3) se aprecia que el 1CP también separa a los grupos por tamaño, aún cuando puede observarse que esta separación se suaviza. Sin embargo, en el 2CP, la posición del híbrido es prácticamente intermedia en relación a los progenitores. Se observa además en el cuadro, que todos los coeficientes son estadísticamente diferentes de cero (0) en el 1CP, mientras que en el 2CP la variable V8-7 no es estadísticamente distinta de cero.
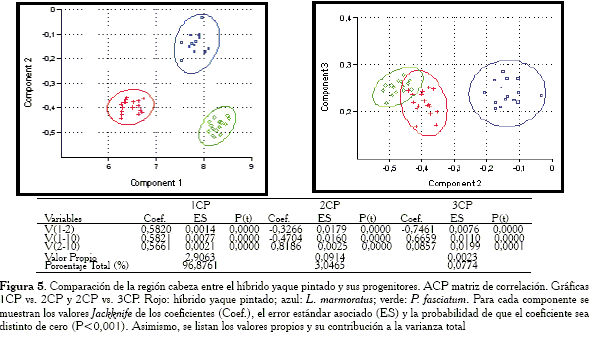
Comparación de la región cabeza
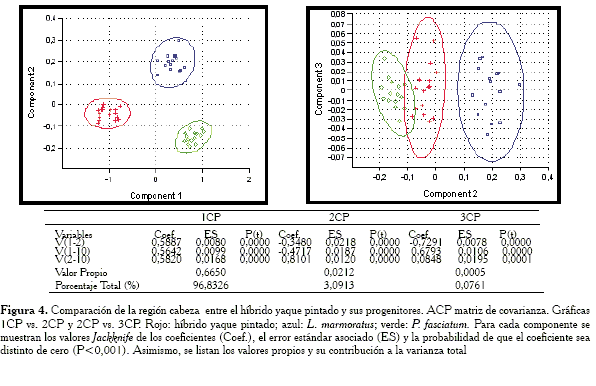
En este caso las gráficas (Figuras 4 y 5) muestran como la matriz de correlación y la covarianza presentan resultados muy similares: en ambas la elipse del híbrido yaque pintado y la especie P. fasciatum se solapan, lo que indica que para este conjunto de medidas hay coincidencia. Asimismo, se observó en las Tablas (Figuras 4 y 5), que tanto para la matriz de covarianza como de correlación, el 1CP puede definirse perfectamente como un vector de tamaño isométrico. Al considerar las cargas del 2CP se puede inferir que la agrupación de las muestras híbrido yaque pintado y P. fasciatum tal como se observan en las gráficas, se debe a las variables V1-2, V1-10 y V 2-10.
En los componentes todas las cargas son distintas de cero (P<0,001).
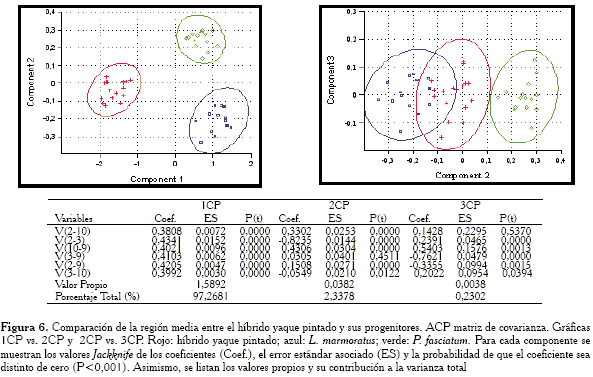
Comparación de la región media
En las gráficas (Figura 6) obtenidas con la matriz de covarianza, el primer componente separa perfectamente a los grupos por tamaño, mientras que el 2CP ubica a los híbridos en posición intermedia con respecto a los progenitores, pero su elipse se solapa con L. marmoratus. Esto indica que para esta región, tanto el híbrido como la especie L. marmoratus serían similares. Entre las variables responsables de esta similitud están V2-10, V2-3 y V10-9.
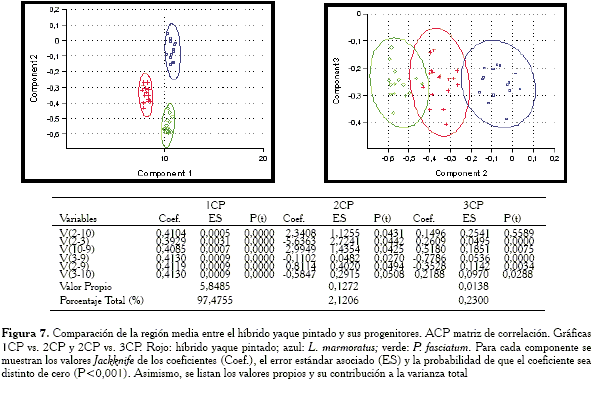
En las gráficas (Figura 7) obtenidas con la matriz de correlación, se aprecia que en el 2CP el híbrido ocupa prácticamente una posición intermedia. No obstante, al observar en el cuadro las cargas del 2CP se verifica que éstas no son estadísticamente distintas de cero (P>0,001), por lo que se descartaría su interpretación y se tomaría únicamente la proporcionada por la matriz de covarianza.
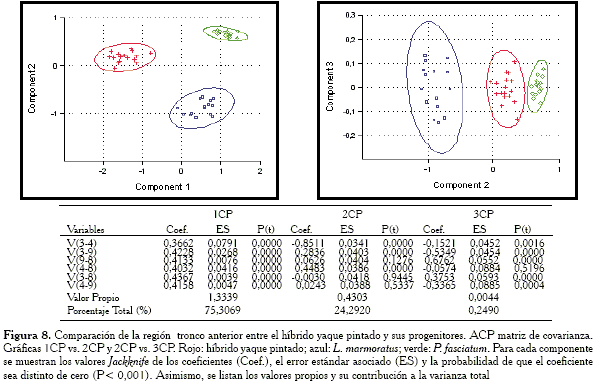
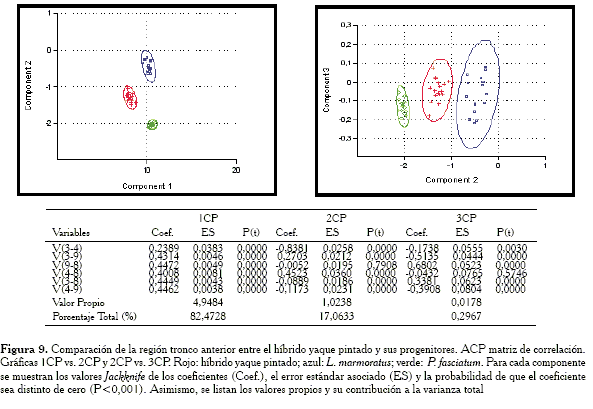
Comparación de la región tronco anterior
Para esta región (Figura 8) se observa al híbrido en posición intermedia con relación a las especies parentales; sin embargo, más cercano a P. fasciatum, pero sin llegar a solaparse las elipses de confianza, mientras que en las gráficas (Figura 9) obtenidas con la matriz de correlación, los híbridos ocupan una posición prácticamente intermedia.
En el cuadro anexo a las gráficas (Figuras 8 y 9) se verifica que tanto la matriz de covarianza como la de correlación extraen en su 1CP un vector de tamaño isométrico, aún cuando éste es más convincente en el caso del componente a partir de la matriz de covarianza.
Las variables responsables de las semejanzas entre el híbrido y la especie P. fasciatum para el ACP con la matriz de covarianza son: V3-9, V4-8 y V3-4 .
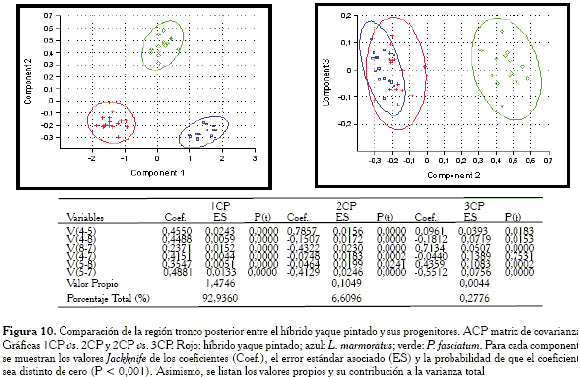
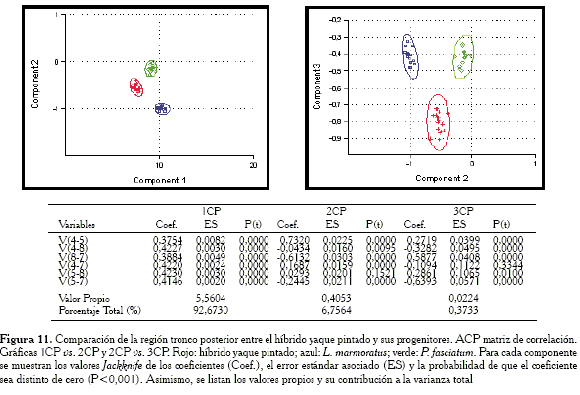
Comparación de la región tronco posterior
En las gráficas (Figura 10) se puede evidenciar que los híbridos son agrupados para esta región junto con el progenitor L. marmoratus. Como se solapan ampliamente sus elipses de confianza, entonces ambos exhiben el mismo contorno para esta región. Las variables responsables de la igualdad entre el híbrido y L. marmoratus son: V4-5, V8-7 y V5-7.
Cuando se evalúan las gráficas obtenidas con la matriz de correlación (Figura 11), el híbrido ocupa una posición intermedia.
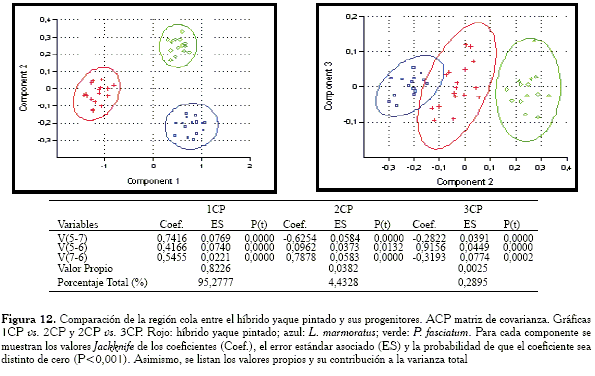
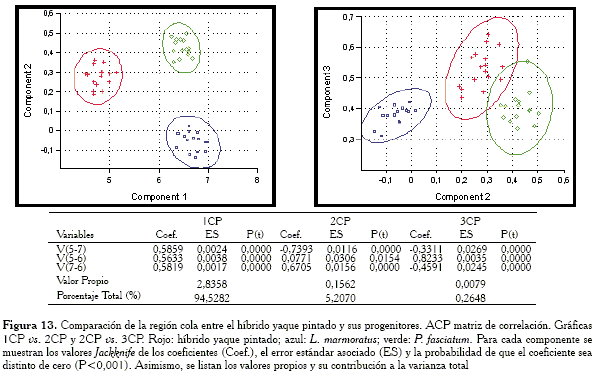
Comparación de la región cola
Para esta región, las gráficas obtenidas con la matriz de covarianza y correlación muestran respuestas distintas (Figuras 12 y 13). En el primer caso, el híbrido se encuentra en posición intermedia pero solapando un poco la elipse de confianza con la especie L. marmoratus, mientras que con la matriz de correlación la elipse del híbrido se pone en contacto con la especie P. fasciatum. Las variables responsables de las similitud en ambos casos (ACP con matriz de covarianza y ACP con matriz de correlación) son: V5-7 y V7-6.
Al observar las cargas del primer componente, se tiene que en el caso de la matriz de covarianza, éste dista de lo que sería un vector de tamaño isométrico puesto que los pesos de las cargas son muy distintos entre sí. Por el contrario, el 1CP con la matriz de correlación sí parece extraer un vector de tamaño isométrico.
Discusión
Los distintos ACP aplicados a los contornos regionales confirman lo siguiente: las características consideradas en un híbrido no siempre son intermedias en relación al intervalo de variación de dichas características en las especies progenitoras (Smith, 1973; Thorpe y Leamy, 1983; Strauss, 1986; Purdom, 1993). Lo anterior es cierto principalmente cuando el estudio morfométrico parte de una matriz de covarianza (Figuras 4, 6, 8, 10 y 12,) con la cual se pudo constatar que los híbridos, en forma moderada y en algunos casos ampliamente, entraron en el intervalo de uno de los progenitores, demostrando así que compartían más similitud con un progenitor en contraste que con el otro. Por el contrario, en la mayoría de los casos en los cuales se parte de una matriz de correlación para la aplicación del ACP, se observó que las diferencias y/o semejanzas para los caracteres considerados parecen promediarse y es por eso que en las gráficas de los componentes, se observa a los híbridos en posición intermedia con respecto a sus progenitores (Figuras 3, 7, 9 y 11).
Las cargas de las variables en los componentes indican la extensión en que cada variable contribuye a la variación en estos componentes. La técnica jackknife permitió asociar a cada carga una probabilidad, indicando si éste estaba estadísticamente correlacionado o no con el componente.
Por lo general en los estudios, las cargas en el primer componente principal fueron positivas y aproximadamente de la misma magnitud, por lo que éste correspondió al incremento conjunto de todas las variables, indicando así que efectivamente el componente expresa tamaño. Sin embargo, no siempre se pudo constatar un tamaño isométrico, es decir, aquél sin alteración de la forma. Esto ocurrió en la mayoría de los ACP con matriz de covarianza (Figuras 6, 8, 10 y 12).
Las cargas en el segundo componente principal fueron positivas y negativas y en el caso de los análisis que partieron de una matriz de covarianza, éstas indicaron cuáles variables son responsables de las agrupaciones. Por área de evaluación éstas fueron:
-Región cabeza: las variables responsables de la agrupación entre el híbrido y P. fasciatum son: V1-2, V1-10 y V2-10.
-Región media: las variables responsables de la agrupación entre el híbrido y L. marmoratus son: V2-10, V10-9 y V2-3.
-Región tronco anterior: las variables responsables de la agrupación entre el híbrido y P. fasciatum son: V3-9, V4-8 y V3-4.
-Región tronco posterior: las variables responsables de la agrupación entre el híbrido y L. marmoratus son: V4-5, V8-7 y V5-7.
-Región cola: las variables responsables de la agrupación entre el híbrido y L. marmoratus son: V5-7 y V7-6.
-Contorno corporal total: las variables responsables de la agrupación entre el híbrido y L. marmoratus son: V1-2, V1-10, V2-3, V3-4, V4-8, V3-8, V4-9, V4-5, V4-7 y V5-6.
Puesto que las distancias entre las observaciones en el plano de los componentes se interpretan en términos de patrones de similitud de respuesta al conjunto de variables (Peña, 2002), se puede entonces considerar que aquellos casos en los cuales las elipses de confianza se solapan (para los ACP con matriz de covarianza), los individuos serán similares para esa región (Figura 4, 6 y 12 ); y, en especial, en aquellos casos en los cuales prácticamente los centroides de las nubes de puntos coinciden, podría afirmarse que las muestras bajo estas condiciones son iguales (Figura 10). Esto es válido si se interpreta al centroide como el individuo promedio dentro de cada elipse de confianza (Peña, 2002).
Es importante señalar que efectivamente tal como sugiere Somers (1986), los análisis morfométricos con un ACP partiendo de una matriz de correlación, logran, como se observa en la mayoría de los resultados extraer en el primer componente un mejor vector de tamaño isométrico (Figuras 3, 7, 11, 13), en comparación con el componente extraído con una matriz de covarianza. Sin embargo, la metodología presenta un serio inconveniente cuando se llevan las interpretaciones a las gráficas. Tal como se mencionó al inicio de la discusión, en las representaciones, el híbrido suele ubicarse en posición intermedia en relación a sus progenitores, dando la impresión de que éstos heredan los caracteres de uno y otro padre, prácticamente en la misma proporción.
Comparación del híbrido y sus progenitores
El análisis con la matriz de covarianza evidencia que para la forma corporal general, el híbrido ocupa aproximadamente una posición intermedia con respecto a los progenitores. Por el contrario, en las regiones de la cabeza (Figura 4) y el tronco anterior (Figura 8) se observan patrones de similitud entre el híbrido y P. fasciatum, mientras que en la parte media (Figura 6), la región del tronco posterior (Figura 10) y la cola (Figura 12) se nota la relación del híbrido con L. marmoratus. Lo anterior puede interpretarse como un híbrido cuyo contorno total es intermedio entre los progenitores, pero con la parte anterior del cuerpo (cabeza y parte media), similar a P. fasciatum. En un estudio alométrico, Mateo y López (2005) comentan al respecto, que a simple vista el híbrido parece heredar buena parte de los caracteres morfométricos del progenitor P. fasciatum. Castillo et al. (1995) basados en un análisis de componentes principales sostienen que el híbrido es prácticamente intermedio entre los progenitores aun cuando se observa un ligero acercamiento hacia el progenitor P. fasciatum. Se ha de resaltar que en su estudio no se especifica el tipo de matriz utilizada y no emplea elipses de confianza, las cuales podrían haber ilustrado en forma más clara las relaciones entre los grupos.
Resulta interesante que el híbrido yaque pintado exprese buena parte de la conformación del progenitor P. fasciatum, ya que en el país, el Rayado (P. fasciatum) es considerada la especie de bagre de mayor demanda comercial (Román, 1985; Barbarino y Taphorn, 1995; Kossowski, 1999).
A pesar de que el presente trabajo es de tipo descriptivo, representa un aporte a los trabajos de Smith (1973), Neff y Smith (1979), Thorpe y Leamy (1983), Shea (1985), Strauss y Fuiman (1985), Strauss (1985, 1986), Brooks (1991), Wilson y Herbert (1993), Castillo et al. (1995), Agnese et al. (1997), ya que se emplearon dos metodologías que proporcionan significancia estadística (elipses de confianza y la técnica de remuestreo jackknife aplicada a las cargas).
Figuras y Tablas
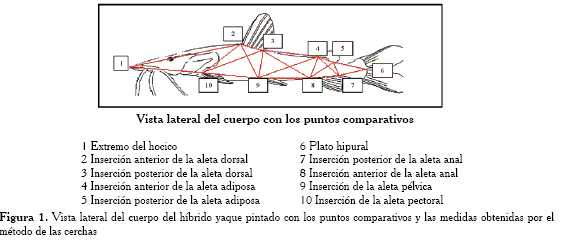
Figura 1. Vista lateral del cuerpo del híbrido yaque pintado con los puntos comparativos y las medidas obtenidas por el método de las cerchas
Referencias
1. Agnese, J.; Teugels, G.; Galbuseras, P.; Guyomard, R.; Volckaert, F. 1997. Morphometric and genetic characterization of sympatric populations of Clarias gariepinus and C. anguillaris from Senegal. J. Fish. Biol., 50: 1143-1157. [ Links ]
2. Barbarino, A.; Taphorn, D. 1995. Especies de la pesca deportiva. UNELLEZ- Fundación Polar. pp. 64-97. [ Links ]
3. Bookstein, F. 1989. Size and shape: A comment on semantics. Syst. Zool., 38: 173-180. [ Links ]
4. Bookstein F.; Chernoff, R.; Elder, J.; Humphries, G.; Smith, Y.; Strauss, R. 1985. Morphometrics in evolutionary biology. The
5. Brooks, M. 1991. The ontogeny of sexual dimorphism: Quantitative models and a case study in Labrisomid blennies (Teleostei: Paraclinus). Syst. Zool., 40: 271-283. [ Links ]
6. Castillo, O.; Nass, P.; Kossowski, C. 1995. Análisis morfométrico multivariado del bagre yaque pintado, un híbrido artificial entre Pseudoplatystoma fasciatum y Leiarius marmoratus (Siluriformes, Pimelodidae). Biollania, 11: 13-26. [ Links ]
7. Corruccini, R. 1983. Principal components for allometric analysis. Am. J. Physical Anthropol., 60: 451-453. [ Links ]
8. Cuadras, M. 1991. Métodos de Análisis Multivariante. Promociones y Publicaciones Universitarias, S. A. Barcelona, España. pp. 70-72. [ Links ]
9. Dujardin, J. 1999. Introducción a a Morfometría, con énfasis en Triatominae y Phlebotominae. I Curso Internacional de Morfometría. La Paz, Bolivia. Resumen. s/p.
10. Gallo-Da-Silva, V.; Cavalcanti, M.; Duarte, P. 1998. Comparative Morphometrics of Semionotidae, Lepisosteidae and Amiidae (Actinopterygii: Neopterygii) by Multivariate Analysis of Truss Networks. Ann. Acad. Bras. Cien., 70: 117-123. [ Links ]
11. Gary, W. 1984. Multivariate Morphometric Variability in Pacific Salmon: Technical Demonstration. Can. J. Fish. Aquat. Sci., 41: 1150-1159. [ Links ]
12. Gibson, A.; Baker, A.; Moeed, A. 1984. Morphometric variation in introduced populations of the common myna (Acridotheres tristis): an application of jackknife to principal component analysis. Syst. Zool., 33: 408-421. [ Links ]
13. Hammer, O.; Harper, D.; Ryan, P. Copyright 2003. PAST - Palaentological Statistics, version 0.98. http://folk.uio.no/ohammer/past. [ Links ]
14. Jolicoeur, P. y Mosimann, J. 1960. Size and Shape Variation in the Painted Turtle. A Principal Component Analysis. Growth, 24: 339-354. [ Links ]
15. Klingenberg, C. 1996. Multivariate Allometry. Advances in Morphometrics. Plenum Press, New York. pp. 23-49. [ Links ]
16. Kossowski, C. 1991. Experiencias iniciales sobre la Hibridización de Leiarius marmoratus (Gill) 1871 por Pseudoplatystoma fasciatum (Linnaeus) 1766 (Pisces, Siluriformes, Pimelodidae). Acta. Cient. Venez., 42: 48-50. [ Links ]
17. Kossowski, C. 1996. Reproducción inducida del bagre cajaro y avances sobre su hibridación con dos especies de pimelodidos (Pisces, Siluriformes). Bioagro. 8: 4-20. [ Links ]
18. Kossowski, C. 1999. Perspectivas del cultivo de bagre en Venezuela. En Congreso Sur-Americano de Acuicultura, 2°. Puerto La Cruz ( Venezuela). Memorias, Tomo I. pp. 287-297. [ Links ]
19. Kossowski, C.; Madrid, F. 1991. Observaciones de los Estadios Embrionario y Larval del bagre rayado cabezón Pseudoplatystoma fasciatum (Linnaeus) 1766 (Pisces, Siluriformes). Biollania, 8: 9-15. [ Links ]
20. Marcus, L. 1990. Traditional morphometrics. The University of Michigan Museum of Zoology, Special Publication No. 2 Ann Arbor. pp. 77-122. [ Links ]
21. Mateo, F.; López, H. 2005. Comparación alométrica entre los híbridos yaque pintado (Pseudoplatystoma fasciatum x Leiarius marmoratus) y chorrosco (Pseudoplatystoma fasciatum x Pimelodus blochi) (Siluriformes: Pimelodidae). Rev. Fac. Cs. Vets. UCV. 46: 99-107 [ Links ]
22. Monteiro, L. 1997. Allometric growth and functional integration in the skull of the black caiman Melanosuchus niger (Crocodylia: Alligatoridae). A jackknife approach. Rev. Brasil. Biol., 57: 31-37. [ Links ]
23. Neff, N.; Smith, G. 1979. Multivariate Analysis of Hibrid Fishes. Syst. Zool., 28: 176-196. [ Links ]
24. Owen, J.; Chmielewski, M. 1985. On Canonical Analysis and the Construction of Confidence Ellipses in Systematic Studies. Syst. Zool., 34: 366-374. [ Links ]
25. Peña, D. 2002. Análisis de datos multivariantes. Mc Graw- Hill. Madrid, España. 536 p. [ Links ]
26. Peres, P.; Bizerril, C. 1994. The Jackknifing of multivariate allometric coefficient (Jolicouer, 1963): A case study on allometry and morphometric variation in Corydoras barbatus (Quoy & Gaimard,1824) (Siluriformes, Callicthydae). Arq. Biol. Technol., 37: 449-454. [ Links ]
27. Pla, L. 1986. Análisis multivariado: métodos de componentes principales. Serie matemática. Organización de los Estados Americanos. Washington D.C. 92 p. [ Links ]
28. Purdom, C. 1993. Genetics and Fish Breeding. Chapman and Hall. 270 p. [ Links ]
29. Román, B. 1985. Peces de Agua Dulce de Venezuela. Fundación Científica Fluvial de los Llanos. Caracas, Venezuela. 175 p. [ Links ]
30. Shea, B. 1985. Bivariate and Multivariate growth allometry: statistical and biological considerations. J. Zool., Lond., 206: 367-390. [ Links ]
31. Smith, G. 1973. Analysis of several hybrid cyprinid fishes from western
32. Somers, K. 1986. Multivariate allometry and removal of size with principal components analysis. Syst. Zool., 35: 359-368. [ Links ]
33. Strauss, R. 1985. Evolutionary allometry and variation in body form in the South American catfish, Genus Corydoras (Callichthyidae). Syst. Zool., 34: 381-396.
34. Strauss, R. 1986. Natural hybrids of the freshwater sculpins Cottus bairdi and Cottus cognatus (Pisces: Cottidae): electrophoretic and morphometric evidence. Am. Mid. Nat., 115: 87-105.
35. Strauss, R.; Bookstein, F. 1982. The truss: body form reconstructions in morphometrics. Syst. Zool., 31: 113-135.
36. Strauss, R.; Fuiman, L. 1985. Quantitative comparisons of body form and allometry in larval and adult Pacific sculpins (Teleostei: Cottidae). Can. J. Zool., 63: 1582-1589.
37. Thorpe, R.; Leamy, L. 1983. Morphometric studies in inbred and hybrid house mice (Mus sp.): Multivariate analysis of size and shape. J. Zool., 199: 421-432.
38. Wilson, C.; Hebert, P. 1993. Natural Hybridization between Artic Char (Salvelinus alpinus) and Lake Trout (S. namaycush) in the Canadian Artic. Can. J. Fish. Aquat. Sci., 50: 2652-2657.
39. Zelditch, M.; Fink, W.; Swiderski, D. 1995. Morphometrics, homology and phylogenetics: quantified characters as synapomorphies. Syst. Biol., 44: 179-189. [ Links ]

