











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.26 n.9 Caracas set. 2001
RASGOS BIOLÓGICOS DE Salix humboldtiana Willd. Y RÉGIMEN DE PULSOS DE INUNDACIÓN
Jorge Liotta
Jorge Liotta. Licenciado en Ciencias Biológicas, Universidad Nacional del Sur. Especialización en Ecología Acuática Continental, Universidad Nacional del Litoral. Responsable del Museo de Ciencias Naturales "Rvdo. P. Antonio Scasso", en San Nicolás. Dirección: Museo "P. Scasso", Don Bosco 580, (2900), San Nicolás, Buenos Aires, Argentina. e-mail: jliotta@cablenet.com.ar
Resumen
El pulso de inundación es el principal regulador en el establecimiento de los bosques de sauce criollo (Salix humboldtiana Willd.) del río Paraná inferior. El control se expresa principalmente a través de dos procesos: a) la aparición de bancos colonizables por esta especie que, por sedimentación pueden llegar a transformarse en albardones y b) la erosión fluvial, que elimina progresivamente los bosques ya establecidos. Varios rasgos de la biología del sauce, como producción de gran cantidad de semillas que germinan en ambientes muy húmedos y alejados de los árboles de la generación anterior, alta velocidad de crecimiento, adaptaciones a la inundación, han permitido su supervivencia en este ambiente tan dinámico.
Summary
The flood pulse is a key-factor in the eccesis of the Salix humboldtiana Willd. (creole willow) forests in the lower Paraná River. The control is expressed mainly by two processes: a) the appearance of banks suitable for the colonization by these trees. These gallery forests also increase the soil level through retention of sediments by the woods determining the levee formation, and b) the erosion, which progressively remove the levees with the mature trees. Several traits of the biology of the willow, such as production of great quantity of small seeds that germinate in very humid substrates, and far from the trees of the previous generation, high speed of growth, adaptations to the flood, have allowed their survival in this highly dynamic environment.
Resumo
O pulso de inundação é o fator fundamental no estabelecimento das matas de Salix humboldtiana Willd. no baixo rio Paraná. O controle é expressado principalmente por dois processos: a) o aparecimento de bancos para a colonização por isto espécies que então, por causa de uma sedimentação aumentado pela presença das matas, chega a ser um dique marginal, e b) a erosão que progressivamente elimina os bosques já estabeleceu. Várias características da biologia do sauce, produção de grande quantidade de sementes que germinam em sustratos muito úmidos e longe das árvores da geração prévia, alta velocidade de crescimento, adaptações para a inundação, permitem a sobrevivência deles num ambiente altamente dinâmico.
PALABRAS CLAVE / Ecología / Río Paraná / Salix humboldtiana / Régimen de Pulsos /
Recibido: 15/06/2001. Modificado: 31/08/2001. Aceptado: 10/09/2001
Introducción
Los grandes ríos siempre han sido sistemas difíciles de estudiar por sus dimensiones y su sorprendente dinámica espacial y temporal. Recientemente han surgido teorías y conceptos que pretenden explicar su funcionamiento (Vanotte et al., 1980; Junk et al., 1989). En la teoría más ajustada para sistemas de este tipo, Junk et al. (1989) postulan que la dinámica del sistema formado por el río más su llanura aluvial está controlada fundamentalmente por el llamado "pulso de inundación". Este concepto fue reformulado posteriormente por Neiff (1990) para quien la fase de sequía es tan importante como la de inundación en el control de muchos procesos del sistema, resultando por esto el concepto de "pulso de inundación-sequía" o, simplemente pulso. El río Paraná, con 3998km de longitud y un caudal promedio de 16000m3/seg, tiene un amplio sistema de llanura aluvial, que en el sector estudiado aquí alcanza unos 60km de ancho (Bonetto y Wais, 1989). Está dividido en dos brazos principales: Paraná Guazú (1,5km de ancho) y Paraná Pavón (0,6km de ancho).
Según mediciones tomadas entre marzo 81 y mayo 82 (Depetris y Kempe, 1990) el río Paraná transporta 65,9x106 Tm·año-1 de sedimentos. Éstos generan las numerosas y dinámicas geoformas que, en conjunto, integran la llanura aluvial.
La vegetación de estos ambientes encuentra variaciones en el plano horizontal, en dirección transversal al eje de escurrimiento y de magnitud decreciente con la distancia a los cauces principales; en el plano vertical, las variaciones son originadas por fluctuaciones del nivel del agua que periódicamente inunda grandes extensiones, que fueran calificadas como zonas de transición acuático-terrestre (ATTZ por sus siglas en inglés) por Junk et al. (1989). Según Neiff (1986), la mayor o menor tolerancia a las fases de inundación y sequía es el factor que controla la distribución de las poblaciones y estructura las unidades de vegetación, tanto herbáceas como arbóreas.
Entre las especies que mejor soportan las fases de inundación se encuentra el sauce criollo (Salix humboldtiana Willd.). Este árbol, el más abundante de este tramo del río, crece hasta 15 o más metros de altura y se distribuye en ríos y riberas arenosas de América cálida, alcanzando el norte de la Patagonia (Cabrera y Zardini, 1993).
De esta especie se conocen varias adaptaciones morfológicas y fisiológicas, como son la presencia de raíces adventicias utilizadas en la absorción de nutrientes y oxígeno, en la atenuación del escurrimiento y en el consiguiente aumento del nivel del suelo (Depetris et al., 1992); la capacidad de reordenar el sistema radicular y el follaje en respuesta a sedimentaciones en gran escala; la posibilidad de traslocar oxígeno desde el tronco a las raíces ubicadas en suelos que, después de inundados, se hacen anóxicos (Neiff, 1986; Neiff et al., 1985); la gran capacidad de reproducirse vegetativamente a partir de ramas quebradas; etc.
Por otra parte, es notable la peculiaridad que presenta esta especie arbórea en su reproducción sexual, ya registrada por una serie de autores desde hace varias décadas (Burkart, 1957; Franceschi y Lewis, 1979). Las semillas germinan alcanzando densidades de miles por metro cuadrado y creciendo rápida y uniformemente.
Sin embargo, aun poseyendo estas adaptaciones, los bosques de sauce criollo no se encuentran dispersos por toda la llanura aluvial.
El objetivo de este trabajo es analizar la influencia de los pulsos en la ocurrencia de los bosques de S. humboldtiana en el tramo inferior del río Paraná.
Hipótesis
Se plantean las siguientes hipótesis:
1. Las poblaciones de sauce sólo pueden establecerse en sitios con una elevada tasa de cambio geomorfológico.
2. Si es cierto que los sauces sólo nacen en bancos de cauce o sitios similares y, dado que los bosques "maduros" aparecen característicamente en zonas de albardones, debe encontrarse una relación directa entre el gradiente de los niveles topográficos de los albardones y las edades de los bosques que se encuentran sobre ellos.
3. Dado que los sitios ocupados por albardones no cumplen con las características requeridas para el establecimiento de bosques, no debe hallarse renovación significativa de los mismos.
4. Debido a la alta tasa de cambio geomorfológico propia de los sectores ocupados por los bosques, los fenómenos de erosión deben afectar intensamente a los bosques maduros.
Materiales y Métodos
Análisis cartográficos
Se utilizaron las cartas topográficas del Instituto Geográfico Militar de Argentina, denominadas "San Nicolás de los Arroyos" (Nº 3360-21-4) de 1970 y "Planta General Savio" (Nº 3360-27-2) de 1967, y las imágenes satelitales (escala 1:50000) denominadas de igual manera correspondientes al año 1996.
Sobre un sector del material cartográfico (Figura 1) se dispuso una cuadrícula con celdas de 2km de lado, acorde con las divisiones ya existentes en la serie más antigua de cartas. Cada celda se dividió en cuatro cuadrantes de 1km de lado. A su vez, cada cuadrante se dividió en 16 unidades de 0,0625km2. El sector seleccionado está limitado por el cauce principal (Paraná Guazú) y por la línea media entre éste y el brazo secundario en este tramo (Paraná Pavón). De esta manera se incluyeron en el análisis sectores del valle que se encuentran a diferente distancia con respecto al cauce principal. El total de la superficie seleccionada del sistema fue de 212km2.
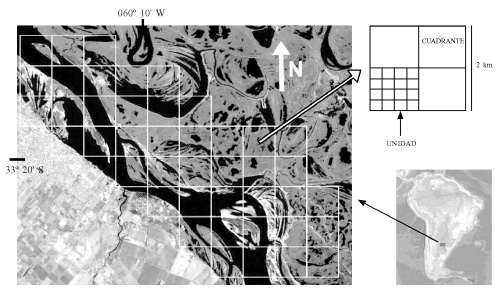
Figura 1. Imagen satelital del área y esquema de la metodología utilizada.
Tasa de cambio geomorfológico. Utilizando la cartografía mencionada, se clasificó a los ambientes en tres tipos distintos, que se denominaron "isla" (cualquier clase de terreno emergido), "agua" y "bancos" (distinguibles en las cartas con un punteado y en las imágenes con color blanco).
En el sector de las cartas topográficas cuadriculado en la Figura 1, se identificó cada unidad y se registró el tipo de ambiente preponderante. Se consideró preponderante al ambiente que ocupaba más de la mitad de la superficie de la unidad. Para el centro de cada cuadrante se determinó la distancia mínima existente hasta la línea media del cauce principal (se consideró el brazo Paraná Guazú). El procedimiento de clasificación se repitió para el mismo sector de las imágenes satelitales.
Luego se sumó el número de unidades en que hubo transición de un tipo de ambiente a otro (Margalef, 1991), obteniéndose así tasas de cambio para los procesos de erosión y sedimentación, a partir de las unidades de "isla" y "bancos" que se transformaron en "agua", y de "agua" transformados en "isla" o "banco", respectivamente. Finalmente se obtuvieron valores de superficie al multiplicar los resultados en unidades por la superficie de una unidad (0,0625km2). Por ejemplo, la tasa de cambio isla ® agua se calculó como

De igual modo, el total de las unidades con transiciones de cualquier tipo se denominó tasa global de cambio geomorfológico.
Abundancia y disposición espacial de los bosques y asociación con las tasas de cambio geomorfológico. Se determinó la superficie de llanura aluvial ocupada por bosques de acuerdo a las cartas topográficas nombradas, en el mismo sector cuadriculado de la Figura 1 y siguiendo la metodología ya descripta para la obtención de las tasas de cambio geomorfológico. En este caso se consideró el número de unidades de 0,0625km2 con más de la mitad de su área ocupadas por un bosque.
Por otra parte, utilizando índices de asociación que relacionan la ocurrencia de cambio geomorfológico con la presencia de bosques en cada cuadrante del sector cuadriculado (Tabla I) se analizó la asociación existente entre ambas.

Se utilizaron tres índices cualitativos: I1, de Sokal y Michener (1958) que involucra las dobles ausencias; I2, de Jaccard (1908), a los efectos de comparar los resultados en caso de no considerar las dobles ausencias; e I3 o PHI (Guilford, 1942), que varía de -1 a 1 indicando asociaciones positivas y negativas. El significado de las letras en las fórmulas de los índices se encuentra en la Tabla I.
Erosión en bosques establecidos. Debido a lo extenso de los plazos necesarios para estudiar en el campo los procesos de erosión, se decidió estimar su ocurrencia a partir de los datos de tasa de erosión, obtenidos al estudiar las tasas de cambio geomorfológico. Para estimar cuantitativamente el impacto de los procesos erosivos sobre los bosques ya establecidos, se determinó el número de cuadrantes en los que se registraron simultáneamente bosques en la cartografía más antigua y se detectaron fenómenos de erosión al comparar la cartografía más antigua con la más moderna,
A pesar de que no se puede asegurar que la totalidad de los árboles hayan desaparecido efectivamente por efecto de la erosión (podrían haber muerto por otras causas antes de que se erosione su albardón), lo que se pretende demostrar es que las disposiciones espaciales de bosques y procesos de erosión están asociadas.
Registros de campo
Nivel topográfico y tamaño de los árboles. Se intentó determinar la existencia de una relación entre nivel topográfico y edad de los árboles. Para ello se midieron los diámetros a la altura del pecho (DAP) en los ejemplares adultos o el diámetro basal en los renovales y los niveles topográficos de 139 árboles de 11 bosques ubicados en islas de cauce del Paraná inferior en la zona cercana a San Nicolás. Para esto, se utilizaron jalones de 1,20m y una manguera transparente llena de agua, a modo de nivel (principio de los vasos comunicantes). Todos los niveles topográficos están referidos al cero del hidrómetro de San Nicolás (ubicado a 2,30m sobre el nivel del mar), para permitir su correcta comparación.
Proporción de renovales. Para hallar la proporción de renovales dentro de bosques pioneros ya establecidos, se realizaron transecciones perpendiculares a la línea de ribera, atravesando el albardón costero, en cinco bosques de la misma isla. Sobre cada transección, cada 10m, se consideró un punto a partir del cual se determinaron cuatro cuadrantes. En cada cuadrante se registró la presencia del individuo de sauce, renoval o adulto, más cercano al punto.
Resultados
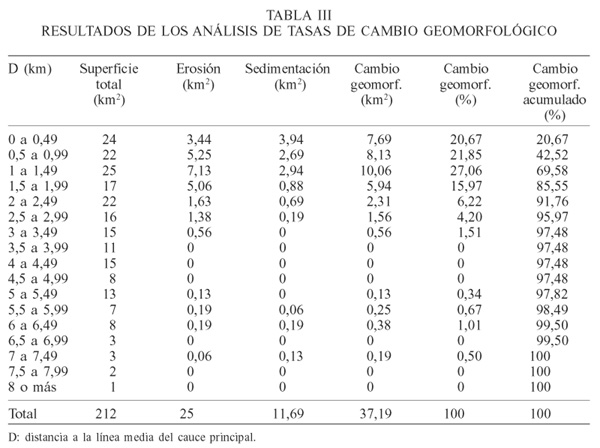
Tasa de cambio geomorfológico. La tasa global o total con cambio geomorfológico hallada en el período de tiempo transcurrido entre ambas series de cartas (37 y 40 años) fue de 595 unidades, lo que conformó una superficie modificada de 37,19km2 (17,54% del total de 212km2).
Se observó una predominancia global de los fenómenos erosivos sobre los de sedimentación: 25,00km2 (11,42%) frente a 12,19km2 (5,75%), respectivamente. También se apreció la pequeña proporción de aparición de bancos (0,50km2, es decir 0,16%) existente en ese momento con respecto al total.
En general se observó una mayor tasa de cambio en los sectores cercanos al cauce principal, mostrando la potencia modeladora del mismo.
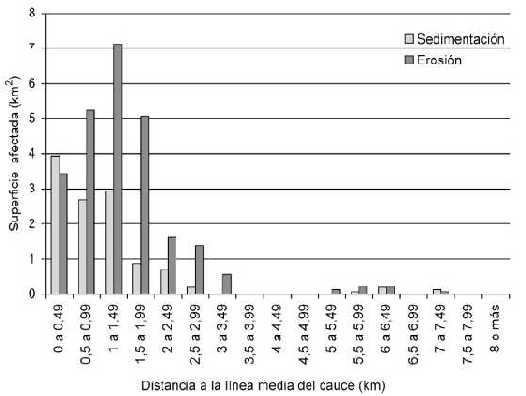
En los primeros 0,5km, los procesos erosivos y sedimentarios muestran valores semejantes, y algo menores a los inmediatamente más alejados del cauce. En esta distancia ocurre la máxima sedimentación en el sector estudiado. A mayor distancia de la línea media del cauce, los fenómenos erosivos son de mayor amplitud que los sedimentarios.
Los procesos erosivos muestran valores muy altos hasta una distancia de 2km de la línea media del cauce principal, con máximo entre 1 y 1,5km, si bien ocurren hasta aproximadamente los 3,5km.
En la Figura 2 se observa la ubicación de las zonas con mayor tasa de cambio geomorfológico, con respecto a su distancia al centro del cauce principal. Dentro de los 2km más cercanos a la línea media del cauce se produce el 85,55% del cambio geomorfológico, y dentro de los 3km el porcentaje alcanza el 95,97% del cambio.
Figura 2. Comparación de las superficies que sufrieron sedimentación y erosión en cada rango de distancias referidas al centro del cauce.
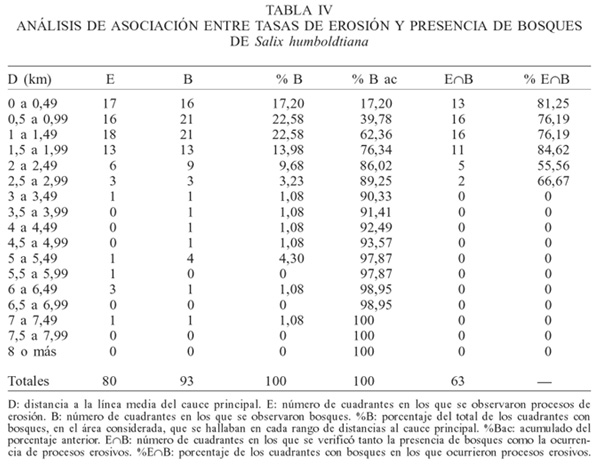
Abundancia y disposición espacial de los bosques y asociación con las tasas de cambio. Sobre una superficie de 212km2, los bosques señalados ocuparon 35,19km2, es decir 16,60% del sector seleccionado del sistema. El 89,25% de los bosques está ubicado en los 3km más cercanos al cauce principal (Tabla IV) mientras que, salvo pequeños parches a orillas de algún brazo interno, no se hallan bosques en los sitios alejados del cauce principal.
Los valores de los índices utilizados para cuantificar la asociación existente entre la presencia de bosques y la ocurrencia de cambios geomorfológicos fueron 0,821 (I1) 0,667 (I2) y 0,638 (I3). En los tres casos, existió una correlación positiva entre la presencia de bosques y la tasa de cambio geomorfológico hallada.
Al ubicar ambas variables ordenadas en un gradiente de distancia creciente desde el centro del cauce principal hasta los puntos de la llanura aluvial más alejados de los mismos (Figura 3), se puede observar gráficamente la correlación obtenida con los índices de asociación.
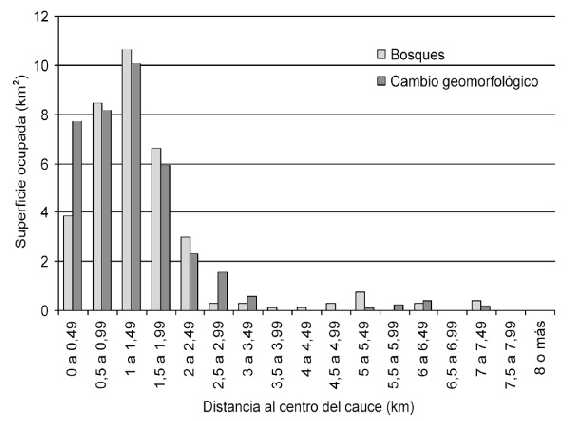
Figura 3. Superficies que sufrieron cambio geomorfológico y áreas ocupadas por bosques, en cada rango de distancias referidas al centro del cauce.
Nivel topográfico y tamaño (DAP). La Figura 4 muestra el DAP (usado como un estimador de la edad) y el nivel topográfico en que se hallaba cada árbol. Se indican, además, los porcentajes de tiempo de inundación del suelo en cada nivel. Se utilizó la serie de tiempo de los niveles hidrométricos diarios del puerto de San Nicolás correspondiente al período 1974-1995.
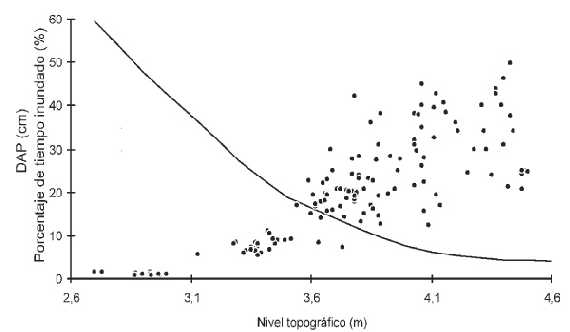
Figura 4. Diámetro a la altura del pecho (DAP) y nivel topográfico ocupado por los árboles registrados. La línea negra continua indica el porcentaje de tiempo que permaneció inundado cada nivel hidrométrico en el período de tiempo considerado (1974-1995).
Al analizar bosques ubicados en niveles topográficos mayores se evidenció un aumento gradual del DAP. El valor de la regresión hallado (r2) fue de 0,666.
Renovales. En la Tabla II se observa una escasa presencia de renovales en el interior de bosques maduros (4,35% de los ejemplares). En ningún caso se observó la agrupación típica de renovales, dispuestos en parches de alta densidad, sino que se encontraron ejemplares pequeños y aislados. Es común hallar retoños de tipo vegetativo, como rebrotes de árboles mayores quebrados o sepultados por crecientes anteriores, que han sido descriptos por Worbes (1997) para poblaciones del Amazonas.

Erosión en bosques establecidos. En la Tabla IV puede apreciarse que 67,74% del total de los cuadrantes (=63/93.100) que contenían bosques en las cartas topográficas sufrió también procesos de erosión, de acuerdo a las tasas obtenidas a partir de la comparación con las imágenes satelitales más modernas. De los bosques situados a menos de 3km de la línea media del cauce (que hacen el 89,3% del total), el porcentaje de cuadrantes coincidentes asciende al 75,9%.
Discusión
Las tasas de cambio halladas para la zona implican una renovación importante de las geoformas más dinámicas (islas de cauce y sectores más cercanos a los cauces principales). El porcentaje del sistema modificado resultó de 17,54% del total en el período de tiempo transcurrido, con un 95% de estas modificaciones ocurridas en los 3km más cercanos a los cauces, lo que indica (como se observa además en las Figuras 2 y 3) que las fracciones de llanura aluvial más cercanas a los cauces principales presentan la mayor tasa de cambio geomorfológico; a su vez, se ha mostrado que cerca del 90% de los bosques de sauce ocupan las fracciones más dinámicas de llanura aluvial.
Estos resultados se condicen con la variación transversal hallada por Marchese y Ezcurra de Drago (1992) al analizar la distribución del bentos en ambientes de la planicie, desde los cauces principales a las zonas centrales de la llanura de inundación del río Paraná. Al acercarse a los sectores con mayor influencia de los cauces principales, estas autoras señalan valores crecientes de velocidad de la corriente, profundidad, proporción de arena y contenido de oxígeno, que pueden asociarse cualitativamente con una dinámica hidrosedimentológica mayor.
Los sauces pueden hallarse en un amplio rango de valores de nivel topográfico: desde los 2,5m hasta los 4,5m con respecto al cero del hidrómetro del puerto de San Nicolás (Figura 4), lo que implica que pueden soportar una amplia gama de condiciones de inundabilidad (<4% a >60% del tiempo con el suelo inundado). Si se considera la edad de los ejemplares, puede observarse además que individuos de mayor tamaño ocupan niveles topográficos gradualmente más elevados, lo que verifica la existencia de una relación directa entre el gradiente de los niveles topográficos de los albardones y las edades de los bosques que se encuentran sobre ellos.
Los mecanismos físicos y biológicos que elevan gradualmente el nivel topográfico son conocidos: disminución de la velocidad de la corriente, la consiguiente pérdida de la capacidad de transporte de sedimentos (Derruau, 1991) y "rugosidad biológica" (atenuación del escurrimiento, Depetris et al., 1992). Avanzado el proceso, puede observarse un bosque maduro, sobre un albardón con horizontes superficiales arcillosos y capas arenosas más profundas (Lewis y Franceschi, 1979).
La distribución de las poblaciones de sauce en la llanura aluvial está limitada por su nicho de generación (Grubb, 1977), definido aquí como el conjunto de las condiciones compatibles con el establecimiento de brinzales. Sólo en los bancos de reciente formación es donde se conjugan las condiciones necesarias para permitir la germinación: sustrato húmedo1 y poco consolidado, con buena carga de nutrientes que permita el rápido crecimiento de las plántulas, con densidad suficiente de semillas (arrastradas por el agua y el viento) y alta disponibilidad de luz (Neiff et al., 1985; Niiyama, 1990).
1 En muchas especies de Salicáceas (Niiyama, 1990) se ha comprobado que la germinación sólo ocurre en condiciones de humedad. Por ejemplo, las semillas de S. nigra sólo germinan si se dan condiciones continuas de humedad durante todo el proceso (McLeod y McPherson, 1973). En varias especies de Salix de Alaska se ha demostrado que las semillas pueden germinar incluso bajo el agua (Krasny et al., 1988). En S. nigra, el órgano primario de absorción de agua de las plántulas es un círculo de pelos situados en la unión de la raíz y el hipocótilo; durante la primera semana de crecimiento, la raíz no se desarrolla lo suficiente; como estos pelos no penetran en el suelo, es necesario que exista permanentemente un grado importante de humedad superficial (McLeod y McPherson, 1973).
Debe tenerse en cuenta que la aparición de bancos adecuados para la colonización no es un fenómeno continuo. El pequeño porcentaje de nuevos bancos (0,16% de los 212km2 de superficie total) indica lo episódico de su formación. Deben darse además variaciones adecuadas del nivel hidrométrico que permitan la germinación de las semillas, que no inunden las plántulas demasiado rápido, por demasiado tiempo o, por el contrario, que provoquen un tiempo largo de sequía. McLeod y McPherson (1973) señalan que el establecimiento de bosques de S. nigra es un evento relativamente raro, debido a los especiales requerimientos de sus semillas para germinar. En poblaciones norteamericanas de S. lasiolepis, por ejemplo, son suficientes de 7 a 9 días con el banco emergido y sin lluvia para provocar gran mortalidad por déficit de agua (Sacchi y Price, 1992). Lo crítico del nicho de generación parece darse también para otra especie arbórea pionera como el aliso (Tessaria integrifolia), que en el río Amazonas presenta establecimientos de bosques cada cuatro años en promedio (Salo et al., 1986).
Burkart (1957) menciona, al tratar los sauzales de S. humboldtiana del delta del Paraná, que se encuentran renovales "en bancos e islas nuevas, donde nace en densas masas, como en almácigo", a la vez que señala la exigüidad de la reproducción en islas ya pobladas. En la región, Lewis y Franceschi (1979) indican que cada banco es colonizado una sola vez y que todos los individuos son coetáneos. La presente experiencia de conteo de renovales confirma que, en la zona del presente estudio, no se da prácticamente renovación en el interior de los bosques.
La longevidad de S. humboldtiana ha sido determinada, para poblaciones del Amazonas (isla de Marchantaria, Brasil) en 20 años, con ejemplares de diámetro a la altura del pecho de 32cm como máximo (Worbes, 1997). En un tramo del río Paraná Superior, el DAP osciló mayormente entre 20 y 30cm (Neiff, 1986). El máximo diámetro hallado en el presente trabajo para S. humboldtiana en los bosques pioneros del Paraná Inferior fue de 49,7cm. No se hallaron trabajos regionales que relacionen DAP y edad para esta especie, pero puede inferirse que estos ejemplares superan ampliamente la edad estimada tanto para el Amazonas como para tramos superiores del río Paraná.
En todos los bosques de sauces estudiados aquí, ubicados en albardones costeros, se observaron los efectos de la erosión. En los sectores con corrientes fuertes, se halló habitualmente una línea de árboles caídos y parcialmente sumergidos; en los sitios con corrientes de menor intensidad, con márgenes arenosas con declive suave, se observan ejemplares con el sistema radicular en forma de plato emergiendo a veces más de 1 m del nivel actual del suelo. Es decir que al menos una parte del porcentaje de bosques en los que a la vez se verificó erosión (Tabla IV) ha desaparecido efectivamente por efecto de ésta.
La importancia de la erosión como factor de control de las poblaciones se refleja en el porcentaje de la superficie del sistema que, estando cubierta de bosques sufrió procesos erosivos en un período de alrededor de 40 años.
Es posible que este proceso sea diferente en tramos superiores del río, donde aparecen las selvas mixtas de inundación, que son unidades de vegetación adaptadas a niveles topográficos más elevados y poseen una mayor variedad de especies, entre las que puede hallarse S. humboldtiana, pero con carácter recesivo (Neiff, 1986). De esta manera el factor de control de las poblaciones pasaría de ser biótico (competencia interespecífica en una sucesión) en el tramo superior a ser mayormente abiótico (erosión) en el tramo inferior.
Los puntos anteriores pueden integrarse en un esquema (Figura 5) en que se resaltan la retroalimentación entre el ambiente (controlado por el pulso) y los bosques, en sus distintas edades: dado el carácter dinámico y temporario de los sitios en que se encuentran, los bosques no pueden permanecer en el largo plazo en un mismo sitio; las semillas crecen en cambio en bancos generados recientemente por la dinámica del pulso de inundación; estos bancos se transformarán luego -por sedimentación- en albardones, en un proceso favorecido por la propia presencia de los brinzales.
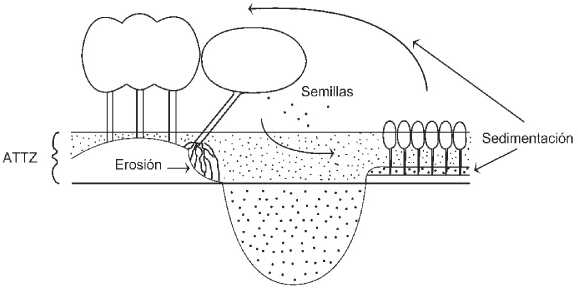
Figura 5. Esquema de la integración de Salix humboldtiana al pulso de inundación - sequía. ATTZ: zona de transición acuática-terrestre.
La reproducción y crecimiento de los bosques de sauce criollo están de tal manera enlazados con el pulso de inundación-sequía, que sólo se hallan donde éste se manifiesta.
Margalef (1968) señala que el plancton utiliza las perturbaciones del flujo para evitar su exportación hacia niveles inferiores de los sistemas lóticos. Tablado y Oldani (1984) sugieren que muchos peces de importancia comercial realizan migraciones ascendentes reproductivas, con el fin de asegurar la permanencia de sus juveniles dentro del sistema fluvial. De manera semejante, la colonización de los bancos por los brinzales asegura la permanencia de las poblaciones de sauces, ya que si los bosques se perpetuasen en los mismos sitios, en algunas decenas de años la elevada tasa de cambio geomorfológico los haría desaparecer junto con los albardones en los que se encuentran.
El hecho de hallar adaptaciones semejantes en poblaciones tan distintas entre sí como plancton, árboles y peces confirma la predominancia del control de los factores abióticos (contenidos en el concepto del pulso) sobre la biota de los sistemas fluviales.
AGRADECIMIENTOS
A Juan José Neiff por el apoyo y la dirección brindados desinteresadamente. A Mario Wagner y Beatriz Giacosa por su colaboración en innumerables horas de trabajos de campo y por sus comentarios y consejos.
REFERENCIAS
1 Bonetto AA, Wais IR (1989) Las grandes planicies aluviales del sistema potámico Paraná-Paraguay. Características estructurales, funcionales, preservación y manejo. Mus. Arg. C. Nat. "B. Rivadavia". Hidrobiol., 6(4): 3-29. [ Links ]
2 Burkart A (1957) Ojeada sinóptica sobre la vegetación del Delta del río Paraná. Darwiniana 11: 457-561. [ Links ]
3 Cabrera AL, Zardini EM (1993) Manual de la flora de los alrededores de Buenos Aires. ACME. Buenos Aires. 755 pp. [ Links ]
4 Depetris PJ, Kempe S (1990) The impact of the El Niño 1982 event on the Paraná River, its discharge and carbon transport. Palaeogeog, Palaeoclim, Palaeoecol. 89: 239-244. [ Links ]
5 Depetris C, Orfeo O, Neiff JJ (1992) Atenuación del escurrimiento fluvial por bosques de Tessaria integrifolia. Ambiente subtropical 2: 33-43. [ Links ]
6 Derruau M (1991) Geomorfología. Ariel Geografía. Barcelona. 499 pp. [ Links ]
7 Franceschi EA, Lewis JP (1979) Notas sobre la vegetación del valle santafesino del río Paraná (República Argentina). Ecosur 6(11): 55-82. [ Links ]
8 Guilford JP (1942) Fundamental Statistics in Psychology and Education. Nueva York, McGraw-Hill. 333 pp. [ Links ]
9 Grubb PJ (1977) The maintenance of species richness in plant communities: the importance of the regeneration niche. Biol. Rev. 52: 107-145. [ Links ]
10 Jaccard P (1908) Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat. 44: 223-270. [ Links ]
11 Junk WJ, Bayley PB, Sparks RE (1989) The flood-pulse concept in river-floodplain systems. En Dodge DP (Ed.) Proceedings of the International Large River Symposium, Can. Spec. Publ. Fish Aquat. Sci. 106: 110-127. [ Links ]
12 Krasny ME, Vogt KA, Zasada JC (1988) Establishment of four Salicaceae species on river bars in interior Alaska. Holartic Ecol. 11: 210-219. [ Links ]
13 Lewis JP, Franceschi EA (1979) Notas sobre la dinámica de la vegetación del valle del río Paraná. Ecosur 6(12): 145-163. [ Links ]
14 Marchese M, Ezcurra de Drago I (1992) Benthos of the lotic environments in the middle Paraná River system: transverse zonation. Hydrobiologia 237: 1-13. [ Links ]
15 Margalef R (1968) Perspectivas de la teoría ecológica. Blume. Barcelona. 110 pp. [ Links ]
16 Margalef R (1991) Teoría de los sistemas ecológicos. Publicacions Universitat de Barcelona. Barcelona. 290 pp. [ Links ]
17 McLeod KW, McPherson JK (1973) Factors limiting the distribution of Salix nigra. Bull. Torrey Bot. Club 100 (2): 102-110. [ Links ]
18 Neiff JJ (1986) Las grandes unidades de vegetación y ambiente insular del río Paraná en el tramo Candelaria - Itá-Ibaté. Rev. Asoc. Cs. Nat. Litoral 17: 7-30. [ Links ]
19 Neiff JJ (1990) Ideas para la interpretación ecológica del Paraná. Interciencia 15: 424-441. [ Links ]
20 Neiff JJ, Reboratti HJ, Gorleri MC, Basualdo M (1985) Impacto de las crecientes extraordinarias sobre los bosques fluviales del Bajo Paraguay. Bol. Com. Espec. Río Bermejo. Cámara de Diputados de la Nación (Buenos Aires) 4: 13-30. [ Links ]
21 Niiyama K (1990) The Role of Seed Dispersal and Seedling Traits in Colonization and Coexistence of Salix Species in a Seasonally Flooded Habitat. Ecol. Res. 5: 317-331. [ Links ]
22 Sacchi CF, Price PW (1992) The relative roles of abiotic and biotic factors in seedling demography of arroyo willow. Am. J. Botany 79: 395-405. [ Links ]
23 Salo J, Kalliola R, Hakkinet I, Makinen Y, Niemela P, Puhakka M (1986) River dynamics and the diversity of Amazon lowland forest. Nature 322: 254-258. [ Links ]
24 Sokal RR, Michener CD (1958) A statistical method for evaluating systematic relationships. Univ. Kansas Sci. Bull. 38: 1409-1438. [ Links ]
25 Tablado A, Oldani N (1984) Consideraciones generales sobre las migraciones de peces en el río Paraná. Bol. Asoc. Cienc. Nat. Litoral 4(3): 31-34. [ Links ]
26 Vanotte RL, Minshall GW, Cummins KW, Seddell JR, Cushing CE (1980) The River Continuum Concept. Can. J. Fish. Aquat. Sci. 37: 130-137. [ Links ]
27 Worbes M (1997) A study of forest dynamics and wood production in flooded forests in the Amazon basin, using ring trees. Nat. Resources & Developm., 45/46: 54-75. [ Links ]
