











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.32 n.1 Caracas ene. 2007
Las micorrizas arbusculares como alternativa para una agricultura sustentable en areas tropicales
Gisela Cuenca, Alicia Cáceres, Giovanny Oirdobro, Zamira Hasmy y Carlos Urdaneta
Gisela Cuenca Licenciada en Biología, Universidad Central de Venezuela, (UCV). M.Sc. y Ph.Sc. en Ecología, Instituto Venezolano de Investigaciones Científicas (IVIC). Investigador Asociado Titular, IVIC. Dirección: Laboratorio de Ecología de Suelos, Centro de Ecología, IVIC. Apartado Postal 21827, Caracas 1020-A, Venezuela. e-mail: gcuenca@ivic.ve
Alicia Cáceres Licenciada en Biología, UCV, Venezuela. M.Sc. en Ecología, IVIC. Doctor en Ciencias, UCV. Profesor, UCV, Venezuela. e-mail: alicia2001@gmail.com
Giovanny Oirdobro Ingeniero Agrónomo, Universidad Centro Occidental Lisandro Alvarado, Venezuela (UCLA). Estudiante de Maestría, IVIC.
Zamira Hasmy Licenciada en Biología, Universidad del Zulia, Venezuela. Profesional Asociado a la Investigación, Centro de Ecología, IVIC, Venezuela.
Carlos Urdaneta Técnico Superior Universitario en Producción Agropecuaria, Universidad Nacional Experimental del Sur del Lago, Venezuela. Técnico Asociado a la Investigación, Centro de Ecología, IVIC, Venezuela.
RESUMEN
Las micorrizas arbusculares (MA) son asociaciones ecológicamente mutualistas entre hongos del Phyllum Glomeromycota y la inmensa mayoría de las plantas, pudiendo ser una herramienta muy útil para una agricultura sustentable. Entre sus efectos beneficiosos están: mayor absorción de elementos poco móviles como P, Cu y Zn; protección contra patógenos; mayor resistencia a la sequía; y contribución a la formación de la estructura del suelo. Se presentan resultados preliminares de un proyecto para producir inoculantes comerciales de MA a ser utilizados en suelos ácidos, y las potencialidades de su aplicación en áreas tropicales, particularmente en Venezuela. Se muestran pasos cruciales en la producción de inoculantes y sus ventajas en cultivos de lechuga y yuca. Se produjeron inoculantes de Glomus manihotis, Acaulospora lacunosa, Entrophospora colombiana, Scutellospora fulgida y S. heterogama, que fueron probados tanto en el invernadero como en el campo. Los mayores pesos frescos de lechuga se obtuvieron con G. manihotis y S. fulgida, mientras que S. heterogama produjo un resultado pobre. Con yuca, la mayor producción de tubérculos se obtuvo con G. manihotis y la lograda con A. lacunosa no se distinguió del control. Los resultados indican que aunque la simbiosis carece de especificidad taxonómica, las especies de MA no son equivalentes. Solo con un manejo ético y responsable que considere las condiciones ecológicas del cultivo que se va a inocular, se podrá utilizar exitosamente esta simbiosis y aumentar la productividad, minimizando daños ambientales.
Arbuscular mycorrhizae as an alternative for a sustainable agriculture in tropical areas
SUMMARY
Arbuscular mycorrhizae (AM) are ecologically mutualistic associations between Glomeromycota Phyllum fungi and the majority of plants, and could be very useful to achieve a sustainable agriculture. Among the benefits obtained are: increase in absorption of nutrients that are sparingly mobile in soils, such as P, Cu and Zn; better resistance to pathogens; water shortage endurance; and contribution to the formation of soil structure. Preliminary results of a project on commercial production of AM inocula to be used in acidic soils are presented, and the potential of AM for their application in tropical areas, particularly in Venezuela, is discussed. Some crucial steps in the AM-inoculum production are shown, as well as advantages of their application for lettuce and cassava production. Inocula of Glomus manihotis, Acaulospora lacunosa, Entrophospora colombiana, Scutellospora fulgida and S. heterogama were produced, and their effect tested both in the greenhouse and field. Higher values of lettuce dry weight were obtained with G. manihotis and S. fulgida, while S. heterogama led to poor results. With cassava the highest tuber production was obtained with G. manihotis, and with A. lacunosa it was indistinguishable from controls. Results indicate that although mycorrhizal symbiosis lacks taxonomic specificity, not all the AM are equivalent. An ethical and responsible management of AM, that takes into account the ecological conditions of the crop to be inoculated, is necessary for a successful application of this technology, so as to increase crop productivity with minimal environmental damage.
As micorrizas arbusculares como alternativa para uma agricultura sustentável em áreas tropicais
RESUMO
As micorrizas arbusculares (MA) são associações ecologicamente mutualistas entre fungos do Phyllum Glomeromycota e a imensa maioria das plantas, podendo ser uma ferramenta muito útil para uma agricultura sustentável. Entre seus efeitos beneficiosos estão: maior absorção de elementos pouco móveis como P, Cu e Zn; proteção contra patógenos; maior resistência a seca; e contribuição à formação da estrutura edáfica. Apresentam-se resultados preliminares de um projeto para produção comercial de inoculantes de MA a ser utilizados em solos ácidos, e as potencialidades de sua aplicação em áreas tropicais, particularmente na Venezuela. Mostram-se passos cruciais na produção de inoculantes e suas vantagens em cultivos de alface e mandioca. Se produziram inoculantes de Glomus manihotis, Acaulospora lacunosa, Entrophospora colombiana, Scutellospora fulgida e S. heterogama, que foram provados em cultivos de alface em invernadeiro e de mandioca no campo. Os maiores pesos frescos de alface se obtiveram com G. manihotis e S. fulgida, enquanto que S. heterogama produziu um resultado pobre. Com mandioca, a maior produção de tubérculos se obteve com G. manihotis e a lograda com A. lacunosa não se distinguiu do controle. Os resultados indicam que ainda que a simbiose carece de especificidade taxonômica, as espécies de MA não são equivalentes. Somente com um manejo ético e responsável que considere as condições ecológicas para a aplicação se poderá utilizar exitosamente esta simbiose e aumentar a produtividade, minimizando danos ambientais.
PALABRAS CLAVE / Agricultura Sustentable / Ahorro de Fertilizantes / Fertilización / Lechuga / Micorrizas / Yuca /
Recibido: 23/08/2006. Modificado: 23/11/2006. Aceptado: 27/11/2006.
Las micorrizas arbusculares (MA) son asociaciones ecológicamente mutualistas que se establecen entre un selecto grupo de hongos (Glomeromycota) y la gran mayoría de las plantas. Aproximadamente un 80% de las familias de plantas existentes tienen la potencialidad de formar este tipo de asociación (Trappe, 1987). Las MA son el tipo de micorrizas que forman la mayoría de las plantas de interés agrícola. En dicha asociación, el hongo forma arbúsculos que son las estructuras donde se realiza el intercambio de carbono y fósforo entre el hongo y la planta. Algunos hongos micorrízicos forman vesículas en el micelio interno, las cuales son estructuras de reserva del hongo.
Actualmente son bien conocidos los efectos beneficiosos de las MA, los cuales comprenden la mayor absorción de elementos poco móviles en el suelo como el fósforo, cobre y zinc por parte de las plantas micorrizadas en comparación con las no micorrizadas (Smith y Read, 1997). Además, gracias al uso más eficiente que hacen las plantas micorrizadas de los nutrientes del suelo, permiten ahorrar fertilizantes químicos y reducir por consiguiente los problemas de contaminación que el uso excesivo de fertilizantes conlleva.
Por otra parte, las plantas micorrizadas son capaces de hacer un mejor uso de los fertilizantes orgánicos, bien sea debido a la producción de fosfatasas por parte de los hongos mismos (Dodd et al., 1987; Joner y Johansen, 2000) o bien gracias a la asociación existente entre las hifas de las MA y los microorganismos que participan en la mineralización de la materia orgánica (Azcón-Aguilar y Barea, 1992). Además, existen evidencias de que las MA protegen a las plantas del ataque de patógenos (Newsham et al., 1995) y del déficit hídrico (Ruiz-Lozano y Azcón, 1995). Finalmente, también son conocidos los efectos de las MA en la formación de la estructura del suelo, a través de su papel en la constitución de agregados estables al agua, en los que el micelio externo de las MA tiene una notable participación (Miller y Jastrow, 2000), así como a través de la producción de una glicoproteína llamada Glomalina, la cual por sus características químicas favorece la agregación de las partículas de suelo (Rillig, 2004).
El manejo agrícola conlleva varios problemas ambientales entre los que destacan la ya mencionada excesiva aplicación de fertilizantes químicos que terminan contaminando los cuerpos de agua y causando su eutroficación. La adición de fertilizantes sin el análisis previo de las condiciones del suelo, además, puede conducir a un desbalance iónico de los mismos, con los consiguientes problemas para las plantas que viven en dicho suelo y sus micorrizas asociadas (Herrera-Peraza et al., 1984).
En las últimas décadas se ha intentado cambiar en el ámbito global los paradigmas de la producción agrícola que implicaban el uso intensivo de energía, maquinaria y sustancias químicas (la llamada revolución verde) por un nuevo concepto, el de la agricultura sustentable. Según ese nuevo paradigma la agricultura sustentable es un "sistema integrado de prácticas de producción vegetal y animal que a largo plazo debe a) satisfacer las necesidades humanas de fibra y alimentos b) mejorar la calidad ambiental y la base de recursos naturales de los cuales depende la economía agrícola c) hacer un uso eficiente de los recursos no renovables d) sostener la viabilidad económica de las actividades agrícolas y e) aumentar la calidad de vida de los agricultores y de la sociedad como un todo" (Jeffries y Barea, 2001).
De tal definición puede deducirse que el uso de las micorrizas podría ser una herramienta útil para acercarnos a una agricultura sustentable. De hecho, existe una creciente conciencia ambiental gracias a la cual está aumentando la demanda de productos con certificación "orgánica", es decir aquellos con la garantía de que durante su fase de cultivo y procesamiento no se han utilizado sustancias químicas artificiales. Por otra parte, en la América tropical en general y en particular en Venezuela existen todavía fuentes de aguas prístinas que es deseable conservar, aunque en algunas regiones ya se confrontan serios problemas de eutroficación como sucede en los lagos de Valencia y de Maracaibo (Infante et al., 1979; González, 2004). En muchos países latinoamericanos la agricultura carece actualmente de una eficiente extensión técnica, lo que ha conducido al uso indiscriminado de agroquímicos y fertilizantes. Ello hace necesario desarrollar y divulgar estrategias de manejo agrícola que permitan minimizar estos problemas.
En este trabajo se presentan los resultados preliminares de un proyecto que pretende la producción comercial de inoculantes de micorrizas arbusculares para ser utilizados en la agricultura de suelos ácidos y las potencialidades que ofrece esta simbiosis para ser utilizada en Venezuela. Ello es particularmente importante porque al tratarse de un país tropical, tiene extensas áreas de suelos muy lixiviados, ácidos y pobres en nutrientes especialmente en fósforo (Paolini, 1988), donde las micorrizas pueden ser aplicadas con éxito. El objetivo de este trabajo, el cual constituye uno de los primeros intentos de producir inoculantes de HMA en Venezuela, es señalar algunos pasos cruciales en la producción de dichos inóculos y demostrar las ventajas del uso de los mismos en sistemas usuales en la región, tales como un cultivo de lechuga y uno de yuca.
Materiales y Métodos
Preselección de los hongos a reproducir
El Laboratorio de Ecología de Suelos del Instituto Venezolano de Investigaciones Científicas (IVIC) cuenta con una colección viva de Glomeromycota, algunos de ellos aislados localmente y otros provenientes de colecciones internacionales. Dichos cultivos han sido mantenidos por largo tiempo en macetas de 4l de capacidad llenos con suelo estéril proveniente del bosque nublado local. La planta hospedera utilizada es Vigna luteola, una maleza que crece en los alrededores del instituto que es muy micorrízica (Hernández et al., 2000).
Para seleccionar los cultivos con mayor número de esporas y más apropiados para reproducir a una escala comercial, se revisaron 47 cultivos de la colección y de cada uno se aislaron las esporas presentes, utilizando el método del tamizado húmedo y decantado seguido de centrifugación en sacarosa según Sieverding (1991). Bajo el estereoscopio se contaron las esporas presentes por cada 100g de sustrato y se descartó la presencia de nemátodos u otros agentes patógenos. Se seleccionaron aquellos cultivos que poseían el mayor número de esporas saludables e intactas y en los que no se detectó agente patógeno alguno.
Preparación de los inoculantes
Los hongos micorrízicos arbusculares (HMA) no son capaces de crecer en ausencia de una planta hospedera (Azcón-Aguilar y Barea, 1999). La manera más usual de cultivarlos consiste en colocar sus esporas en un suelo previamente esterilizado y sembrar luego la semilla pre-germinada de una planta capaz de formar micorrizas (Brundrett et al., 1996). Al cabo de 3-4 meses el suelo y las raicillas de la planta hospedera pueden utilizarse como un inoculante de HMA.
En este trabajo se sustituyó el suelo por un sustrato más liviano para producir inoculantes que resultasen más fáciles de transportar y distribuir para su comercialización. Trabajos previos permitieron seleccionar a las arcillas expansivas como el sustrato más adecuado para la reproducción de los HMA (Giovanny Oirdobro, datos no publicados). El origen exacto de dicha arcilla no será revelado en este trabajo en vista de la próxima necesidad de patentar el producto.
La arcilla se diluyó con arena o con cáscara de arroz, para seleccionar la mezcla y dilución que produjese el inoculante de la mayor calidad. Para ello se realizaron los siguientes ensayos:
Optimización de la proporción arcilla/arena silícea. En este ensayo se diluyó la arcilla con distintas dosis de arena silícea a saber: 1/1, 1/2, 1/4 y 1/8 (v/v). De tales diluciones se prepararon dos tratamientos: 1) Control no micorrizado y 2) Inoculación con una mezcla de Glomus manihotis y Scutellospora fulgida. El inóculo contenía 16359 esporas de S. fulgida·100g-1 y 4349 esporas de G. manihotis·100g-1.
De cada uno de estos tratamientos se hicieron triplicados y se utilizó como planta hospedera a Vigna luteola. Las plantas se sembraron en recipientes plásticos (tubetes) de 350ml de capacidad. El inóculo se colocó por debajo del hoyo de siembra en la cantidad de 30g por planta. En total el ensayo constó de 24 plantas y se mantuvo en el invernadero por 2 meses, al cabo de los cuales se cosecharon las plantas para determinar su peso seco y la presencia de micorrizas en las raíces. El peso seco tanto de la parte aérea como de la raíz se determinó después de secar el material a 60ºC hasta peso constante. La presencia de micorrizas se cuantificó en las raíces finas, las cuales se tiñeron con azul de tripán según Phillips y Hayman (1970). Las raíces teñidas se cortaron en segmentos de ~2cm los cuales se montaron horizontalmente en un portaobjetos con glicerina. La presencia de micorrizas se cuantificó de acuerdo a McGonigle et al. (1990), lo que permite determinar el porcentaje de campos microscópicos en los que aparecen arbúsculos, vesículas e hifas micorrízicas.
A lo largo del experimento las plantas recibieron semanalmente una solución nutritiva completa según Hewitt (1952), a la cual se le redujo la dosis de fósforo a 1/5 de la original.
Ensayo para tratar de disminuir el peso del inoculante. Debido a que la mezcla arcilla/arena es muy pesada, se sustituyó la arena por un material mucho más liviano como lo es la cáscara de arroz, para así obtener un inoculante más fácil de transportar. Para ello se estableció un ensayo idéntico al anterior pero donde la arcilla se diluyó con cáscara de arroz en lugar de arena. Para ello se utilizaron las mismas diluciones que en el ensayo anterior (1/1, 1/2, 1/4 y 1/8 v/v), los mismos tratamientos e inóculos de HMA y la misma planta hospedera. Al final del ensayo se determinó el peso seco de las plantas así como la presencia de micorrizas siguiendo los mismos métodos señalados en el ensayo anterior.
Selección de la arcilla más apropiada para producir los inoculantes. Debido a que para producir los inoculantes es necesario contar con una fuente confiable y repetible de los materiales, se compararon dos fuentes de arcilla expansiva (B y C) diluidas con arena o cáscara de arroz. Para ello las arcillas de diluyeron 1/1 o 1/4 (v/v) con arena silícea ó con cáscara de arroz. Se hicieron dos tratamientos: 1) Control no micorrizado y 2) Inoculación con una mezcla de G. manihotis y S. fulgida. El inóculo y la planta hospedera fueron similares a los utilizados en el primero de estos ensayos.
El experimento se llevó a cabo por triplicado para un total de 24 plantas en todo el ensayo. El ensayo se mantuvo en el invernadero del IVIC por 1,5 meses al cabo de los cuales se evaluó el peso seco del vástago y el porcentaje de colonización micorrízica utilizando los mismos métodos señalados en el primer ensayo. Además, se cuantificó el número de esporas de HMA presentes en cada maceta al final del ensayo. Para ello se aislaron las esporas utilizando la mencionada técnica de Sieverding (1991). Una vez aisladas las esporas, se contaron bajo un microscopio estereoscópico y el número total se expresó por 100g de sustrato seco.
Validación del efecto de los inoculantes
La dilución y sustratos seleccionados en los experimentos anteriores (ver Resultados) se utilizaron para producir los inoculantes, por ser donde las micorrizas se reprodujeron de manera óptima. Las especies de HMA reproducidas en cada caso fueron las seleccionadas como mejores en el análisis de la colección de Glomeromycota del IVIC. Así se obtuvieron cinco tipos de inoculantes: Acaulospora lacunosa, Glomus manihotis, Entrophospora colombiana, Scutellospora heterogama y S. fulgida.
Validación en invernadero. Para evaluar la eficacia de tales inoculantes con una planta de interés agrícola se realizó un experimento en el invernadero del IVIC, con lechuga como planta hospedera. El experimento consistió en seis tratamientos: 1) Control, 2) Inoculado con G. manihotis, 3) Inoculado con S. fulgida, 4) Inoculado con S. heterogama, 5) Inoculado con E. colombiana, y 6) Inoculado con A. lacunosa
Al control se le adicionó una cantidad arcilla/arena equivalente a la aplicada en los tratamientos inoculados es decir, 30g. El suelo utilizado pertenece al bosque nublado localizado en terrenos del IVIC, con pH de 3,9; 0,41% N, contenido de materia orgánica de 5,37% y P intercambiable de 3,1µg·g-1 (Marulanda, 1998). Previamente a la instalación del ensayo, el suelo fue esterilizado mediante radiaciones gamma (>8 kGy). Con dicho suelo, el cual se mantuvo a capacidad de campo para el momento del transplante, se llenaron macetas plásticas de 1kg.
Semillas certificadas de Lacttuca sativa variedad americana (Grandes Lagos 659) fueron germinadas en cámaras húmedas y mantenidas en bandejas con suelo estéril por 45 días, cuando se transplantó una plántula a cada maceta. En los tratamientos inoculados se agregaron 30g del correspondiente inoculante en el hoyo de siembra. Se aplicaron 15ml de solución nutritiva (Hayman, 1974) por planta cada 7 días, durante los 80 días que duró el ensayo. Al final del experimento, se evaluó el peso seco de la parte aérea de las plantas y la presencia de micorrizas arbusculares en las raicillas utilizando la misma metodología señalada en el ensayo de optimización del sustrato.
Validación en el campo. Esta parte del trabajo se realizó al noroeste del Estado Amazonas, Venezuela, en un área utilizada típicamente para agricultura de subsistencia (tumba y quema del bosque) por los indígenas de la etnia Piaroa, en la comunidad de Coromoto de Cuao en la Reserva Forestal de Sipapo. En un área recientemente deforestada y quemada para esos fines se demarcó una parcela de 25×14m dentro de la cual se establecieron aleatoriamente cuatro parcelas de 3×3m, las cuales se asignaron también aleatoriamente a cada uno de los tratamientos. En cada parcela se sembraron 20 plantas de yuca dulce (Manihot utilissima) siguiendo la metodología típica empleada por los habitantes del área. Se realizaron cuatro tratamientos, a saber: 1) Control no inoculado (micorrizas nativas), 2) Inoculado con G. manihotis, 3) Inoculado con A. lacunosa, y 4) Inoculado con S. fulgida.
Se colocaron 100g de los inóculos en el hoyo de siembra. El experimento se mantuvo por 10 meses, tal como es lo usual para dicho cultivo en la zona. No se aplicaron fertilizantes ni agroquímicos, y el único manejo realizado fue la remoción de las malezas a mano. Al final del experimento se cosecharon 10 plantas de cada tratamiento y se determinó de cada una de ellas el peso fresco del tubérculo (yuca) producido.
Análisis estadístico
Los datos de los experimentos para preparación de los inoculantes y de validación en invernadero fueron analizados por ANOVA de dos o tres vías según el caso, seguido del test de Tukey para p<0,05. En todos los casos los valores de colonización micorrízica fueron transformados a arcoseno para cumplir con los supuestos del ANOVA. El experimento de validación en el campo se analizó por ANOVA seguido de una prueba de LSD (Zar, 1996).
Resultados
Los hongos de la colección de Glomeromycota del IVIC seleccionados para iniciar la producción de inoculantes aparecen en la Tabla I. Dichos inóculos poseían una pureza mayor al 95% y contenían números elevados de esporas que lucían intactas y con contenido lipídico visible, hecho que suele ser indicativo de un estado vital y saludable (Walker, 1992). El inóculo seleccionado de E. colombiana fue el que presentó la mayor concentración de esporas y el de A. lacunosa el menor.
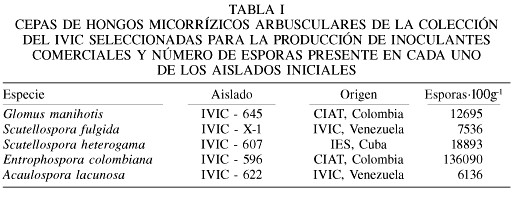
En el experimento llevado a cabo para optimizar la proporción arcilla/arena (Figura 1a) se obtuvo una clara respuesta del crecimiento de V. luteola ante la inoculación con HMA. En promedio, el peso seco de la planta aumentó en más de un 4000% en relación a las no inoculadas y la dilución 1/1 arcilla/arena fue la que favoreció más el crecimiento de las plantas micorrizadas.
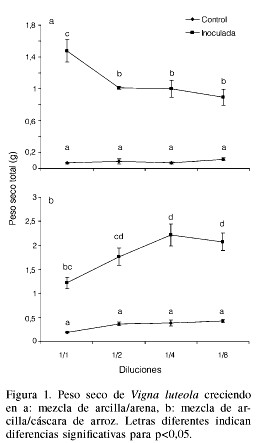
En el experimento realizado para tratar de sustituir la arena por un material más liviano (Figura 1b) las plantas en general produjeron más biomasa que cuando se utilizó la mezcla arcilla/arena. En promedio, las plantas de los tratamientos micorrizados presentaron un incremento respecto a los no micorrizados del 315%. En este experimento las diluciones que resultaron más favorables fueron la 1/4 y 1/8.
Al analizar la colonización micorrízica producida por los distintos tratamientos en este experimento (Tabla II) se observa un significativo efecto del sustrato sobre la formación de las micorrizas, siendo claro que la mezcla arcilla/arena fue la que arrojó los valores más elevados de todas las estructuras micorrízicas evaluadas. En particular se destacan las diluciones 1/1 y 1/4 en la mezcla arcilla/arena y la 1/1 en la mezcla arcilla/cáscara de arroz con los valores más elevados de colonización micorrízica.
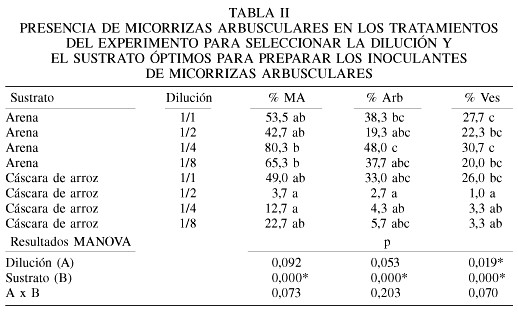
Sobre la base de estos resultados, para los experimentos subsiguientes se decidió utilizar las diluciones 1/1 y 1/4, pues parecen ser las más prometedoras tanto en la promoción del crecimiento de las plantas, como para la formación de micorrizas.
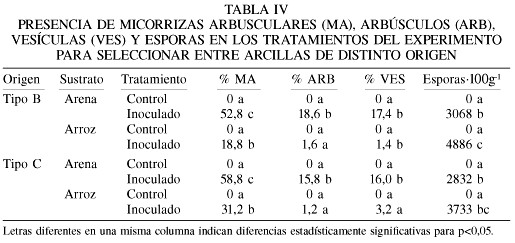
En cuanto a la selección de la fuente de arcilla más apropiada, los resultados del peso seco de V. luteola (Tabla III) indican una clara respuesta a las micorrizas en los experimentos cuando se utilizó la arcilla B (la misma utilizada en el experimento de optimización de la proporción arena/arcilla). Pero en cambio, cuando se utilizó la arcilla C mezclada con arena en la proporción 1/1 no se obtuvieron diferencias significativas entre el crecimiento de las plantas micorrizadas y los controles. Cuando dicha arcilla se mezcló con cáscara de arroz sí se produjeron diferencias estadísticamente significativas entre el tratamiento inoculado y el control, pero las plantas micorrizadas no incrementaron tanto su biomasa como cuando se utilizó la arcilla B en un tratamiento similar (2,52 vs 3,75g).

El análisis de la presencia de micorrizas en este experimento (Tabla IV) muestra de nuevo una mayor colonización micorrízica en los tratamientos donde se utilizó la arena para diluir la arcilla, en comparación a cuando se utilizó la cáscara de arroz. En contraste, la producción de esporas fue mayor en los tratamientos diluidos con cáscara de arroz pero especialmente cuando se utilizó la arcilla B. Así, los resultados obtenidos indican a la mezcla arcilla/arena 1/1 como la óptima para reproducir los HMA pre-seleccionados.
Cuando los inoculantes producidos fueron puestos a prueba en el invernadero, con lechuga Grandes lagos, se obtuvo un mayor crecimiento con los preparados de G. manihotis y S. fulgida (Figura 2). En dicho experimento todas las plantas control murieron antes del final del ensayo, salvo una, y el inoculante que resultó ser el menos efectivo fue el de S. heterogama. En este caso los resultados de la colonización micorrízica fueron inesperados (Tabla V) ya que el tratamiento inoculado con A. lacunosa presentó una mayor colonización en comparación con el tratamiento inoculado con E. colombiana, pero el crecimiento de la planta no tuvo ninguna relación con tales hechos. En general, los valores de colonización micorrízica obtenidos fueron de medios a altos y la única planta del control que sobrevivió se mantuvo libre de micorrizas hasta el final del experimento.
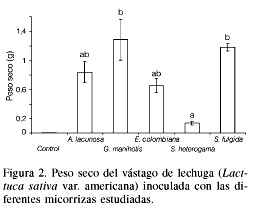

Finalmente, la aplicación de los preparados de HMA en suelos del Amazonas para la producción de yuca de la manera en que tradicionalmente lo hacen los habitantes de la zona arrojó resultados muy prometedores (Tabla VI). En efecto, la aplicación del inoculante con G. manihotis produjo un incremento en la cosecha de más del 200%, sin adición de fertilizantes químicos. Por otra parte, aunque la inoculación con A. lacunosa produjo un incremento en la producción de tubérculos, esta diferencia no fue significativa respecto al control.

Discusión
Los hongos de la colección de Glomeromycota del IVIC que fueron preseleccionados para producir los inoculantes comerciales tenían concentraciones muy altas de esporas, las cuales son consideradas "saturantes" es decir, debido a que su concentración es muy superior a la que se encuentra normalmente en un suelo agrícola (Sieverding, 1991), el número de esporas no debió ser un factor limitante en la producción de los inoculantes.
Los experimentos realizados con los diversos materiales y diluciones ilustran que al producir un inoculante de HMA se debe ensayar con las características del sustrato, como aireación y fuentes de materiales confiables, de manera de producir un preparado que se adapte a las condiciones de crecimiento de los hongos y a las condiciones ecológicas a las que se enfrentará el producto en el campo (Abbott et al., 1992)
Los resultados presentados indican la alta dependencia micorrízica presentada por la planta utilizada para producir los inoculantes, es decir V. luteola (Hernández et al., 2000), característica ésta que fue decisiva al seleccionarla como hospedera. En efecto, las plantas control prácticamente no crecieron ni en el experimento para optimizar el sustrato ni en el ensayo para disminuir el peso del inoculante.
Los resultados acerca de la presencia de micorrizas utilizando arena o cáscara de arroz para diluir la arcilla arrojaron valores mayores de las variables micorrízicas analizadas cuando se utilizó arena que cuando se utilizó cáscara de arroz. Un resultado similar ha sido reportado por Gaur y Adholeya (2000) quienes obtuvieron una óptima producción de inóculos de MA cuando utilizaron materiales con partículas con diámetro de 0,50-0,78mm. Dichos autores atribuyen el efecto a una mayor aireación, drenaje, suministro de O2 y crecimiento radical, y señalan que cuando utilizaron partículas de mayor tamaño no se formaron tantos propágalos. Lo atribuyeron a que las partículas más grandes poseen una menor capacidad de retención de agua, lo cual puede causar condiciones de baja disponibilidad de agua, más frecuentemente durante la producción de inoculantes. En el presente caso, aunque se utilizó arena bastante gruesa (2,06mm), se obtuvo con ella una mayor micorrización que cuando se utilizó la cáscara de arroz, de mucho mayor tamaño (en promedio 9,80mm), probablemente debido a las razones señaladas por Gaur y Adholeya (2000).
La total ausencia de micorrizas en los controles de los tres ensayos comentados señala que los materiales seleccionados están libres de esporas y otros propágulos de MA. En este mismo experimento se obtuvo una mayor esporulación por parte de las plantas que crecieron en la mezcla arcilla/cáscara de arroz, en comparación con las que crecieron en la mezcla arcilla/arena. También, en este caso los tratamientos con la arcilla B fueron los que produjeron la mayor cantidad de esporas. En efecto, dicha cantidad de esporas es muy elevada si se toma en cuenta que este experimento solo duró un mes y medio.
En la literatura aparece reportada la tendencia de los HMA a esporular dentro de otras esporas de HMA (Koske, 1984) sugiriendo ello que las esporas muertas de los HMA proveen de un microhábitat favorable para la formación de esporas. También se ha reportado una mayor esporulación de HMA dentro de restos de insectos, semillas muertas y cubiertas seminales (Rabatin y Rhodes, 1982), lo que ha sido interpretado como un mecanismo que favorecería la supervivencia de las esporas expuestas a condiciones ambientales adversas. Este hecho ha sido aprovechado por algunos autores para la utilización de arcillas expandidas en la producción de inoculantes comerciales (Feldmann e Idczak, 1992), dado que las esporas proliferan dentro de las cavidades producidas por las arcillas. En este caso, la cáscara de arroz seguramente provee un microhábitat más favorable para la esporulación que las partículas de arcilla no expandidas.
La validación de los experimentos realizados con la lechuga en el invernadero mostró varios aspectos a ser tomados en cuenta por los productores de inoculantes de MA: En primer lugar, aunque la simbiosis micorrízica arbuscular carece de especificidad taxonómica (Smith y Read, 1997) obviamente no todas las especies de HMA son equivalentes. Es decir, existe una cierta "compatibilidad funcional" entre la planta, el suelo y el hongo, y obviamente hay combinaciones que funcionan mejor que otras (van der Heijden et al., 1998). De allí que la producción y comercialización de los inoculantes de HMA requiera la aplicación de un protocolo previo a la parcela del productor o cliente potencial, que permita indagar rápidamente si las plantas que se pondrán a crecer en una parcela particular necesitan ser inoculadas o no. Ello va a depender del número de esporas presentes en la comunidad de HMA nativas, del potencial micorrízico del suelo y de sus niveles nutricionales.
También es indispensable estudiar previamente el comportamiento de los diversos inoculantes, de manera de poder seleccionar una especie de HMA adaptada a las condiciones edáficas en las que se va a aplicar y estudiar, con anterioridad a su empleo, el comportamiento de los diversos inoculantes que se van a comercializar con los cultivos de interés. En el caso presente resultó claro que el inoculante de S. heterogama es una especie de HMA inadecuada para la lechuga sembrada en un suelo ácido. Ello resulta lógico si tomamos en cuenta el origen de S. heterogama, la cual proviene de Cuba (Tabla I) y por lo tanto de suelos mucho más fértiles que los utilizados en el ensayo de invernadero presentado en este trabajo.
Por otra parte, G. manihotis y S. fulgida, ambas bien adaptadas a suelos ácidos, resultaron ser las que promovieron de manera más efectiva el crecimiento de la lechuga. Inesperado fue, sin embargo, el resultado obtenido con E. colombiana, el cual es un hongo muy conocido por su eficiencia en suelos ácidos (Sieverding, 1991) y que en este caso, sin embargo, produjo un resultado más bien pobre al igual que A. lacunosa, esta última a pesar del alto nivel de colonización micorrízica producido.
Los resultados obtenidos apoyan hechos señalados previamente por otros autores, que indican que los porcentajes de colonización micorrízica no son buenos índices para predecir la respuesta a la inoculación con HMA y que tanto la medición de la colonización interna como la externa (micelio externo) son necesarias para tener una representación certera de la comunidad micorrízica presente (Hart y Reader, 2002). Lamentablemente la determinación del micelio externo es una medida que requiere una gran inversión de tiempo y minuciosidad, y sería poco práctico proponer su manejo como parte de la producción de inoculantes.
El inoculante de A. lacunosa resultó ineficaz en la promoción del crecimiento tanto de la lechuga como de la yuca, a pesar de su indudable capacidad infectiva. Podría presumirse que especies como ésta cumplen en la naturaleza funciones no nutritivas, pues de otra forma deberían haber sido eliminadas por efecto de la selección natural (Newsham et al., 1995). Por lo tanto hay que estar atentos, a la hora de involucrarse en la producción de inoculantes de HMA, acerca de cuáles cepas recomendar y para qué circunstancias, lo que hace indispensable imprimirle un alto contenido ético a tal actividad y no tratarla simplemente como un negocio lucrativo, debido a las consecuencias ecológicas que podría traer el uso de hongos que no funcionen como mutualistas en determinadas circunstancias (Johnson, 1993).
Finalmente, el efecto logrado en un sistema como el de Amazonas es alentador. Uno de los aspectos más importantes a considerar es la ubicación geográfica, las características ecológicas y sociales de la zona. El difícil acceso a la zona de estudio y las pocas fuentes de trabajo en las comunidades Piaroa no permiten el desarrollo de una economía basada en la producción de excedentes de yuca para el comercio. En el caso del experimento realizado en el Amazonas podía haberse anticipado una gran respuesta ante la inoculación debido a la bien conocida escasez nutricional de dichos suelos (Herrera et al., 1978). Además, resultados previos obtenidos en esa misma zona (Kalinhoff y Cáceres, 2002) han demostrado que el número de propágulos infectivos (número de esporas) es bajo en comparación con el número de propágulos de los inoculantes preparados artificialmente. Por esta razón, el manejo de las micorrizas por las comunidades indígenas permitiría implementar tecnologías novedosas y de bajo costo, y al mismo tiempo contribuir a la conservación del ambiente. Particularmente, en esas comunidades Piaroa el uso de agroquímicos como insumo para incrementar la productividad de los cultivos no es viable, no solamente por el alto costo, sino porque ellos llevan a cabo una agricultura de subsistencia orientada a satisfacer las necesidades básicas de alimento, y solo ocasionalmente llega a constituir una fuente de ingresos a través de la venta de excedentes de algunos productos. Para alcanzar estas metas es necesario mantener e implementar proyectos que permitan la continuidad en el tiempo de estas líneas de acción en la región.
En suma, en suelos pobres en fósforo disponible como los que abundan en Venezuela y en muchas regiones tropicales, las micorrizas surgen como una herramienta ecológica y productiva, cuya implementación debería ser encarada pronto y con un alto contenido ético por los profesionales relacionados con la agricultura.
Agradecimientos
Los autores agradecen el apoyo de Erasmo Meneses, Milagros Lovera, Carolina Kalinhoff y Víctor Romero tanto en el trabajo de campo como de laboratorio. Este trabajo fue financiado parcialmente con fondos del Proyecto 2004000368 del BID-Fonacit II.
REFERENCIAS
1. Abbott LK, Robson AD, Gazey C (1992) Selection of inoculant vesicular-arbuscular mycorrhizal fungi. En Norris JR, Read DJ, Varma AK (Eds.) Methods in Microbiology, Vol 24. Academic Press. San Diego, CA, EEUU. pp 1-21. [ Links ]
2. Azcón-Aguilar C, Barea JM (1992) Interactions between mycorrhizal fungi and other rhizosphere microorganisms. En Allen MF (Ed.) Mycorrhizal Functioning. An Integrative Plant Fungal Process. Chapman & Hall. New York, EEUU. pp 163-198 [ Links ]
3. Azcón-Aguilar C, Barea JM (1999) Saprophytic growth of arbuscular mycorrhizal fungi. En Varma A, Hock B (Eds.) Mycorrhiza. 2nd ed. Springer. Heidelberg, Alemania. pp 391-408. [ Links ]
4. Brundrett M, Bougler N, Dell B, Grove T, Malajczuk N (1996) Working with Mycorrhizas in Forestry and Agriculture: Australian Centre for International Agricultural Research. Canberra, Australia. 344 pp. [ Links ]
5. Dodd JC, Burton CC, Burns RG, Jeffries P (1987) Phosphatase activity associated with the roots and the rhizosphere of plants infected with vesicular-arbuscular mycorrhizal fungi. New Phytol. 107: 163-172. [ Links ]
6. Feldmann F, Idczak E (1992) Inoculum production of vesicular-arbuscular mycorrhizal fungi for use in tropical nurseries. En Norris JR, Read DJ, Varma AK (Eds.) Methods in Microbiology. Vol 24. Academic Press. San Diego, CA, EEUU. pp 339-357. [ Links ]
7. Gaur A, Adholeya A (2000) Effects of the particle size of soil-less substrates upon AM fungus inoculum production. Mycorrhiza 10: 43-48. [ Links ]
8. González EJ (2004) Lemna en el Lago de Maracaibo (Venezuela). Red de Eutrofización de Lagos y Embalses. http://rele.fcien.edu.uy/publicaciones/ [ Links ]
9. Hart M, Reader RJ (2002) Taxonomic basis for variation in the colonization strategy of arbuscular mycorrhizal fungi. New Phytol. 153: 335-344. [ Links ]
10. Hayman DS (1974) Plant growth responses to vesicular-arbuscular mycorrhiza. VI: Effect of light and temperature. New Phytol. 73: 71-80. [ Links ]
11. Herrera R, Jordan CF, Klinge H, Medina E (1978) Amazon ecosystems: their structure and functioning with particular emphasis on nutrients. Interciencia 3: 223-232. [ Links ]
12. Herrera-Peraza RA, Ferrer RL, Orozco MO, Hernández G, Vancura V (1984) Fertilización y micorrizas VA. II. Análisis del balance de macroelementos en varios experimentos. Acta Bot. Cubana 20: 111-142. [ Links ]
13. Hernández G, Cuenca G, García A (2000) Behaviour of arbuscular-mycorrhizal fungi on Vigna luteola growth and its effect on the exchangeable (32P) phosphorus of soil. Biol. Fert. Soils 31: 232-236. [ Links ]
14. Hewitt EJ (1952) Sand and water culture methods used in the study of plant nutrition: Tech Comm 22 Farnhan Royal Commonwealth Agricultural Bureau. Buks, RU pp. 193. [ Links ]
15. Infante A, Infante O, Márquez M, Lewis WM, Weibezahn F (1979) Conditions leading to mass mortality of fish and zooplankton in Lake Valencia, Venezuela. Acta Cient. Venez. 30: 67-73. [ Links ]
16. Jeffries P, Barea JM (2001) Arbuscular Mycorhiza- a key component of sustainable plant-soil ecosystems. En Hock B (Ed.) The Mycota IX Fungal Associations. Springer. Berlín, Alemania. pp. 95-113. [ Links ]
17. Johnson NC (1993) Can fertilization of soil select less mutualistic mycorrhizae? Ecol. Applic. 3: 749-757. [ Links ]
18. Joner EJ, Johansen A (2000) Phosphatase activity of external hyphae of two arbuscular mycorrhizal fungi. Mycol. Res. 104: 81-86. [ Links ]
19. Kalinhoff C, Cáceres A (2002) Efecto de la perturbación producida por el establecimiento de conucos tradicionales Piaroas sobre las micorrizas arbusculares (MA) y la biomasa de raíces finas en la Reserva Forestal Sipapo, Edo Amazonas. Tesis. Universidad Central de Venezuela. pp.135. [ Links ]
20. Koske RE (1984) Spores of VAM fungi inside spores of VAM fungi. Mycologia 76: 853-862. [ Links ]
21. Marulanda O (1998) Efectos de los cambios florísticos y estructurales en la dinámica de raíces finas y el flujo de CO2 del suelo en un mosaico sucesional de la Cordillera de la Costa, Altos de Pipe. Tesis, Instituto Venezolano de Investigaciones Científicas. pp. 222. [ Links ]
22. McGonigle TP, Miller JE, Evans DG, Fairchild GL, Swan JA (1990) A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 115: 495-501. [ Links ]
23. Miller RM, Jastrow JD (2000) Mycorrhizal fungi influence soil structure. En Kapulnik Y, Douds DD (Eds.) Arbuscular Mycorrhizas: Physiology and Function. Kluwer. Dordrecht, Holanda. pp. 3-18. [ Links ]
24. Newsham KK, Fitter AH, Watkinson AR (1995) Arbuscular mycorrhiza protects an annual grass from root pathogenic fungi in the field. J. Ecol. 83: 991-1000. [ Links ]
25. Paolini J (1988) Physico-chemical properties of Venezuelan soils and their sensitivity to acidification. En Rodhe H, Herrera R (Eds.) Acidification in Tropical Countries. Wiley. Chichester, RU. pp 201-210. [ Links ]
26. Phillips JM, Hayman DS (1970) Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Brit. Mycol. Soc. 55: 158-161. [ Links ]
27. Rabatin SC, Rhodes LH (1982) Acaulospora bireticulata inside oribatid mites. Mycologia 74: 859-861. [ Links ]
28. Rillig M (2004) Arbuscular mycorrhizae, glomalin, and soil aggregation. Can. J. Soil Sci. 84: 355-363. [ Links ]
29. Ruiz-Lozano JM, Azcón R (1995) Hyphal cotribution to water uptake in mycorrhizal plants as affected by the fungal species and water status. Physiol. Plant. 95: 472-478. [ Links ]
30. Sieverding E (1991) Vesicular-arbuscular mycorrhiza management in tropical agrosystems. GTZ. Eschborn, Alemania. pp. 371. [ Links ]
31. Smith, SL, Read, D (1997) Mycorrhizal symbiosis. 2ª edición. Capítulo 1. Academic Press. [ Links ]
32. Trappe JM (1987) Phylogenetic and ecological aspects of mycotrophy in the angiosperms from an evolutionary standpoint. En Safir DG (Ed.) Ecophysiology of VA mycorrhizal plants. CRC. Boca Raton, FL, EEUU. pp 5-25. [ Links ]
33. van der Heijden MGA, Klironomos JN, Ursic M, Moutoglis P, Streitwolf-Engel R, Boller T, Wiemken A, Sanders IR (1998) Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396: 69-72. [ Links ]
34. Walker C (1992) Systematics and taxonomy of the arbuscular endomycorrhizal fungi (Glomales) a possible way forward. Agronomie 12: 887-897. [ Links ]
35. Zar JH (1996) Biostatistical Analysis. 3ª ed. Prentice Hall. Upper Saddle River, NJ, EEUU. pp. 662. [ Links ]
