











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.32 n.12 Caracas dic. 2007
EFECTO DE ENZIMAS FIBROLÍTICAS EXÓGENAS EN LA DEGRADACIÓN RUMINAL IN VITRO DE DIETAS PARA VACAS LECHERAS
Robert Moreno, Juan Manuel Pinos-Rodríguez, Sergio González, Gregorio Álvarez, Juan Carlos García, Germán Mendoza y Ricardo Bárcena
Robert Ángel Moreno Jaramillo. MVZ, Doctor en Ciencias en Nutrición de Rumiantes, Colegio de Postgraduados (COLPOS), Montecillo, México. Profesor, Universidad Autónoma Gabriel René Moreno, Santa Cruz, Bolivia.
Juan Manuel Pinos-Rodríguez. Médico Veterinario, Universidad Veracruzana, México. Maestro y Doctor en Ciencias, COLPOS, Montecillo, México. Profesor Investigador, Universidad Autónoma de San Luis Potosí (UASLP), México. Dirección: Instituto de Investigación de Zonas Desérticas, UASLP. Altair 200, Fracc. del Llano. 78377. San Luis Potosí, S.L.P. México. e-mail: jpinos@uaslp.mx
Sergio S. González Muñoz. Ph.D., University of Nebraska, EEUU. Profesor Investigador, COLPOS, Montecillo, México.
Gregorio Álvarez. Doctor en Ciencias en Nutrición de Rumiantes, COLPOS, Montecillo, México. Profesor Investigador, UASLP, México.
Juan Carlos García López. Doctor en Ciencias, COLPOS, Montecillo, México. Profesor Investigador, UASLP, México.
Germán David Mendoza Martínez. Ph.D., University of Nebraska, EEUU. Profesor Investigador, Universidad Autónoma Metropolitana, Xochimilco, México.
Ricardo Bárcena. Ph.D. University of Arizona, EEUU.
RESUMEN
Para determinar el efecto in vitro de enzimas fibrolíticas exógenas en la degradación ruminal de la materia seca (DIVMS) y de la fibra detergente neutro (DIVFDN) se incubaron de 3 a 72h en licor ruminal, tres dietas con 40:60, 50:50 y 60:40 relación forraje:concentrado (F:C), con o sin enzimas fibrolíticas. Se usó un diseño de bloques completos al azar con un arreglo factorial 3×2×6 (ración F:C de 40:60, 50:50, 60:40; enzima, 0 o 2g·kg-1 MS; tiempo de incubación de 3, 6, 12, 24, 48 y 72h). La DIVMS de dieta 60:40 F:C fue mayor (P≤0,05) que la de 50:50 o 40:60. Por su parte, la DIVFDN de la dieta 50:50 F:C fue mayor que la de 40:60 o 60:40. La interacción entre F:C de la dieta, enzima y el tiempo de incubación indicó que las enzimas fibrolíticas exógenas tuvieron mayor impacto en la DIVMS y en la DIVFDN en las primeras 12h de incubación en la dieta con mayor contenido de fibra.
EFFECT OF EXOGENOUS FIBROLYTYC ENZYME ON IN VITRO RUMINAL DEGRADATION OF RATIONS FOR DAIRY COWS
SUMMARY
To determine the effect of an exogenous fibrolytic enzyme on in vitro dry matter and neutral detergent fiber degradation (IVDDM and IVDNDF, respectively), three total mixed rations with 40:60, 50:50, 60:40 forage:concentrate ratio (F:C), each with or without enzyme, were incubated for 3 to 72h. A randomized block design with a factorial arrangement of treatments 3×2×6 was utilized (FC ratios of 40:60, 50:50, 60:40; enzyme, 0 and 2g·kg-1 DM; incubation times of 3, 6, 12, 24, 48 and 72h). The IVDDM was higher (P≤0,05) in rations with F:C of 60:40 than 50:50 or 40:60 F:C. The IVDNDF was higher with 50:50 than 40:60 or 60:40. The F:C × enzyme × incubation time suggests that the main effect of the fibrolytic enzymes on IVDDM and IVDNDF were found during the first 12h of incubation in the diet with the highest fiber content.
EFEITO DE ENZIMAS FIBROLÍTICAS EXÓGENAS NA DEGRADAÇÃO RUMINAL IN VITRO DE DIETAS PARA VACAS LEITEIRAS
RESUMO
Para determinar o efeito in vitro de enzimas fibrolíticas exógenas na degradação ruminal da matéria seca (DIVMS) e da fibra detergente neutro (DIVFDN) se incubaram de 3 a 72h em licor ruminal, três dietas com 40:60, 50:50 e 60:40 relação forragem:concentrado (F:C), com ou sem enzimas fibrolíticas. Foi utilizado um desenho de blocos completos aleatório com um arranjo fatorial 3×2×6 (ração F:C de 40:60, 50:50, 60:40; enzima, 0 o 2g·kg-1 MS; tempo de incubação de 3, 6, 12, 24, 48 e 72h). A DIVMS de dieta 60:40 F:C foi maior (P≤0,05) que a de 50:50 ou 40:60. Por sua parte, a DIVFDN da dieta 50:50 F:C foi maior que a de 40:60 ou 60:40. A interação entre F:C da dieta, enzima e o tempo de incubação indicou que as enzimas fibrolíticas exógenas tiveram maior impacto na DIVMS e na DIVFDN nas primeiras 12h de incubação na dieta com maior conteúdo de fibra.
PALABRAS CLAVE / Degradación In Vitro / Enzimas Fibrolíticas / Vacas Lecheras /
Recibido: 26/12/2006. Modificado: 08/10/2007. Aceptado: 10/10/2007.
Introducción
La alimentación dentro de un sistema de producción pecuaria es el mayor rubro económico, por lo que se han empleado diversas tecnologías para mejorar la utilización de los nutrientes, principalmente de los carbohidratos estructurales de la fibra. La adición de enzimas fibrolíticas en alimentos para rumiantes puede estimular la ganancia diaria de peso (Beauchemin et al., 1995) y la producción láctea (Lewis et al., 1999; Rode et al., 1999; Yang et al., 2000; Pinos-Rodríguez et al., 2005). Ello se ha explicado por el efecto positivo que ejercen las enzimas exógenas sobre la hidrólisis de los carbohidratos de la pared celular, favoreciendo un aumento en la cantidad de sustrato disponible para los microorganismos ruminales (Pinos-Rodríguez et al., 2002a; Jurkovich et al., 2006). Sin embargo, en varios estudios in vitro (Feng et al., 1996; Sheperd y Kung, 1996; Kung et al., 2000; Pinos-Rodríguez et al., 2001) los resultados observados sobre la digestión de la fibra detergente neutro (FDN) y de la materia seca (MS) no han sido consistentes. Aún cuando varios factores pueden afectar el nivel de respuesta a la adición de enzimas exógenas (Beauchemin et al., 2003), aquellas que comprometen la actividad fibrolítica ruminal parecen ser factores relevantes. Ejemplos de ellos son la presentación física del forraje (López et al., 2006) y posiblemente el contenido de carbohidratos de fácil fermentación en la ración.
El objetivo de la presente investigación fue evaluar la respuesta de la utilización de una enzima exógena en la degradación ruminal in vitro de la materia seca (DIVMS) y de la fibra detergente neutro (DIVFDN) de dietas con tres (40:60, 50:50 y 60:40) proporciones de forraje:concentrado (F:C).
Materiales y Métodos
El experimento se realizó en el Instituto de Investigaciones de Zonas Desérticas, de la Universidad Autónoma de San Luis Potosí, México. Se utilizó como donador de inóculo un novillo B. taurus×B. indicus (450kg PV), con cánula ruminal y alimentado con una ración con 50:50 forraje:concentrado (F:C). El agua, alimento y las sales minerales (en mg·kg-1: 10 Se, 215 K, 50 Fe, 20 Co, 50 Zn, 1600 Mn, 300 Cu, 70 I, 220 Ca, 280 P; S 30 g·kg-1; y sal común 950 g·kg-1) se ofrecieron ad libitum. Después de tres semanas de adaptación se recolectó líquido ruminal con una bomba de vacío; el fluido se filtró y conservó en un termo a 40°C.
Las dietas contenían los mismos ingredientes (Tabla I) pero con diferente proporción F:C (40:60, 50:50 o 60:40), y fueron incubadas in vitro con y sin enzima. El compuesto enzimático fibrolítico exógeno (Fibrozyme, Alltech Inc., Nicholasville, KY, EEUU.) es un extracto de la fermentación de Aspergillus niger y Trichoderma viride. Su actividad xilanásica (una unidad xilanásica es la cantidad de enzima necesaria para liberar un µmol de xilosa) reportada por el fabricante es 100UI·g-1. La dosis empleada del compuesto enzimático (recomendada por el fabricante) fue de 2g·kg-1 MS de dieta.
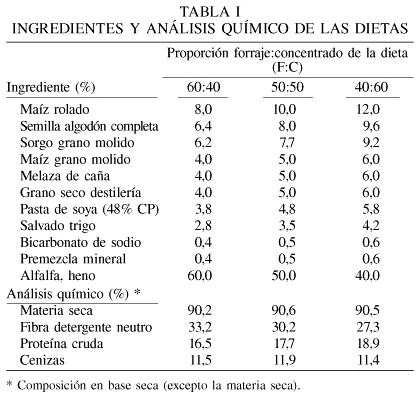
Las muestras de forraje fueron molidas (molino Willey) con malla de 1mm para el análisis químico, y con malla de 2mm para la degradación ruminal in vitro. En cada dieta se determinó el contenido de MS en una estufa de aire forzado con una temperatura de 65°C durante 48h, así como de MO, proteína (%N×6,25), cenizas (AOAC, 1990) y FDN (Van Soest et al., 1991).
Para la degradación ruminal in vitro de la materia seca (DIVMS), 500mg de cada dieta experimental (previo secado a 65°C por 24h) fueron incubadas en tubos de propileno con 50ml de una mezcla (4:1) de saliva de McDougall y licor ruminal durante 3, 6, 12, 24, 48 y 72h (tiempo de incubación; TI) con o sin el agregado de enzima fibrolítica. Para la fibra detergente neutro (DIVFDN) se siguió el mismo protocolo, pero el sustrato fue un residuo insoluble en solución detergente neutro para cada dieta. Para obtener la FDN, 500g de cada alimento molido (malla 2mm) fueron colocados en bolsas de poliseda (poro de 52 ±10µm) y depositados en un recipiente con 12 litros de una solución detergente neutro (Van Soest et al., 1991), el cual permaneció en ebullición durante 1h a 90°C. Posteriormente las bolsas fueron lavadas hasta remover completamente el detergente, se secaron al ambiente (12h) y se colocaron en estufa a 55°C hasta obtener un peso seco constante (24h en promedio). Cada uno de los residuos (correspondiente a las tres dietas) fue fraccionado en dos, utilizando uno como testigo y al otro se le agregó el compuesto enzimático. La DIVMS y la DIVFDN fueron realizadas por duplicado con un intervalo de tiempo de dos semanas. La tasa de degradación (kd) y el tiempo medio (T½) fueron calculados con un análisis de regresión de acuerdo a Mertens y Ely (1982).
El análisis estadístico de las variables respuesta (DIVMS y DIVFDN) se efectuó bajo un diseño en bloques completos al azar, con un arreglo factorial 3×2×6, donde los factores principales fueron relación forraje:concentrado (60:40, 50:50, y 40:60), nivel de enzima (0 o 2g·kg-1 MS), y factor tiempo de incubación (3, 6, 12, 24, 48 y 72h). El criterio utilizado para la conformación de los bloques fueron las corridas (n= 2), las cuales se realizaron con un mes de diferencia entre ambas. Los datos fueron analizados con el procedimiento GLM del SAS (1999). La comparación de medias entre niveles de cada factor se realizó con la prueba de Tukey, y por contrastes ortogonales (Steel y Torrie, 1986) para aquellos tratamientos con tiempos equivalentes dentro de cada ración totalmente mezclada (RTM) con y sin compuesto enzimático.
Resultados y Discusión
La Tabla II presenta la significancia y el error estándar de la media de los factores individuales y de sus interacciones. Para DIVMS la interacción F:C × tiempo de incubación sugiere que durante las primeras 6h, la MS de la dieta con F:C de 40:60 se degrada más rápidamente que aquellas con 50:50 o 60:40, posiblemente como resultado de la mayor concentración de carbohidratos no estructurales, fácilmente degradables en rumen (Rooney y Pflugfelder, 1986). Después de 12h de incubación la DIVMS fue similar en todas las dietas. El nivel de forraje en las dietas tuvo una respuesta semejante a un efecto cuadrático en la DIVFDN. En dietas con 40 y 60% de concentrado la DIVFDN no fue diferente; sin embargo, la dieta con 50% de concentrado tuvo una DIVFDN mayor que con las otras dos. Hay evidencias (Grant, 1994) que muestran que el pH bajo, generado por dietas con niveles altos en concentrado, disminuye DIVFDN, pero los presentes resultados muestran con 50% de forraje una mayor DIVFDN que con 60%.
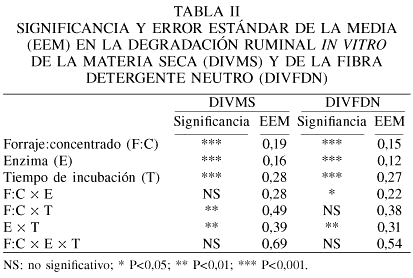
La DIVMS fue de 50,3% sin enzima y 51,6% con enzima. No se encontró interacción significativa entre F:C de la ración y el nivel de enzima. Ello sugiere que la enzima a 2g·kg-1 MS mejoró la DIVMS un 2,5% en comparación con el nivel 0 de enzima, independientemente de F:C en la ración. Resultados similares fueron encontrados por Feng et al. (1996).
Los valores de DIVMS y de DIVFDN de la interacción enzima × tiempo de incubación son presentados en la Figura 1. La enzima mejoró la DIVMS y la DIVFDN durante las primeras horas de incubación, quedando de manifiesto que la viabilidad de la enzima dependerá del tiempo de exposición al ambiente ruminal. Después de 12h, ningún efecto de la enzima fue observado. Estas observaciones concuerdan con otros estudios (Nsereko et al., 2000; Colombatto et al., 2003; Giraldo et al., 2007) que muestran que las enzimas estimulan la fase inicial de degradación del sustrato en cuestión, pero esos efectos disminuyen conforme el tiempo de incubación aumenta. Esos resultados apoyan la idea general de que las enzimas incrementan la tasa, pero no la extensión, de la degradación ruminal de los alimentos (Pinos-Rodríguez et al., 2002b; Giraldo et al., 2007).
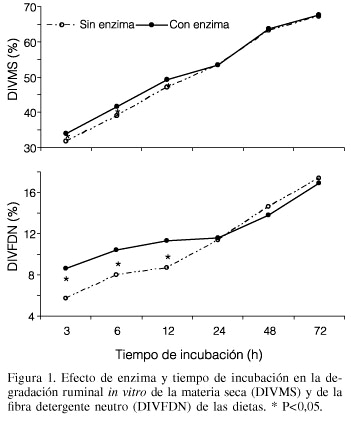
La interacción F:C × enzima en la DIVFDN indica que el efecto de la enzima depende del F:C de la dieta (Figura 2). En las dietas 60:40 y 50:50, las enzimas incrementaron la DIVMS, pero no así en la dieta 40:60. Este resultado posiblemente se debe a que en las dietas con mayor cantidad de fibra, las enzimas xilanolíticas presentes en el compuesto enzimático cuentan con mayor cantidad de sustrato (FDN) para actuar, aunado al posible pH más cercano a la neutralidad, que beneficia su mecanismo de acción (Colombatto et al., 2007). En un experimento previo (Pinos-Rodríguez et al., 2005) se encontró que estas enzimas fibrolíticas exógenas mejoraron, en mayor medida, la DIVMS de ingredientes fibrosos (cascarilla de soya y ensilado de maíz) que de ingredientes energéticos o proteínicos (maíz y pasta de soya).
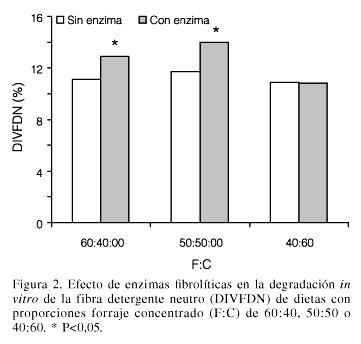
Hay evidencias que sugieren que las enzimas fibrolíticas exógenas además de actuar como agentes degradantes de la fibra, promueven una mayor colonización del alimento e incrementan el número de microorganismos, tanto fibrolíticos como no fibrolíticos en el líquido ruminal; lo cual incrementa la degradación en la fracción del alimento digerida más lentamente por los microorganismos ruminales (Bowman et al., 2003).
La cinética ruminal (kd y T½) de DIVMS y DIVFDN no fue afectada por F:C ni el nivel de enzima presente. La falta de efecto de enzimas fibrolíticas sobre la degradación de alimentos para rumiantes ha sido reportado en varios estudios (Sheperd y Kung, 1996). Sin embargo, otros (Pinos-Rodríguez et al., 2002b: Colombatto et al., 2007; Eun y Beauchemin, 2007; Giraldo et al., 2007) demuestran las ventajas del uso de las enzimas fibrolíticas exógenas para mejorar la digestibilidad de alimentos para rumiantes bajo diferentes condiciones experimentales.
Conclusiones
Bajo las condiciones descritas en el presente experimento, los resultados obtenidos permiten concluir que el efecto positivo de las enzimas evaluadas sobre la degradación de la fibra permanece por 12h, pero que este efecto es mayor cuando al menos un 50% de forraje está presente en las dietas para vacas lecheras.
REFERENCIAS
1. AOAC (1990) Official Methods of Analysis. 15th ed. Association of Official Analytical Chemists. Washington, D.C. 1928 pp. [ Links ]
2. Beauchemin KA, Rode LM, Sewalt VJ (1995) Fibrolytic enzymes increase fiber digestibility and growth rate of steers fed dry forages. Can. J. Anim. Sci. 75: 641-644. [ Links ]
3. Beauchemin KA, Colombatto D, Morgavi DP, Yang WZ (2003) Use of exogenous fibrolytic enzymes to improve feed utilization by ruminants. J. Anim. Sci. 81(Suppl. 2): E37-E47. [ Links ]
4. Bowman GR, Beauchemin KA, Shelford JA (2003) Fibrolityc enzymes and parity effects on feeding behavior, salivation, and ruminal pH of lactating dairy cows. J. Dairy Sci. 86: 565-575. [ Links ]
5. Colombatto D, Hervás G, Yang WZ, Beauchemin KA (2003) Effects of enzyme supplementation of a total mixed ration on microbial fermentation in continuous culture, maintained at high and low pH. J. Anim. Sci. 81: 2617-2627. [ Links ]
6. Colombatto D, Mould FL, Bhat MK, Owen E (2007) Influence of exogenous fibrolytic enzyme level and incubation pH on the in vitro ruminal fermentation of alfalfa stems. Anim. Feed Sci. Technol. 137: 150-162. [ Links ]
7. Eun JS, Beauchemin KA (2007) Assessment of the efficacy of varying experimental exogenous fibrolytic enzymes using in vitro fermentation characteristics. Anim. Feed Sci. Technol. 132: 298-315. [ Links ]
8. Feng P, Hunt CW, Pritchard GT, Julien WE (1996) Effect of enzyme preparations on in situ and in vitro degradation and in vivo digestive characteristics of nature cool season grass forage in beef steers. J. Anim. Sci. 74: 1349-1357. [ Links ]
9. Giraldo LA, Tejido ML, Ranilla MJ, Carro MD (2007) Effects of exogenous fibrolytic enzymes on in vitro ruminal fermentation of substrates with different forage:concentrate ratios. Anim. Feed Sci. Technol. doi: 10.1016/j.anifeedsci.2007.06.013 [ Links ]
10. Grant RJ (1994) Influence of corn and sorghum starch on the in vitro kinetics of forage fiber digestion. J. Dairy Sci. 77: 1563-1569. [ Links ]
11. Jurkovich V, Kutasi J, Febel H, Reiczigel J, Brydl E, Konyves L, Rafai P (2006) Rumen fermentation response to a direct-fed xylanase enzyme preparation from Thermomyces lanuginosus in sheep. Acta Vet. Hung. 54: 333-342. [ Links ]
12. Kung LJr, Treacher RJ, Nauman GA, Smagala AM, Endres KM, Cohen MA (2000) The effect of treating forages with fibrolytic enzymes on its nutritive value and lactation performance of dairy cows. J. Dairy Sci. 83: 115-122. [ Links ]
13. Lewis GE, Sánchez WK, Hunt CW, Guy MA, Pritchard GT, Swanson BI, Treacher RJ (1999) Effect of direct-fed fibrolytic enzymes on the lactational performance of dairy cows. J. Dairy Sci. 82: 611-617. [ Links ]
14. López SMA, Arellano GE, Barreras SA, González VV, García D, Plascencia JA, Zinn R (2006) Influencia de una enzima fibrolítica exógena y el proceso de maceración en un forraje de baja calidad sobre digestión y función ruminal en vacas Holstein secas. Vet. Mex. 37: 275-289. [ Links ]
15. Mertens DR, Ely LO (1982) Relationship of rate and extent of digestion to forage utilization. A dynamic model evaluation. J. Anim. Sci. 54: 895-905. [ Links ]
16. Nsereko VL, Morgavi DP, Rode LM, Beauchemin KA, McAllister TA (2000) Effects of fungal enzyme preparations on hydrolysis and subsequent degradation of alfalfa hay fiber by mixed rumen microorganisms in vitro. Anim. Feed Sci. Technol. 88: 153-170. [ Links ]
17. Pinos-Rodríguez JM, González SS, Mendoza GD, Bárcena R, Cobos MA (2001) efecto de enzimas fibrolíticas glucosiladas en la digestibilidad in vitro de MS y MO de alfalfa (Medicago sativa) y ballico (Lolium perenne). Rev. Cient. FCV-LUZ 11: 505-509. [ Links ]
18. Pinos-Rodríguez JM, González SS, Mendoza M, Bárcena R, Cobos M (2002a) Efecto de enzimas fibrolíticas exógenas en la digestibilidad in vitro de la pared celular de heno de alfalfa (Medicago sativa) o de ballico (Lolium perenne). Interciencia 27: 28-32. [ Links ]
19. Pinos-Rodríguez JM, González SS, Mendoza GD, Bárcena R, Cobos MA, Hernández A, Ortega ME (2002b) Effect of exogenous fibrolytic enzyme on ruminal fermentation and digestibility of alfalfa and rye-grass hay fed to lambs. J. Anim. Sci. 80: 3016-3020. [ Links ]
20. Pinos-Rodríguez JM, González S, Mendoza G, García JC, Miranda L, De La Cruz GA, De Lerma V (2005) Efecto de enzimas fibrolíticas exógenas en la degradación in vitro de ingredientes alimenticios, y en la producción de leche de vacas Holstein. Interciencia 30: 752-757. [ Links ]
21. Rode LM, Yang WZ, Beauchemin KA (1999) Fibrolytic enzyme supplements for dairy cows in early lactation. J. Dairy Sci. 82: 2121-2126. [ Links ]
22. Rooney LW, Pflugfelder RB (1986) Factors affecting starch digestibility with special emphasis on sorghum and corn. J. Anim. Sci. 63: 1607-1617. [ Links ]
23. SAS (1999) SAS Users Guide. Version 8. SAS Institute Inc. Cary, NC, EEUU. 956 pp. [ Links ]
24. Sheperd AC, Kung LJr (1996) Effects of an enzyme additive on composition of corn silage ensiled at various stages of maturity. J. Dairy Sci. 19: 1767-1773. [ Links ]
25. Steel GR, Torrie JH (1986) Bioestadística: Principios y Procedimientos. 2a ed. McGraw-Hill. México. pp. 167-171. [ Links ]
26. Van Soest PJ, Robertson JB, Lewis BA (1991) Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74: 3583-3592. [ Links ]
27. Yang WZ, Beauchemin KA, Rode LM (2000) A comparison of methods of adding fibrolytic enzymes to lactacting cow diets. J. Dairy Sci. 83: 2512–2520. [ Links ]
