











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.34 n.10 Caracas oct. 2009
Análisis de la variación espacio-temporal de la capturabilidad de la flota artesanal de caleta peñuelas que captura mesodesma donacium en Bahía de Coquimbo, Chile
Javier E. Chávez V. y Eduardo P. Pérez E.
Javier E. Chávez V. Biólogo Universidad Católica del Norte (UCN), Chile. Profesional de Pesquerías, Subsecretaría de Pesca, Coquimbo, Chile. e-mail: jchavez@subpesca.cl
Eduardo P. Pérez E. Biólogo Marino, UCN, Chile. M.Sc. y Ph.D., Cinvestav, México. Profesor, UCN, Chile. Investigador, Centro de Estudios Avanzados en Zonas Áridas (CEAZA), Chile. Dirección: UCN, Larrondo 281, Casilla 117 Coquimbo, Chile. e-mail: eperez@ucn.cl
RESUMEN
Recientemente se mostró evidencia de variación temporal del coeficiente de capturabilidad en la pesquería de Mesodesma donacium ("macha") en la zona Norte de Chile, relación previamente descrita solo para pesquerías pelágicas y demersales. Dada la heterogeneidad en la distribución espacial de la "macha" entre zonas de pesca, este trabajo exploró la existencia de variabilidad conjunta espacio-temporal en el coeficiente de capturabilidad en la flota de pesca asociada a la caleta de Peñuelas. En este caso la flota está definida en términos de la operación de buzos individuales. La capturabilidad fue calculada semanalmente mediante la ecuación de Baranov y una relación entre captura por unidad de esfuerzo (CPUE medida como kg por hora de buceo) y capturabilidad usando un modelo potencial fue utilizada para probar la hipótesis de variabilidad conjunta. Los resultados evidenciaron que la dinámica de las CPUE y de la capturabilidad mostró trayectorias inversas, representada por una relación potencial entre las variables. De la misma manera, se reafirma la existencia de variabilidad temporal de la capturabilidad, no así variabilidad espacial, en la pesquería del recurso M. donacium operado por la flota de Peñuelas en el banco de bahía Coquimbo, Chile. Se presenta una posible explicación a la relación inversa entre CPUE y capturabilidad en este tipo de recursos.
Analysis of space and time catchability variation in the artisanal fleet of peñuelas cove catching mesodesma donacium in Coquimbo Bay, Chile
SUMMARY
Recently, temporal variation in the catchability coefficient in the fishery of Mesodesma donacium ("macha") at the North region in Chile has been shown. This kind of relationship has been described previously only for pelagic and demersal fisheries. Given the heterogeneity in the spatial distribution of the species among different fishing grounds, this study evaluated the occurrence of spatial and temporal variations in the catchability coefficient in the fleet associated to Peñuelass cove. In this case, the fleet was defined in terms of the individual divers operation. Catchability was estimated weekly using the Baranovs equation and the relationship between catch per unit of effort (CPUE in kg per hour of diving) and catchability was evaluated to test the joint temporal and spatial variabilities. Results showed that dynamic of CPUE and catchability are inversely related, following a potential relationship between the two. Furthermore, temporal variability, but not a spatial one, was confirmed for Peñuelas´s fleet in Coquimbo bay bank. A possible explanation for these results with this kind of resource is presented.
Análise da variação espaço-temporal da capturabilidade da frota artesanal de caleta peñuelas que captura mesodesma donacium na Bahia de Coquimbo, Chile
RESUMO
Recentemente se mostrou evidência de variação temporal do coeficiente de capturabilidade na pescaria de Mesodesma donacium ("macha") na região Norte do Chile, relação previamente descrita somente para pescarias pelágicas e demersais. Devido à heterogeneidade na distribuição espacial da "macha" entre áreas de pesca, este trabalho explorou a existência de variabilidade conjunta espaço-temporal no coeficiente de capturabilidade na frota de pesca associada à caleta de Peñuelas. Neste caso a frota está definida em termos da operação de mergulhadores individuais. A capturabilidade foi calculada semanalmente mediante à equação de Baranov e uma relação entre captura por unidade de esforço (CPUE medida como kg por hora de mergulho) e capturabilidade usando um modelo potencial foi utilizada para provar a hipótese de variabilidade conjunta. Os resultados evidenciaram que a dinâmica das CPUE e da capturabilidade mostrou trajetórias inversas, representada por uma relação potencial entre as variáveis. Da mesma forma, se reafirma a existência de variabilidade temporal da capturabilidade, mas não da variabilidade espacial, na pescaria do recurso M. donacium operado pela frota de Peñuelas no banco da bahia Coquimbo, Chile. Apresenta-se uma possível explicação da relação inversa entre CPUE e capturabilidade neste tipo de recursos.
PALABRAS CLAVE / Capturabilidad / Flota de Peñuelas / Mesodesma donacium / Variabilidad /
Recibido: 03/06/08. Modificado: 29/09/2009. Aceptado: 30/09/2009.
Introducción
La capturabilidad, que representa la proporción de individuos de un stock capturado por unidad de esfuerzo (Peterman y Steer, 1981; Gulland, 1983), es considerada como un parámetro clave en pesquerías (Arreguín-Sánchez, 1996) dado que, conociendo las CPUE es posible estimar la biomasa de una manera simple y sin necesidad de gran cantidad de información pesquera. Sin embargo, se ha demostrado la existencia de variabilidad en la capturabilidad entre algunos recursos marinos de distinta naturaleza, como pelágicos y demersales (MacCall, 1976; Peterman y Sterr, 1981; Ye y Mohammed, 1999; Pérez y Defeo, 2003; Ziegler et al., 2003; 2004) y bentónicos (Chávez, 2000; Pérez y Chávez, 2004), lo cual implica que la CPUE no podría ser utilizada como un índice de abundancia relativa y por lo tanto se limitaría su validez como insumo para el cálculo de la biomasa.
Para recursos bentónicos una de las primeras evidencias asociada a la variabilidad de la capturabilidad son los trabajos de Chávez (2000) y Pérez y Chávez (2004), quienes mostraron la variación temporal del coeficiente de capturabilidad de la flota artesanal que explota el bivalvo Mesodesma donacium en la zona Centro-Norte de Chile. Este antecedente permitió extender a pesquerías basadas en recursos bentónicos lo descrito previamente para pesquerías pelágicas y demersales (MacCall, 1976; Peterman y Steer, 1981; Bannerot y Austin, 1983; Crecco y Savoy, 1985, Gordoa y Hightower, 1991, Swain et al., 2000, Pérez y Defeo, 2003; Pérez y Chávez 2004). Aunque la alta varianza asociada a los datos (Pérez y Chávez, 2004) tuvo efectos en el poder predictivo de la capturabilidad dinámica en las capturas futuras, la relación inversa entre la proporción del stock que remueve cada unidad de pesca y la abundancia, permitió alertar sobre el peligro de colapso del recurso (Pérez y Chávez, 2004; Pérez, 2005).
Por otra parte, Pérez y Defeo (2003) mostraron que para el recurso demersal Heterocarpus reedi no solo existe variabilidad temporal en el coeficiente de capturabilidad de una flota, sino que además existe variabilidad espacial. Así, al interior de la misma zona es posible esperar variabilidad, dentro y entre temporadas de pesca, asociada a la abundancia local del recurso, así cómo variabilidad de la capturabilidad entre zonas de pesca (Pérez y Defeo, 2003).Esta diferencia refleja la variabilidad espacio-temporal a nivel de macro zonas geográficas (a escala de cientos de km) o micro zonas geográficas (a escala de km).
De la misma manera que es de esperar diferencias en la distribución y abundancia espacial a macroescala de los recursos, existe una abundante literatura que da cuenta de la heterogeneidad espacial a escalas espaciales más reducidas (Caddy, 1975; Hilborn y Walters, 1987; Seijo et al., 1994; Ortiz y Stotz, 1996; Caddy y Seijo, 1998). En efecto, dentro de un mismo banco de pesca los recursos se distribuyen en parches de diferente abundancia y estructura de tamaños (Seijo et al., 1994, 1997, 2004). Autores como Caddy (1975) y Seijo et al. (2004) llamaron loci a estas unidades homogéneas que conforman una entidad heterogénea cuando se le mira a una escala espacial mayor. Así, múltiples loci constituirían un banco de pesca, con distribución espacial contagiosa, con loci que difieren en tamaño, abundancia y estructura.
Dada la evidencia de variabilidad espacial a macroescala reportada por Pérez y Defeo (2003) y dada la heterogeneidad espacial que presentan los recursos bentónicos en playas de fondos blandos (Defeo y Rueda, 2002), este trabajo explora la posibilidad de la existencia de variabilidad conjunta espacio-temporal al interior de un banco de pesca, en las denominadas "pesquería S" (sensu Orensanz et al., 2005) es decir, de pequeña escala, espacialmente estructuradas y con especies sedentarias como especie objetivo.
El análisis usa como objeto de estudio la población de M. donacium en el banco de bahía de Coquimbo, Chile, debido a la existencia de una sólida base de datos que da cuenta de la distribución espacial del recurso (Ariz et al., 1994, 1995; Ortiz y Stotz, 1996; Pérez, 1996; Arias et al., 1998), así como de los rendimientos de pesca por zona y por hora de buceo. Además, para el caso de este banco de pesca, la variabilidad temporal del coeficiente de capturabilidad es un hecho demostrado (Pérez y Chávez, 2004).
Materiales y Métodos
Antecedentes de la pesquería
El área de estudio fue la Bahía de Coquimbo, Chile (Figura 1), la cual cuenta con una playa de tipo disipativa, caracterizada por una pendiente suave y una amplia zona de barrido de la ola (Defeo y MacLachlan, 2005; Defeo et al., 2009). La captura se realiza mediante buceo semi autónomo (hooka) por uno o dos buzos operando desde una embarcación de madera con motor fuera de borda. El área de distribución del recurso en el banco es de ~15´106m2 (Ariz et al., 1994; Pérez, 1996; Chávez, 2000). La intensidad de pesca es espacialmente heterogénea (Pérez, 1996) y diferentes áreas de pesca son diferenciadas por los pescadores en función de hitos en la costa. El método de operación de los buzos consiste en la recolección de individuos de M. donacium mediante una bolsa colectora tubular (chinguillo) que se extiende sobre el fondo en la medida que el buzo avanza barriendo el fondo. Una vez que la bolsa está completamente extendida, el buzo regresa a su posición inicial y repite el proceso en otra dirección. La colecta continúa hasta que el buzo completa la cuota de captura diaria.

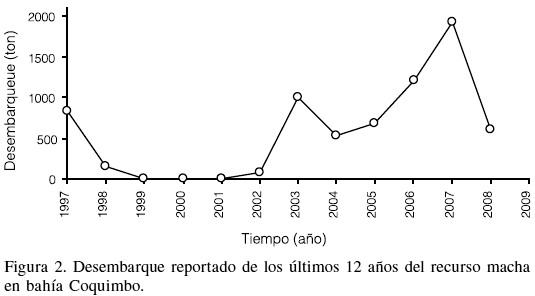
Las capturas de los últimos 12 años del recurso M. donacium en bahía Coquimbo, han mostrado una variabilidad importante, con un decaimiento progresivo desde 1997 al 2001 y una posterior recuperación a partir del 2002 con un máximo el 2007 de 1925 toneladas (Figura 2).
En este tipo de recursos, la dinámica de la abundancia y la estructura espacial, está fuertemente influenciada por el transporte vertical de arena y la migración de reclutas al submareal (Sánchez et al., 1982). En efecto, en verano la playa experimenta su mayor acreción, mientras que en invierno ocurre un proceso de erosión (Jaramillo, 1978). Ortiz y Stotz (1996) no encontraron evidencia de reclutamiento de M. donacium en el submareal, mientras que reportaron las intensidades de reclutamiento más altas a 0-1m de profundidad. Así, el proceso de acreción-erosión, característico de playas de arena permite el transporte de los juveniles desde el intermareal al submareal, donde está presente la fracción adulta de la población sujeta a presión de pesca por parte de los buzos. En el submareal los juveniles crecen hasta el estado adulto, formando capas de individuos de distinto grosor. Así, en parches de altas densidades es usual encontrar grupos de individuos que se distribuyen en tres dimensiones; es decir, en parches de un largo y ancho variable y en varias capas de profundidad en la arena. En contraste, en parches de menor densidad la distribución tiende a ser bidimensional, con una capa única de ejemplares.
Estimaciones de capturabilidad
La información utilizada fueron datos diarios de área barrida por los buzos, horas efectivas de pesca y captura tomadas entre diciembre 1996 y marzo 1998.
La capturabilidad fue calculada semanalmente usando la ecuación propuesta por Baranov (1918)
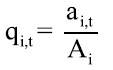 (1)
(1)
donde ai,t: área barrida por hora de buceo en la zona i al tiempo t, y A: área de distribución del recurso en la zona i (parche). Caddy (1975) y Seijo et al. (1994) recomendaron esta ecuación como una herramienta de cálculo útil para pesquerías de recursos bentónicos. De acuerdo con esto, el valor de q estaría dado en función de lo propuesto por Pérez (1996), quien definió el área de pesca barrida por un buzo como
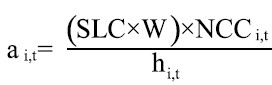 (2)
(2)
donde SLC: longitud del chinguillo, W: ancho de la franja de arrastre en metros (W= 1m; Pérez, 1996), NCCi,t: número de cambios de posición que realiza el buzo con el chinguillo en la zona i al tiempo t, y hi,t: horas de buceo en la zona i al tiempo t. Los datos requeridos para los cálculos fueron obtenidos en coordinación con los buzos, acordando el diseño para la toma de la información. El área de distribución de el recurso en cada zona de pesca (A) fue tomada desde Ariz et al. (1994), Chávez (2000), y Pérez y Chávez (2004).
Un requisito importante para la validez en la estimación de la capturabilidad es que ésta sea calculada en forma independiente de la biomasa o de la CPUE (Seijo et al., 1997). Sin embargo, a partir de la Ec. 1 es factible suponer que el área barrida aumenta en forma proporcional con el esfuerzo (medido como horas de buceo), de manera tal que la estimación del coeficiente de capturabilidad se hace dependiente del esfuerzo y por lo tanto también de la CPUE. Con el fin de evaluar esta posibilidad, se relacionó el esfuerzo (horas de buceo) con el área barrida por el buzo para cada zona de pesca, habiendo tres alternativas posibles: que ambas variables se relacionen en forma positiva, negativa o que no exista relación. Para que efectivamente el área barrida dependa del esfuerzo, se debe obtener que la relación sea positiva y significativa; esto es, que a más horas de buceo, mayor sea el área barrida por el buzo. De no existir relación significativa implica que el área barrida es independiente del esfuerzo, medido como horas de buceo efectivo. Así, con la información disponible sería posible calcular un valor confiable de q a través del tiempo en forma independiente al de las CPUE o de biomasa, si se rechaza la hipótesis de correlación entre horas de buceo y área barrida.
La relación entre la CPUE estandarizada (kg·hr-1) y la capturabilidad (hr-1) fue ajustada mediante un modelo potencial que brindó el coeficiente de determinación más alto
![]() (3)
(3)
donde los subíndices i,t representan a la zona de pesca i en el tiempo t. Para evaluar la significancia de la regresión, se usó un test de ANOVA. Este análisis es importante ya que de existir significancia implica que, dado que cada CPUE está indexada en tiempos distintos, existen diferencia temporales en el análisis. Para comparar la tasa de cambio de la capturabilidad respecto de la CPUE entre distintas zonas de pesca se realizó un análisis de covarianza ANCOVA (Zar, 1999).
Resultados
Relación entre esfuerzo y área barrida
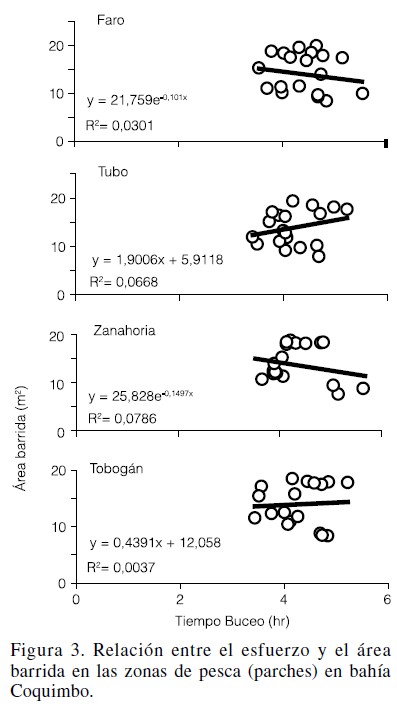
En las zonas de pesca Faro y Zanahoria, los ajustes fueron explicados por modelos exponenciales, mientras que en las zonas Tubo y Tobogán, fue un modelo lineal el que proveyó el mejor ajuste aunque en todos los casos, con un valor bajo en el coeficiente de determinación. De esta manera, en todas las zonas de pesca la pendiente de la relación no fue significativamente distinta de cero, no observándose relación entre el esfuerzo y el área barrida (p>0,259; Figura 3).
Variabilidad temporal entre CPUE y capturabilidad
La dinámica de la CPUE mostró una disminución a través del tiempo (Figura 4) en un intervalo entre 35 y 16kg/hr, mientras que la capturabilidad, mostró la tendencia inversa (Figura 5) con valores entre 1,5´10-5 y 3,9´10-5hr-1. En las cuatro zonas de estudios la relación entre CPUE y q fue significativa (p<0,00057), de tal forma que se observan diferencias en el valor semanal de ambas variables.
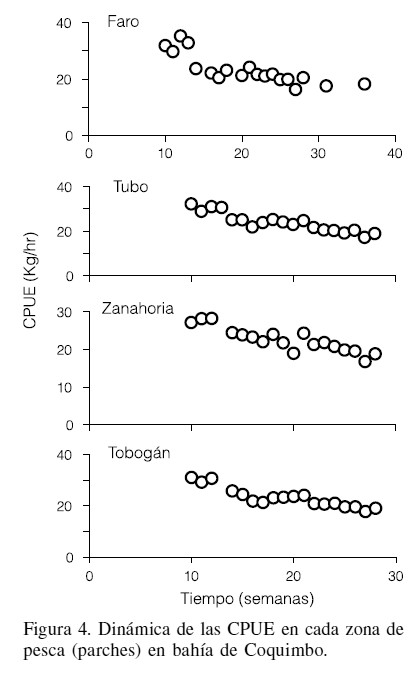
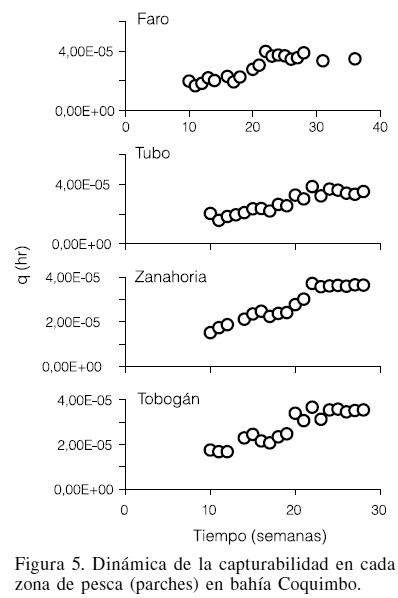
Variabilidad espacial entre CPUE y capturabilidad
Los valores del coeficiente de determinación mostraron una explicación entre el 51 y 71% de la varianza total; mientras que el valor de las pendientes se observó dentro del intervalo de -1,68 a -1,00 (Figura 6), indicando que a medida que disminuye la CPUE la capturabilidad de la flota aumenta. La mayor tasa de cambio se asoció a la zona de Zanahoria, en tanto que la menor fue la del Faro.
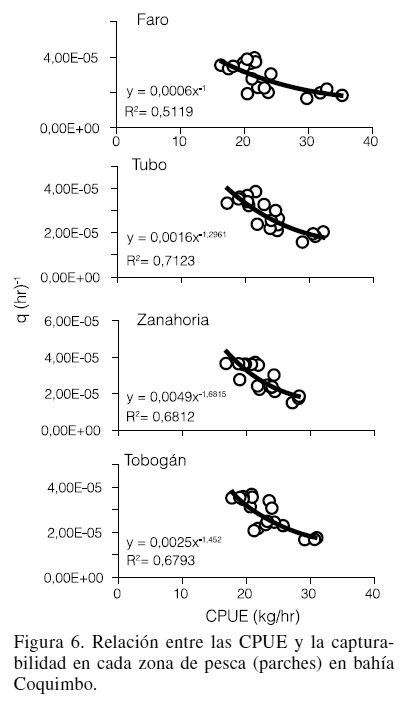
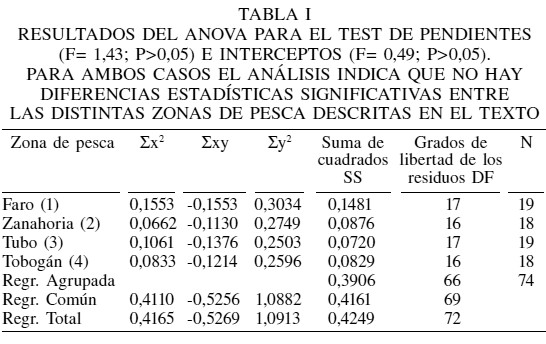
El ANCOVA no mostró diferencias significativas en las pendientes (Tabla I, F= 1,43; p>0,05), pudiéndose calcular una pendiente común de -1,26. De la misma manera, no se encontraron diferencias estadísticamente significativas en los interceptos (Tabla I, F= 0,49; p>0,05), lo que evidencia que corresponde a sitios de pesca con niveles de abundancia similares.
Discusión
Intuitivamente cabe pensar que el área barrida es función de las horas de buceo, que es la unidad de esfuerzo para la medición de la CPUE. Así, al relacionar la capturabilidad con la CPUE se produciría una correlación de variables que no son independientes entre sí. Sin embargo, dado que la pendiente de la relación esfuerzo vs área barrida no difiere significativamente de cero, se puede concluir que en todas las zonas de estudio no hubo relación entre las variables; es decir, el área barrida no es función del tiempo de buceo. Por lo tanto, la estimación de capturabilidad cumple con el requisito fundamental de ser estimada con independencia absoluta de datos de CPUE o de biomasa.
La falta de correlación entre las horas de buceo y el área barrida por el buzo, podría ser explicada porque cuando el buzo encuentra un parche de alta abundancia, se concentra en obtener su captura en un área relativamente pequeña, ocupando bastante tiempo en conseguir una captura deseable. Esto es posible dado que el recurso, cuando es abundante, tiende a concentrarse en capas; es decir, se incorpora una nueva dimensión a la distribución. En este caso, la extracción del recurso hecha por vía manual tomaría tiempo al ser necesario excavar. Por el contrario, cuando las abundancias locales disminuyen, el recurso tiende a distribuirse en una sola capa y así el buzo tendrá que cubrir distancias lineales mucho mayores para obtener las mismas capturas, incluso en un tiempo equivalente al empleado por un buzo en un parche de tres dimensiones. En consecuencia en ambas situaciones los tiempos de buceo pueden ser similares mientras áreas barridas son diferentes, o bien distintas horas de buceo pueden generar áreas barridas equivalentes. Así, los resultados explicarían que el área barrida corresponde a una medida independiente del esfuerzo de pesca medido en tiempo de buceo.
El mecanismo anterior difiere del descrito para otro tipo de recursos, como por ejemplo para recursos pelágicos menores. Aumentos en la capturabilidad producto de la reducción en la población son propios de pequeños pelágicos, producto de su notable efecto de concentración que los hacen mas fáciles de capturar (MacCall, 1976; Csirke y Gummy, 1996). Sin embargo, en el caso de recursos sedentarios existe un cambio en la forma de operar del arte de pesca (el buzo) y no en la conducta del recurso, dada su baja o nula capacidad de movimiento de la fracción explotable. Así, ante la perspectiva de una disminución en la tasa de captura se barre un área mayor respecto del área total de distribución del recurso (ver discusión en Pérez y Chávez, 2004).
De acuerdo a lo esperado, la CPUE decayó en el tiempo, lo cual se explicaría por una disminución de las abundancias locales (Hilborn y Walters, 1987; Pérez y Chávez, 2004), mientras que la capturabilidad fue aumentando en forma inversa a la disminución de la CPUE. El análisis de la relación entre CPUE y capturabilidad mostró que en todas las zonas de pesca las pendientes fueron estadísticamente distintas de cero. El que las parejas de datos de CPUE y capturabilidad estén indexadas en el tiempo (Ec. 3) y que exista significancia en la regresión, implica que hay una diferencia temporal implícita. En efecto, desde el punto de vista estadístico, la significancia de una regresión establece diferencias entre las múltiples parejas de datos. De esta forma, la significancia entre las variables es indicador de diferencias temporales entre ellas.
El análisis espacial indicó que no existe variabilidad en la capturabilidad entre zonas de pesca y que en términos de abundancia las zonas de pesca son similares. En efecto, el hecho que las pendientes de las relaciones entre CPUE y capturabilidad sean paralelas entre las zonas evaluadas indica que las tasas de cambio de q respecto de la CPUE son estadísticamente iguales entre zonas de pesca. De la misma manera, al ser los interceptos no significativos indican que las abundancias locales de los parches analizados en el banco de M. donacium en bahía de Coquimbo son similares.
El banco de M. donacium en bahía de Coquimbo está compuesto de un mosaico de parches, producto de la distribución espacial heterogénea del recurso (Ortiz y Stotz, 1996; Pérez, 1996). En términos de la pregunta original, si hay variación espacio-temporal del coeficiente de capturabilidad entre tales parches, los resultados sugieren que tal variación conjunta no existe; solo existe variabilidad temporal, no espacial. Esto confirma la variabilidad temporal reportada por Pérez y Chávez (2004) para esta misma localidad. En este sentido, los buzos mantienen una efectividad similar, con independencia del parche que explotan.
Pérez y Chávez (2004) aplicaron un modelo de remoción dinámico modificado para explicar el comportamiento de las capturas en el banco de machas de bahía de Coquimbo, usando un coeficiente de capturabilidad variable. En esa línea de investigación, los resultados obtenidos en este trabajo indican que para el banco en su totalidad se puede utilizar solo un par de parámetros para modelar convenientemente la dinámica de la capturabilidad, no siendo necesario un cálculo independiente para cada área de pesca.
Finalmente, los resultados reafirman la limitación que tiene la CPUE como un indicador de abundancia relativa (Ulltang, 1976), particularmente para especies sedentarias (Seijo et al., 1997; Pérez y Defeo, 2003; Pérez y Chávez, 2004) debido a los cambios en la capturabilidad. Estos autores han sugerido que para tener estimados de biomasa a partir de la CPUE observada, ésta debiera estar ponderada por la Ec. 3 (Pérez y Chávez, 2004). De esta manera se obtendrían valores más confiables de biomasa (ver discusión de Pérez y Chávez, 2004).
Referencias
1. Arias JL, Arias E, Defeo O, Pérez E, Stotz W, Valdebenito M (1998) Caracterización Bioeconómica de la Pesquería del Recurso Macha en la Zona Norte y Centro-sur. Proyecto FIP Nº 95-24. Coquimbo, Chile. 113 pp. [ Links ]
2. Ariz L, Jerez G, Pérez E, Potocnjack C (1994) Bases para la Ordenación y Desarrollo de las Pesquerías Artesanales del Recurso Macha (M. donacium) en Chile Central. Informe Final. Instituto de Fomento Pesquero. Valparaíso, Chile. 61 pp. [ Links ]
3. Ariz L, Brown D, Jerez G, Miranda H, Roa R (1995) Monitoreo de las Pesquerías de Machas en la IV Región (Peñuelas-Coquimbo) y en la Zona Norte de la V Región (Longotoma y Ritoque). Instituto de Fomento Pesquero. Valparaíso, Chile. 107 pp. [ Links ]
4. Arreguin-Sánchez F (1996) Catchability: a Key parameter for fish stock assessment. Rev. Fish Biol. Fisher. 6: 221-242. [ Links ]
5. Bannerot SP, Austin CB (1983) Using frequency distribution of catch per unit fishing effort to measure fish-stock abundance. Trans. Am. Fish. Soc. 112: 608-617. [ Links ]
6. Baranov TY (1918) On the question of the biological basis of fisheries. Proc. Inst. Icht. Inv. 1: 81-128. [ Links ]
7. Caddy JF (1975) Spatial models for an exploited shellfish population, and its application to Georges Bank scallop fishery. J. Fish. Res. Board Can. 32: 1305-1328. [ Links ]
8. Caddy JF, Seijo JC (1998) Application of a spatial model to explore rotating harvest strategies for sedentary species. Can. Spec. Publ. Fish. Aquat. Sci. 125: 359-365. [ Links ]
9. Csirke J, Gumy AA (1996) Análisis bioeconómico de la pesquería pelágica peruana dedicada a la producción de harina y aceite de pescado. Bol. Inst. Mar Perú 15: 25-68. [ Links ]
10. Chávez J (2000) Análisis Dinámico del Coeficiente de Capturabilidad y sus Implicancias en la Modelación de Pesquerías: Mesodesma donacium en el Banco de Bahía Coquimbo, un Estudio de Caso. Tesis. Universidad Católica del Norte. Chile. 90 pp. [ Links ]
11. Crecco VA, Savoy TF (1985) Density-dependent catchability and its potencial causes and consequences on Connecticut River American Shad, Alosa sapidissima. Can. J. Fish. Aquat. Sci. 42: 1649-1657. [ Links ]
12. Defeo O, McLachlan A (2005) Patterns, processes and regulatory mechanism in sandy beach macrofauna: a multi-scale analysis. Mar. Ecol. Prog. Ser. 295: 1-20. [ Links ]
13. Defeo O, Rueda M (2002) Spatial structure, sampling design and abundance estimates in sandy beach macroinfauna: some warnings and new perspectives. Mar Biol. 140: 1215-1225. [ Links ]
14. Defeo O, McLachlan A, Schoeman DS, Schlacher TA, Dugan J, Jones A, Lastra M, Scapini F (2009) Threats to sandy beach ecosystems: A review. Estuar. Coast. Shelf Sci. 81: 1-12. [ Links ]
15. Gordoa A, Hightower JE (1991) Changes in catchability in a bottom-trawl fishery for Cape Hake (Merluccius capensis). Can. J. Fish. Aquat. Sci. 48: 1887-1895. [ Links ]
16. Gulland JA (1983) Fish Stock Assessement: A Manual for Basic Methods. Wiley. Nueva York, EEUU. 223 pp. [ Links ]
17. Hilborn RF, Walters CJ (1987) A general for simulation of stock and fleet dynamics in spatially heterogeneus fisheries. Can. J. Fish. Aquat. Sci. 44: 1366-1369. [ Links ]
18. Jaramillo E (1978) Zonación y estructura de la comunidad macrofaunística en playas de arena del sur de Chile (Mehuin, Valdivia). Stud. Neotrop. Fauna Env. 13: 17-92. [ Links ]
19. MacCall AD (1976) Density-dependence of catchability coefficient in the California Pacific sardine, Sardinops sagax caerulea, purse seine fishery. Calif. Coop. Ocean. Fish. Inv. Rep. 18: 136-148. [ Links ]
20. Orensanz J M, Parma A M, Jerez G, Barahona N, Montecinos M, Elias I (2005) What are the key elements for the sustainability of "S-fisheries"? Insights from South America. Bull. Mar. Sc. 76: 527-556. [ Links ]
21. Ortiz M, Stotz W (1996) Distribución de juveniles recientemente asentados de Mesodesma donacium (Lamarck, 1818) (Mollusca: Bivalvia: Mesodesmatidae) en tres bahías de la cuarta región: Variables físicas y químicas que le caracterizan. Biol. Pesq. 25: 27-40. [ Links ]
22. Pérez E P (1996) Análisis de la Pesquería de Mesodesma donacium en el Banco de Peñuelas (Chile, IV región), bajo Condiciones de Riesgo e Incertidumbre. Tesis. CINVESTAV-IPN. Mérida, Mexico. 82 pp. [ Links ]
23 Pérez EP (2005) Un modelo simple para describir la dinámica de la biomasa del camarón nailon Heterocarpus reedi en Coquimbo, Chile. Invest. Mar, Valparaíso 33: 131-142. [ Links ]
24. Pérez EP, Chávez J (2004) Modelling short-term dynamic behavior of the surf clam (Mesodesma donacium) fishery in northern Chile using static and dynamic catchability hypotheses. Interciencia 29: 193-198. [ Links ]
25. Pérez EP, Defeo O (2003) Time-space variation in the catchability coefficient as a function of catch per unit of effort in Heterocarpus reedi (Decapoda, Pandalidae) in North-Central Chile. Interciencia 28: 178-182. [ Links ]
26. Peterman R M, Steer G J (1981) Relation between sportfishing catchability coefficients and salmon abundance. Trans. Am. Fish. Soc. 114: 436-440. [ Links ]
27. Sánchez M, Castilla JC, Mena O (1982) Variaciones verano-invierno de la macrofauna de arena en playa Morrillos (Norte Chico, Chile). Stud. Neotrop. Fauna Env. 17: 31-49. [ Links ]
28. Seijo JC, Caddy JF, Euán J (1994) SPATIAL: Space-Time Dynamics in Marine Fisheries. A Software Package for Sedentary Species. FAO. Comp. Inf. Ser. Fish. 6. 116 pp. [ Links ]
29. Seijo JC, Defeo O, Salas S (1997) Bioeconomía Pesquera: Teoría, Modelación y Manejo. Documento técnico de pesca. Nº 368. FAO. Roma, Italia. 176 pp. [ Links ]
30. Seijo JC, Pérez EP, Caddy JF (2004) A simple approach for dealing with dynamics and uncertainty in fisheries with heterogeneous resource and effort distribution. Mar. Freshw. Res.. 55: 249-256. [ Links ]
31. Swain DP, Poirier GA, Sinclair AF (2000) Effect of water temperature on catchability of Atlantic cod (Gadus morhua) to the bottom-trawl survey in the southern Gulf of St. Lawrence. ICES J. Mar. Sci. 57: 56-68. [ Links ]
32. Ulltang ø (1976) Catch per unit of effort in the Norwegian purse seine fishery for Atlanto-Scandinavian herring. FAO Fish. Tech. Pap. 155: 91-101. [ Links ]
33. Ye Y, Mohammed H (1999) An analysis of variation in catchability of green tiger prawn, Penaeus semisulcatus in waters off Kuwait. Fish. Bull. 97: 702-712. [ Links ]
34. Zar J (1999) Biostatistical Analysis. 3ª ed. Prentice-Hall. Englewood Cliffs, NJ, EEUU. 718 pp. [ Links ]
35. Ziegler PE, Frusher SD, Johnson CR (2003) Space-time variations in catchability of southern rock lobster Jasus edwardsii in Tasmania explained by environmental, physiological and density-dependent processes. Fish. Res. 61: 107-123. [ Links ]
36. Ziegler PE, Haddon M, Frusher SD, Johnson CR (2004) Modelling seasonal catchability of the southern rock lobster Jasus edwardsii by water temperature, moulting and mating. Mar. Biol. 145: 179-190. [ Links ]
