











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkInterciencia
versión impresa ISSN 0378-1844
INCI v.34 n.12 Caracas dic. 2009
Patrón de variación axial y radial del peso específico en la madera de erisma uncinatum warm.
Jesús Velásquez, María E. Toro, Luís Gómez, Frank M. Terzo y Arlenys Márquez
Jesús Velásquez
. Ingeniero Industrial Forestal, Universidad Nacional Experimental de Guayana (UNEG), Venezuela. MSc. Tecnología de Productos Forestales, Universidad de los Andes (ULA), Venezuela. Docente-Investigador, UNEG, Venezuela. Dirección: Laboratorio de Biotecnología de la Madera, CEBIOTEG-UNEG, Venezuela. e-mail: jvelasquez@uneg.edu.veMaría E. Toro. Ingeniera Industrial Forestal, UNEG, Venezuela. M.Sc. en Tecnología de Productos Forestales, ULA, Venezuela. Docente Investigadora, UNEG, Venezuela.
Luis Gómez. Ingeniero Industrial Forestal, UNEG, Venezuela. M.Sc. en Tecnología de Productos Forestales, ULA, Venezuela. Docente Investigador, UNEG, Venezuela.
Frank M. Terzo. Ingeniero Agrónomo, Universidad de Oriente, Venezuela. M.Sc. en Ciencias Ambientales, Universidad Nacional Rómulo Gallegos, Venezuela. Docente Investigador, UNEG, Venezuela.
Arlenys Márquez, Ingeniero Industrial Forestal, UNEG, Venezuela. M.Sc. en Tecnología de Productos Forestales, ULA, Venezuela. Docente Investigador, UNEG, Venezuela.
RESUMEN
La variabilidad natural de la madera es producto de su heterogeneidad celular, evidenciada tanto en su anatomía como en su composición química, las cuales a su vez, se reflejan en sus propiedades físicas y mecánicas. Se realizó la caracterización del patrón de variación axial y radial del peso específico seco en la madera de Erisma uncinatum (mureillo), procedente de la región Guayana, Venezuela. La evaluación se realizó empleando una muestra de seis árboles. Las muestras de madera se tomaron a tres niveles de altura (1,30m, 30% y 60% de la altura comercial). La evaluación radial se realizó sobre cinco puntos equidistantes en la dirección radial médula-corteza (0, 25, 50, 75, 90%). El análisis estadístico señaló que la variación entre árboles no fue estadísticamente significativa en comparación con el efecto distancia radial dentro de cada individuo. El peso específico de la madera, presentó un valor promedio de 0,52 ±0,7. La variación radial mostró un incremento lineal positivo médula-corteza (R2=0,68), con una variación de 22%, evidenciándose una demarcación preliminar entre madera juvenil y adulta, a partir del 50% de la dirección de los radios celulares. La variación axial disminuye con la altura del árbol y es menos consistente que la radial, apenas un 13%. La edad fisiológica de las células del cambium vascular es un factor importante que determina la variación del peso específico en la madera de E. uncinatum.
Axial and radial variation pattern in the specific gravity of the wood of erisma uncinatum warm
SUMMARY
The natural variability of wood is a product of its cellular heterogeneity, evidenced in its anatomy as well as in its chemical composition, which in turn are reflected upon its physical and mechanical properties. A characterization of the axial and radial variation pattern in the dry specific gravity was carried out on wood of the species Erisma uncinatum ("mureillo"), from a forest in the Guayana region, Venezuela. The evaluation was carried out using a sample of six trees. Samples were taken at three different height levels (1.30m, 30% and 60% of commercial height). The radial evaluation was performed on five equidistant points in a pith-bark radial direction (0, 25, 50, 75, 90%). The statistical analysis indicated that the variation between trees was not statistically significant compared to the effect of radial distance in each individual. The specific gravity of the wood presented a mean value of 0.52 ±0.7. The radial variation showed a positive linear increase from pith to bark (R2=0.68), with a 22% variation, evidencing a preliminary boundary between juvenile and adult wood, beginning at 50% of the direction of cellular radii. The axial variation decreases with tree height and is less consistent than the radial variation, barely 13%. The physiological age of the vascular cambium is an important factor that determines the variation in the specific gravity of Erisma uncinatum wood.
Padrão de variação axial e radial do peso específico na madeira de erisma uncinatum warm
RESUMO
A variabilidade natural da madeira é produto de sua heterogeneidade celular, evidenciada tanto em sua anatomia como em sua composição química, as quais por sua vez, são refletidas em suas propriedades físicas e mecânicas. Realizou-se a caracterização do padrão de variação axial e radial
do peso específico na madeira de Erisma uncinatum (cedrinho), procedente da região Guayana, Venezuela. A avaliação foi realizada empregando uma amostra de seis árvores. As amostras de madeira foram colhidas em três níveis de altura (1,30m, 30% e 60% da altura comercial). A avaliação radial foi realizada sobre cinco pontos equidistantes na direção radial medula-crosta (0, 25, 50, 75, 90%). A análise estatística apontou que a variação entre árvores não foi estatisticamente significativa em comparação com o efeito distância radial dentro de cada indivíduo. O peso específico na madeira, apresentou um valor médio de 0,52 ±0,7. A variação radial mostrou um incremento linear positivo medula-crosta (R2=0,68), com uma variação de 22%, evidenciando-se uma demarcação preliminar entre madeira juvenil e adulta, a partir do 50% da direção dos raios celulares. A variação axial diminui com a altura da árvore e é menos consistente que a radial, apenas em 13%. A idade fisiológica das células do câmbio vascular é um fator importante que determina a variação do peso específico na madeira de E. uncinatum.PALABRAS CLAVE / Erisma uncinatum / Peso Específico de la Madera / Variabilidad de la Madera /
Recibido: 02/10/2008. Modificado: 21/10/2009. Aceptado: 22/10/2009.
Introducción
Existen dos fuentes de variación respecto a las propiedades y características de la madera en el árbol, las variaciones entre árboles y las variaciones dentro del árbol (Giménez y López, 2000; Igartúa et al., 2003). Las primeras vienen explicadas por causas genéticas, factores ecológicos y gestión de la masa, mientras que las segundas están condicionadas por su situación dentro del árbol, específicamente relacionadas con variabilidad dentro del incremento de crecimiento, variabilidad producida de médula a corteza (variabilidad radial) y variabilidad ocasionada por diferentes niveles de altura (variabilidad longitudinal; Wang et al., 2000; Savva et al., 2002; Espinoza, 2004). Dado que la mayoría de las veces la variación dentro del árbol es superior a la variación entre árboles, es imprescindible cuantificarla y definirla de la forma más precisa posible, puesto que va a condicionar los productos que se pueden obtener al utilizar este recurso (Zobel y Van Buijtenen, 1989; García et al., 2003).
El modelo de variación radial de la densidad y el peso específico dentro del árbol muestra diferentes patrones de variación, que van desde una relación lineal altamente significativa hasta relaciones lineales negativas con la edad fisiológica de las células del cambium vascular (Zobel y Van Buijtenen, 1989). En cambio, en el sentido axial, las variaciones son en general menos consistentes (Wilken, 1988; León y Espinoza, 1999; Giménez y López, 2000).
Wiemann y Williamson (1988) determinaron un incremento lineal positivo del peso específico desde la médula a la corteza, asociado a estrategias de crecimiento en tres ecosistemas en Costa Rica, especies de bosque seco donde el incremento fue de 20-80%, en bosque montano de 20-40% y en bosques húmedos de 20-270%. Butterfield et al. (1993), igualmente en Costa Rica, al evaluar el peso específico en las especies Hyeronima alchorneoides y Vochysia gatemalensis, determinaron para ambas el mismo patrón de variación, incremento médula-corteza, desde 155 hasta 300% del valor inicial. En el Amazonas peruano, Woodcock et al. (2000) determinaron en especies latifoliadas un incremento de 10-40%. Parolin (2002) evaluó en el Amazonas brasilero 32 especies latifoliadas y determinó incrementos lineales positivos del peso específico de 10-60%, siendo los mayores incrementos asociados a especies pioneras en la zona de evaluación.
Contrariamente, en Quercus garryana procedente de EEUU Lei et al. (1996) concluyeron que para esta especie el peso específico declinó linealmente desde la médula a la corteza en un 27%, y desde la altura de pecho hasta el ápice del árbol. En Canadá, Yanchuk y Micko (1990) no determinaron un patrón característico para la madera de Populus tremuloides, ya que muchos de los individuos evaluados mostraron disminución del peso específico, mientras en otros incrementó en la medida que aumentaba la distancia médula corteza.
El mureillo (Erisma uncinatum) es una de las especies forestales maderables de mayor cuota de aprovechamiento en los lotes boscosos y reservas forestales ubicadas hacia el sur de Venezuela, debido a su elevada presencia y a su reconocido valor comercial, producto de la versatilidad de su madera en relación a sus propiedades físicas y mecánicas (Moret y Ruíz, 1998). En Venezuela los niveles de extracción de la madera de esta especie alcanzan valores superiores a los 15000m3 anuales desde 1992, tasa de extracción que continúa en incremento (Ortegano et al., 2001; Silva et al., 2004; Faría et al., 2005; 2006).
La especie E. uncinatum tiene una amplia distribución natural desde los bosques húmedos tropicales hasta los bosques húmedos subtropicales presentes en Colombia, Brasil, Perú, Surinan y Venezuela (Justiniano y Fredericksen, 1999). Esta especie ha sido aprovechada comercialmente desde los años 60 en la Guayana Venezolana (Pérez, 1973) En el norte del Brasil ha sido utilizada principalmente como madera de construcción y acabados, constituyendo junto a otras dos especies (Dinizia excelsa y Goupia glabra), ~99,7 % de las maderas utilizadas (Barbosa, 1990). En la Guayana Francesa se comenzó a utilizar el E. uncinatum en los años 90 (Fouquet, 1991). En la Amazonia está considerada como la especie forestal de mejor calidad de madera dentro de la familia Vochysiaceae (Mainieri y Pérez, 1989). En el norte de Bolivia, E. uncinatum se utiliza, aunque en menor grado, como materia prima para la construcción de viviendas rústicas y galpones para el almacenamiento de castañas y corteza de palmito (Justiniano y Fredericksen,1999).
Con respecto a la especie E. uncinatum, se han realizado estudios relacionados con la descripción macroscópica y microscópica de la madera (Corothie, 1967; Quirk, 1980; Détienne y Welle, 1998), sus propiedades físicas y mecánicas (IFLA, 1992), ecología de la especie (Justiniano y Fredericksen, 1999) y determinación de la ecuación para el cálculo del volumen, tanto de madera en rolas como en madera aserrada (Moret y Ruiz, 1998). Sin embargo, antecedentes relacionados con la variabilidad de las características y propiedades físicas de la madera son limitados. En tal sentido se plantea evaluar y determinar el patrón de variación, en el sentido axial y radial, del peso específico seco de la madera, entre y dentro de individuos de una de las especies forestales maderables más importantes y de mayor consumo a nivel nacional, proveniente de la Reserva Forestal de Imataca, como lo es la especie E. uncinatum (mureillo).
Materiales y Métodos
Obtención de muestras de madera
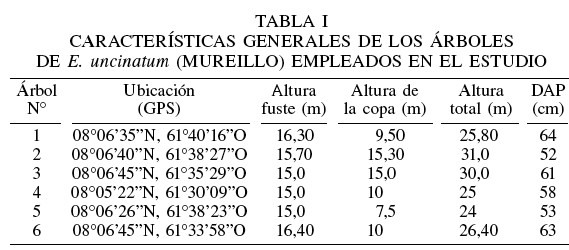
Las muestras para el estudio fueron recolectadas en el compartimiento C5 de la Unidad II en la Reserva Forestal Imataca, Estado Bolívar, Venezuela, ubicada entre 08°00 y 08°36N y entre 61°29 y 61°58O, con una altitud media de 270msnm, temperatura promedio anual de 25,4°C, y precipitación anual de 1696mm (CDF, 2001). En la zona de estudio se seleccionaron seis árboles de la especie forestal E. uncinatum, con copa ubicada en los estratos superiores o emergentes, vigorosas, fácilmente visibles, con fustes sin evidencia de enfermedad o daño, con mínima inclinación respecto a la posición vertical y con un diámetro a la altura de pecho (DAP) superior al diámetro mínimo establecido (40 cm) como apropiado para el corte en esta especie. Las características generales de los árboles seleccionados se muestran en la Tabla I.
En cada ejemplar se demarcó la orientación cardinal Norte-Sur, y para analizar la variación axial, se tomaron las muestras de acuerdo a tres niveles proporcionales a la altura comercial (1,30m, 30% y 60% de la altura comercial). De cada altura de muestreo en cada árbol se extrajo una rodaja de 10cm de espesor; las mismas fueron rotuladas y colocadas en bolsas de polietileno para evitar su desecación brusca. Para el estudio de la variación radial se emplearon las muestras de maderas tomadas a 1,30m. sobre el nivel del suelo (AP). En cada una de las rodajas se tomaron sub-muestras sobre una secuencia proporcional a la longitud de los radios desde la médula hasta la corteza, partiendo de los alrededores de la médula (0%) con incrementos radiales de 25% hasta llegar al 90% de la sección radial (0%, 25%, 50%, 75% y 90%). En cada punto de evaluación radial se elaboraron 10 probetas, lo que representó 50 probetas/altura evaluada, para un total de 150 probetas/árbol.
Determinación del peso específico
La determinación del peso específico seco se realizó según lo establecido en ASTM (1990), siguiendo lo señalado en la norma D 2395-83 y los lineamientos establecidos por Durán (1999), sobre probetas de madera con dimensiones de 2cm de aristas, libres de nudos, pudriciones, defectos y rajaduras. El peso seco se determinó colocando las probetas de madera en estufa (105 ±3ºC) hasta alcanzar peso constante. El volumen del estado seco en cada probeta se determinó por desplazamiento de agua destilada a 26ºC.
Análisis de la información
Se realizó un análisis de varianza (ANOVA) para evaluar la variación del peso específico seco de la madera individualmente, entre árboles (efecto árbol), entre las tres alturas de evaluación (efecto altura) y entre los cinco puntos de muestreo radial (efecto distancia radial; Milton y Arnold, 2004). Se aplicó un test de comparación de medias, utilizando la prueba de Games-Howell, para identificar diferencias estadísticas de la propiedad de interés dentro de los factores evaluados. La variación total del peso específico en cada altura se determinó como promedio de mediciones en cinco puntos equidistantes en la dirección médula-corteza. Para describir la relación entre la propiedad medida, como variable dependiente, y las alturas de medición como variables independientes, se utilizó un análisis de regresión lineal simple. La distribución de la normalidad y la homogeneidad residual de la varianza del peso específico para cada punto de muestreo fue verificada con las pruebas de Kolmogorov-Smirnov y el estadístico de Levene (Pardo y Ruíz, 2005). Todas las pruebas se realizaron empleando el paquete estadístico SPSS 13.0, con un nivel de confianza del 95%.
Resultados y Discusión
Variación axial
La variación axial del peso específico en la madera de E. uncinatum mostró diferencias altamente significativas entre las alturas de muestreo (Tabla II; p<0,05). La prueba de Games-Howell señaló un comportamiento particular para cada individuo bajo evaluación. En el árbol 4 se observó el menor valor de variación en el parámetro evaluado (3,8%), con ausencia de diferencias significativas entre las alturas, indicando posiblemente la formación de madera con características anatómicas más homogéneas entre la AP y el 60% de la altura comercial. En el caso particular de los árboles 1, 2 y 6, se observó ausencia de diferencias estadísticas entre el 30 y 60% de la altura comercial; sin embargo, esta fue evidente al compararla con el punto más bajo de la evaluación (AP). Contrariamente, entre los árboles 3 y 5, no se observaron diferencias en las dos primeras alturas de evaluación, la diferencia se evidenció estadísticamente con las muestras de madera obtenidas en el 60% de la altura comercial.

La agrupación y evaluación de los datos obtenidos de la variación axial del peso específico en la madera de la especie señaló que no existen diferencias estadísticas en los dos últimos estratos de evaluación (30% y 60% de la altura comercial). En cambio, la diferencia fue evidente con los datos colectados y comparados con el primer estrato de evaluación (AP). El valor promedio del peso específico seco determinado aquí para E. uncinatum fue de 0,52 (Tabla II), similar al reportado en la literatura por Vilela (1969) en muestras de madera de la especie procedentes de la Guayana venezolana (0,524). El IFLA (1992), señala un valor menor referido en este caso al peso específico básico (0,47).
Los datos de la Tabla II, muestran en general una disminución de naturaleza lineal del peso específico con la altura de muestreo. Para todos los árboles evaluados, el mayor valor siempre se determinó cercano a la altura de pecho (AP ±0,57), luego este disminuyó gradualmente hasta el extremo superior, alcanzando allí valores entre 0,45-(0,50)-0,54. En líneas generales se observó que el peso específico en AP fue en promedio 12,3% superior al correspondiente a la parte superior del árbol (60% de la altura comercial).
Los resultados del ANOVA (Tabla III) señalan que no existen diferencias significativas en los valores medios del peso específico de la madera entre los árboles bajo estudio al nivel de confianza establecido para la evaluación (95%). La falta de variabilidad de propiedades físicas, químicas y anatómicas entre árboles de la misma especie muchas veces se debe a que comparan individuos que crecen en las mismas condiciones micro-ambientales, y a que esta correlación solo será altamente significativa en aquellos casos donde se evalúan especies sometidas a diferentes condiciones y tasas de crecimiento, ubicación geográfica, manejo forestal y condiciones edafoclimáticas (Barajas, 1987; Zhang, 1995; Jorge et al., 2000; Woodcock et al., 2000; Arias y Terrazas, 2001; Gominho et al., 2001; Miranda y Pereira, 2002). La variación del peso específico en la madera de E. uncinatum encontrada en este estudio coincide con lo señalado en la bibliografía en relación a que existen mayores niveles de variación dentro de un árbol que entre árboles de la misma especie (Panshin y De Zeeuw, 1980; Lei et al., 1996; Gominho et al., 2001; Miranda y Pereira, 2002; Savva et al., 2002).

El comportamiento axial del peso específico se ajustó a un modelo de regresión lineal simple, el cual se muestra en la Figura 1 y cuya expresión fue Pe= -0,043h+0,612 (donde Pe: variable de interés y h: altura de evaluación). El análisis de regresión indicó que la altura de evaluación explica en gran medida la variación del peso específico (R2=0,68); sin embargo, existen aspectos anatómicos de la madera que repercuten sobre el valor de la variable en estudio, los cuales deben ser considerados en evaluaciones futuras para lograr una mayor comprensión de la variabilidad de la madera en la especie.
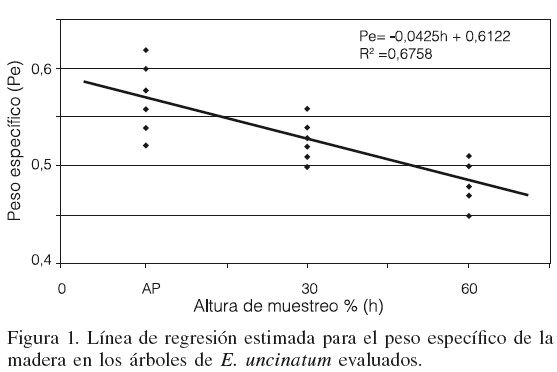
Los resultados del presente estudio revelaron una disminución significativa (a= 0,05) de la magnitud del peso específico seco con la altura en el fuste, por cuanto éste disminuye desde AP hacia el 60% de la altura comercial. Este comportamiento podría estar relacionado con las fluctuaciones que experimentan las células iniciales del cambium durante la formación de la madera en diferentes edades. Por ejemplo, algunos autores como Zobel y Van Buijtenen (1989), Tsoumis (1991), y Urbinati et al. (2003) señalaron que en árboles con edad promedio cercana a 100 años, las células más antiguas del cambium vascular se encuentran en la base del árbol, y en la medida que se produce el crecimiento en longitud, aumentando la altura del individuo, se producen células de menor edad (iniciales fusiformes jóvenes) que originan células xilemáticas con características morfológicas particulares para cada periodo de crecimiento, que repercuten significativamente sobre las propiedades físicas y mecánicas de la madera.
El perfil de variación axial del peso específico determinado en este estudio coincide con lo señalado por diversos autores en diferentes especies latifoliadas. Bosman (1996), en dos especies tropicales (Shorea leprosula y S. parvifolia), señaló que el patrón de variación fue una disminución en los primeros estratos de altura, para luego incrementarse hasta el tope del árbol, igualando en muchos casos el valor inicial cercano a la base. Nogueira et al. (2005), en un estudio sobre 310 árboles del Amazonas brasilero, señalaron que la densidad de la madera en el tope del árbol fue 8% menor que a la altura de pecho en el 87% de los árboles. Igualmente, Lei et al. (1996) encontraron una disminución del 8% del valor del peso específico con la altura de valuación en la especie Quercus garryana.
El patrón de variación axial experimentado por el peso específico de la madera en los árboles de E. uncinatum está de acuerdo con la hipótesis establecida por diversos autores quienes señalan, por un lado, que en los bosques tropicales muchas especies muestran un rápido crecimiento vertical debido a la intensa competencia por la luz solar, lo cual demanda estabilidad estructural, y por otro, que durante el desarrollo de la copa el árbol es sometido a elevados niveles de stress debido a la fuerza ejercida por el viento; ambos aspectos son contrarrestados mediante un refuerzo mecánico, por la producción de madera de mayor densidad y resistencia cercano a la base del árbol (Wiemann y Williamson, 1989; Woodcock y Shier, 2002; Sotelo et al., 2007).
Variación radial
Los resultados de las pruebas de normalidad y homogeneidad de las varianzas se muestran en la Tabla IV. Los niveles críticos asociados a los estadísticos de Kolmogorov-Smirnov y Levene en cada punto de muestreo radial señalaron que los datos obtenidos proceden de poblaciones normales y de muestras no homogéneas, lo cual permite la aplicación del ANOVA y la prueba de Games-Howell para la determinación de las diferencias estadísticas.

El análisis estadístico de la variación radial del peso específico en la madera de E. uncinatum (ANOVA), mostró diferencias altamente significativas (p<0,05), entre los diferentes puntos de muestreo en los árboles bajo estudio. Los resultados de esta evaluación se resumen en la Tabla V. En todos los árboles el peso específico fue relativamente bajo en los alrededores de la médula, con ausencia de diferencias estadísticas entre los dos primeros puntos de muestreo (0 y 25%). Sin embargo, estos mostraron diferencias altamente significativas (p<0,05) con los datos promedios reportados a partir de 50% de la sección radial. En la mayoría de los árboles, así como al considerar los datos en conjunto, se observó que los tres últimos puntos de muestreo no evidenciaron diferencias significativas en los valores obtenidos (50%, 75% y 90% de la sección radial).

En la Figura 2 se muestra gráficamente la tendencia del comportamiento radial experimentado por el peso específico en la madera de E. uncinatum. El patrón de variación general señala un incremento lineal del peso específico en la medida que aumenta la distancia desde las adyacencias de la médula hasta la corteza del los árboles evaluados.
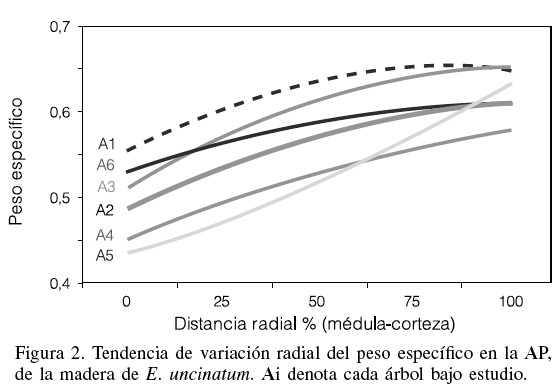
La amplitud de la variación radial promedio del peso específico para la madera de E. uncinatum (0-90%) alcanzó un valor de 22% (0,50-0,61). Sin embargo, se observó que ~73% de esta variación se genera en el árbol por debajo del 50% de la dirección radial. Estos resultados preliminares indican una zona con elevados niveles de variabilidad (adyacente a la médula) y una segunda zona donde los valores del peso específico tienden a mantenerse más o menos constantes, con una tendencia a la estabilización, tal y como se observa en la Figura 2, aspecto que está relacionado con la formación de madera adulta en el árbol (Panshin y De Zeeuw, 1980).
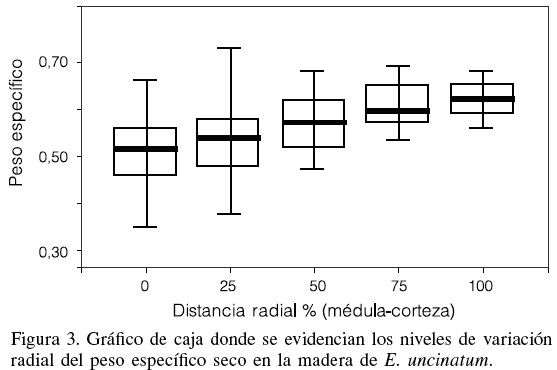
Los diagramas de cajas de la Figura 3 ofrecen información sobre el comportamiento de la variable peso específico en los cinco puntos de evaluación. Se evidencia una mayor variación o dispersión de los datos en los puntos más cercanos a la médula (cajas más altas y bigotes más largos). Sin embargo, a partir del 50% de la longitud de los radios en la dirección médula corteza, la dispersión de los datos disminuyó acercándose al valor medio, observándose además un mayor nivel de homogeneidad del parámetro evaluado entre los tres últimos puntos de evaluación.
La tendencia radial de variación del peso específico en la madera de E. uncinatum, es similar al reportado en la literatura especializada para especies latifoliadas. Por ejemplo, Arroyo (1983) afirmó que el modelo de variación más común del peso específico en la madera de las especies latifoliadas está caracterizado por un aumento en el sentido médula/corteza. Wiemann y Williamson (1988) determinaron incrementos del peso específico de la madera en dirección médula/corteza, asociado a estrategias de crecimiento en especies tropicales. Oliveira et al. (2005) reportaron un aumento en el sentido médula/corteza de la densidad de la madera para cinco especies de eucaliptos de Brasil.
Butterfield et al. (1993) en las especies Hyeronima alchorneoides y Vochysia guatemalensis observaron el mismo patrón de variación radial, con incremento del peso específico médula-corteza. Similares resultados reportaron Woodcock et al. (2000) en especies latifoliadas procedentes del Amazonas peruana. Woodcok y Shier (2002) y Parolin (2002) observaron un patrón de variación similar en especies latifoliadas de la zona templada y tropical, respectivamente, el cual fue atribuido a aquellas especies pioneras que mostraron un crecimiento apical acelerado en las etapas iniciales de desarrollo, y posteriormente incrementaron el valor del peso específico de la madera, ello con el propósito de garantizar soporte mecánico al crecimiento tanto del fuste como a la copa del árbol. Observaciones similares reportaron Sotelo et al. (2007) en la especie Calycophyllum spruceanum, procedente del Amazonas peruana.
El incremento del peso específico con la distancia radial puede explicarse desde el punto de vista fisiológico en relación al crecimiento del árbol; es decir, en los primeros años (zona juvenil) la cantidad de materiales fotosintetizados se utilizan en la creación de nuevas células (multiplicación anticlinal y periclinal), restando poco material para el engrosamiento de la pared celular. En cambio, en la etapa de madurez existe mayor cantidad de materia fotosintetizada destinada a la síntesis de la pared celular, lo que se traduce en un incremento del peso específico de la madera y de otras propiedades (Ismail et al., 1995; Moglia y López, 2001).
La variación radial observada en E. uncinatum puede estar relacionada con la edad fisiológica de las células que definen las características morfológicas del tejido xilemático. La porción del tronco alrededor de la médula presenta cambios progresivos en el aspecto celular y propiedades de la madera, típicos de la madera juvenil, y está caracterizada por la presencia de células cortas, delgadas, pequeñas con ángulos microfibrilares pequeños, grano espiralado y menores propiedades de resistencia (Panshin y De Zeeuw, 1980; Tsoumis, 1991; Amaraseka y Denne, 2002; Koubaa et al., 2005). Las células iniciales fusiformes aumentan en tamaño con el incremento de la edad del árbol, alcanzando un valor máximo, para luego tornarse relativamente estables. A partir de este punto se considera que estructuralmente la madera ha llegado a su etapa de madurez (Urbinati et al., 2003).
Conclusiones
La tendencia de variación del peso específico en la madera de E. uncinatum, no mostró diferencias significativas entre individuos; sin embargo, ésta fue evidente dentro de los árboles bajo estudio. La variación axial del peso específico mostró una disminución en la medida que se incrementó la altura de muestreo, observándose además que esta variación es menos consistente que la radial, apenas un 13% de variación. El patrón de variación radial del peso específico determinado para la especie mostró un incremento lineal desde la médula hasta la corteza, evidenciándose diferencias significativas por debajo del 50% de la dirección del radio, y luego se observó una normalización del parámetro evaluado.
AGRADECIMIENTOS
Los autores agradecen al FONACIT por el financiamiento otorgado a través del fortalecimiento del Centro Biotecnológico de Guayana (CEBIOTEG) de la Universidad Nacional Experimental de Guayana (UNEG) número Pem2001001639, y a la empresa CODEFORSA por el apoyo logístico y técnico en la búsqueda de las muestras de madera.
REFERENCIAS
1. Amareseka H, Denne M (2002) Effects of crown size on wood characteristics of Corsian pine in relation to definitions of juvenile wood, crow formed wood and core wood. Forestry 75: 51-61. [ Links ]
2. Arias S, Terrazas T (2001) Variación en la anatomía de la madera de Pachycereus pecten-aboriginum (Cactaceae). An. Inst. Biol. UNAM, Ser. Botánica 72: 157-169. [ Links ]
3. Arroyo J (1983) Propiedades Físico-mecánicas de la Madera. Facultad de Ciencias Forestales y Ambientales. Universidad de Los Andes. Mérida, Venezuela. 197 pp. [ Links ]
4. ASTM (1990) Standard Test Methods for Specific Gravity of Wood and Wood-Based Materials. ASTM D2395-83. Annual Book of ASTM Standards, Vol 04.09. Philadelphia, PA, EEUU. [ Links ]
5. Barajas J (1987) Wood specific gravity in species from two tropical forests in Mexico. IAWA Bull. 8: 143-148. [ Links ]
6. Barbosa R (1990) Analise do setor madeireiro do Estado de Roraima. Acta Amaz. 20: 1-209. [ Links ]
7. Bosman M (1996) Longitudinal variation in selected wood properties of naturally and plantation grown light Red meranti (Shorea leprosula y S. parvifolia, Dipterocarpaceae). IAWA J. 17: 5-14. [ Links ]
8. Butterfield R, Crook R, Adams R, Morris R (1993) Radial variation in wood specific gravity, fiber length and vessel area for two central American hardwoods: Hyeronima alchorneoides and Vochysia guatemalensis: Natural and plantation-grown trees. IAWA J. 14: 153-161. [ Links ]
9. CDF (2001) Plan de Ordenación y Manejo Forestal. Unidad de Manejo N-2. Reserva Forestal de Imataca. Tomo I, Cap. I-IV. Corporación de Desarrollo Forestal. Ministerio del Ambiente y de los Recursos Naturales. Upata, Venezuela. 200 pp. [ Links ]
10. Corothie H (1967) Estructura Anatómica de 47 Maderas de la Guayana Venezolana: Clave para su Identificación. Laboratorio Nacional de Productos Forestales. Universidad de Los Andes, Mérida, Venezuela. 125 pp. [ Links ]
11. Détienne P, Welle B (1998) Wood and Timber: Vochysiaceae. En Görts A, Jansen M (Eds.) Flora of the Guianas. Royal Botanical Garden. Kew, RU. Fasc. 21. pp. 66-70. [ Links ]
12. Durán J (1999) Manual de Experimentos para la Física de la Madera. Universidad de Los Andes. Mérida, Venezuela. 273 pp. [ Links ]
13. Espinoza J (2004) Gradiente de variación de la densidad de la madera en árboles de Gmelina aborea en Venezuela. IV Congreso Forestal Venezolano. Barinas, Venezuela. 56 pp. [ Links ]
14. Faría J, Rodríguez M, Silva R, León A (2005) Anuario Estadísticas Forestales Nº 8. Años 2002/2003/2004. Ministerio del Ambiente. Caracas, Venezuela. [ Links ]
15. Faría J, Rodríguez M, Pérez O, Ramírez S (2006) Anuario Estadísticas Forestales Nº 9. Año 2005. Ministerio del Ambiente. Caracas, Venezuela. [ Links ]
16. Fouquet D (1991) Utilization of wood and wood by-products in building construction in French Guiana. Boiset Forets des Tropiques. Nº 227. pp. 63-67. [ Links ]
17. García L, Guindeo A, Peraza C (2003) La Madera y su Anatomía. Mundi-Prensa/Fundación Salazar. Madrid, España. 327 pp. [ Links ]
18. Giménez A, López C (2000) Caracteres anatómicos que determinan la variabiliadad del leño de Schinopsis quebracho-colorado, Anacardiaceae. Rev. Inv. Agr. (INIA) 9: 219-235. [ Links ]
19. Gominho J, Figueira J, Rodríguez J, Pereira H (2001) Within-tree variation of hardwood extractives and wood density in the Eucalypt hybrid urograndis (Eucalyptus grandis & E. urophylla). Wood Fiber Sci. 33: 3-8. [ Links ]
20. IFLA (1992) Maderas Comerciales de Venezuela: Mureillo. Ficha técnica N° 5. Instituto Forestal Latioamericano. Mérida, Venezuela. pp. 27. [ Links ]
21. Igartúa D, Monteoliva S, Monterubbianesi M, Villegas M (2003) Basic density and fibre length at breast height of Eucalyptus globulus for parameter prediction of the whole tree. IAWA J. 24: 173-184. [ Links ]
22. Ismail J, Jusoh M, Sahari M (1995) Anatomical variation in planted Kelampayan Neolamarckia cadamo (Rubiaceae). IAWA J. 16: 277-288. [ Links ]
23. Jorge F, Quilho T, Pereira H (2000) Variability of fibre length in wood and bark in Eucalyptus globulus. IAWA J. 21: 41-48. [ Links ]
24. Justiniano M, Fredericksen J (1999) Ecología y Silvicultura de Especies menos Conocidas Cambará Hembra Erisma uncinatum Warm. Vochysiaceae. Proyecto de manejo forestal sostenible Bolfor. Santa Cruz, Bolivia. 29 pp. [ Links ]
25. Koubaa A, Isabel N, Zhang S, Beaulieu J, Bousquet J (2005) Transition from juvenile to mature wood in Black spruce (Picea mariana). Wood Fiber Sci. 37: 445-455. [ Links ]
26. Lei H, Milota M, Gartner B (1996) Between and within tree variation in the anatomy and specific gravity of wood in Oregon White oak (Quercus garryana Dougl.). IAWA J. 17: 445-461. [ Links ]
27. León W, Espinoza N (1999) Variabilidad de la madera de Cordia thaisiana (Boraginaceae) en sentido longitudinal. Rev. Forest. Venez. 43: 33-41. [ Links ]
28. Mainieri C, Pérez J (1989) Fichas de Características das Madeiras Brasileiras. Instituto de Pesquisas Tecnológicas. São Paulo, Brasil. 129-130 pp. [ Links ]
29. Milton S, Arnold J (2004) Probabilidad y Estadística, con Aplicaciones para Ingeniería y Ciencias Computarizadas. McGraw Hill. México. 804 pp. [ Links ]
30. Miranda I, Pereira H (2002) The variation of chemical composition and pulping yield with age and growth factors in young Eucalyptus globulus. Wood Fiber Sci. 34: 140-145. [ Links ]
31. Moglia J, López C (2001) Tendencias de variación radial del leño de Aspidosperma quebracho blanco. Inv. Agr. Sist. Rec. Forest. 10: 69-79. [ Links ]
32. Moret A, Ruiz P (1998) Determinación de ecuaciones de volumen para Mureillo (Erisma uncinatum) en la unidad C4 de la Reserva Forestal Imataca, Bolívar-Venezuela. Rev. Forest. Venez. 42:187-197. [ Links ]
33. Nogueira E, Nelson B, Fearnside P (2005) Wood density in dense forest in central Amazonia, Brazil. Forest Ecol. Manag. 208: 261-286. [ Links ]
34. Oliveira J, Hellmeister J, Filho M (2005) Variação do teor de umidade e da densidade básica na madeira de sete espécies de Eucalipto. Àrvore 29: 115-127. [ Links ]
35. Ortegano O, Silva R, Dugarte D, Valero J, Carrillo J (2001) Boletin Estadístico Forestal Nº 3. Año 2000. Ministerio del Ambiente y de los Recursos Naturales. Caracas, Venezuela. [ Links ]
36. Panshin A, De Zeeuw C (1980) Textbook of Wood Technology. 4a ed. McGraw Hill. Nueva York, EEUU. 722 pp. [ Links ]
37. Pardo A, Ruíz M (2005) Análisis de Datos con SPSS 13 Base. McGraw Hill. Madrid, España. 600 pp. [ Links ]
38. Parolin P (2002) Radial gradients in wood specific gravity in trees of central Amazonian Floodplains. IAWA J. 23: 449-457. [ Links ]
39. Pérez A (1973) Estructura anatómicas de 37 maderas de Guyana Venezolana y claves para su identificación. Acta Bot. Venez. 9: 9-109. [ Links ]
40. Quirk J (1980) Word anatomy of the Vochysiaceae. IAWA Bull. 1: 172-179. [ Links ]
41. Tsoumis G (1991) Science and Technology of Wood: Structure, Properties, Utilization. Van Nostrand Reinhold. Nueva York, EEUU. 494 pp. [ Links ]
42. Savva Y, Schweingruber F, Milyuntin L (2002) Genetic and environmental signal in tree ring from different provenance of Pinus sylvestris L. planted in the southern taiga, central Siberia. Trees 16: 124-131. [ Links ]
43. Silva R, León A, Sulbarán L, Catalán A, Carrillo J, Goudet M (2004) Anuario de Estadísticas Forestales Nº 7. Años 2000-2001. Ministerio del Ambiente y de los Recursos Naturales. Caracas, Venezuela. [ Links ]
44. Sotelo C, Hernández R, Beaulieu J (2007) Radial variation in wood density and correlations with growth of Calycophyllum spruceanum at an early age in the Peruvian Amazon. Wood Fiber Sci. 39: 377-387. [ Links ]
45. Urbinati C, Azevedo A, Monteiro E, Lisboa P (2003) Wood structural quantitative variation of the Terminalia ivorensis A. Chev. Combretaceae. Acta Bot. Bras. 17 (3): 421-437. [ Links ]
46. Vilela J (1969) Propiedades Físicas y Mecánicas de 137 Maderas de la Guyana Venezolana. Laboratorio Nacional de Productos Forestales. Mérida, Venezuela. 87 pp. [ Links ]
47. Wang T, Aitken S, Rozenberg P, Millie F (2000) Selection for improved growth and wood density in Lodgepole pine: Effects on radial patterns of wood variation. Wood Fiber Sci. 32: 391-403. [ Links ]
48. Wiemann M, Williamson G (1988) Extreme radial changes in wood specific gravity in some tropical pioneers. Wood Fiber Sci. 20: 344-349. [ Links ]
49. Wiemann M, Williamson G (1989) Wood specific gravity gradients in tropical dry and montane rain forest trees. Am. J. Bot. 76: 924-928. [ Links ]
50. Wilken J (1988) Variation in wood anatomy within species of Eucaliptus. IAWA Bull. 9: 13-23. [ Links ]
51. Woodcock D, Shier A (2002) Wood specific gravity and its radial variations: the many ways to make a tree. Trees 16: 437-443. [ Links ]
52. Woodcock D, Dos Santos G, Reynel C (2000) Wood characteristics of Amazon forest types. IAWA J. 21: 227-292. [ Links ]
53. Yanchuk A, Micko M (1990) Radial variation of wood density and fibre length in Trembling aspen. IAWA Bull. 11: 211-215. [ Links ]
54. Zhang S (1995) Effect of growth rate on wood specific gravity and selected mechanical properties in individual species from distinct wood categories. Wood Sci. Technol. 29: 451-465. [ Links ]
55. Zobel B, Van Buijtenen J (1989) Wood Variation: its Causes and Control. Springer. Berlín, Alemania. 200 pp. [ Links ]
