










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkArchivos Venezolanos de Farmacología y Terapéutica
versión impresa ISSN 0798-0264
AVFT v.24 n.1 Caracas 2005
Efectos de dietas enriquecidas con distintos aceites vegetales, en la potenciación post-reposo y en el cronotropismo cardiaco
C Alfonso-Pérez1, H Neptalí O2 y O Rodríguez S3.
1 Magíster Science. Sección de Investigaciones Cardio-Renales. Instituto de Medicina Experimental. Facultad de Medicina. Universidad Central de Venezuela.
2 Magíster Science. Escuela de Enfermería. Facultad de Medicina. Universidad Central de Venezuela.
3 PhD. Escuela de Medicina "José María Vargas". Facultad de Medicina. Universidad Central de Venezuela.
E-mail: alfonsoc@ucv.ve
RESUMEN
En este trabajo se evaluó la cinética de la potenciación en ventrículo derecho y el cronotropismo en aurículas aisladas. Utilizamos ratas alimentadas con Ratarina, Ratarina más 5% de aceite de oleína+soya o Ratarina más 5% de aceite de soya, por 6 semanas. Del corazón se aislaron ambas aurículas y tiras del ventrículo derecho. En las aurículas se determinó el efecto cronotrópico a diferentes concentraciones de epinefrina y en el ventrículo con estímulo se determinó la potenciación del primer latido con distintos intervalos de reposo. Se encontró un efecto cronotrópico disminuido en las aurículas del grupo con aceite de soya relacionado con un aumento del ácido linoleico y no con el aumento de las calorías, además, se encontró mayor cantidad de ácido oleico en los lípidos de las aurículas con respecto a los ventrículos. La mayor fuerza de contracción post-reposo y del estado estable se encontró en los animales que ingirieron mayor cantidad de lípidos totales.
Palabras Clave: Aceites vegetal, Tejido cardiaco, Cronotropismo, Potenciación post-reposo, Rata.
ABSTRACT
In this work the kinetic of potentiation post-rest was evaluated in right ventricle and the chronotropism in auricles isolated. We used rats fed with Ratarina, Ratarina more 5% of olein+soybean oil, or Ratarina more 5% of soybean oil, for 6 weeks. Of the heart both auricles and strips of the right ventricle were isolated. In the auricles it determined the chronotropic effect to different concentrations from epinephrine and in the ventricle with stimulus determined the potentiation of the first beat with different rest intervals. Was a diminished chronotropic effect in auricles of the group with soybean oil related to an increase of linoleic acid and not with the increase of the calories, in addition, was greater amount of oleic acid in lipids of the auricles with respect to the ventricles. The greater force of contraction post-rest and the stable state was in the animals that ingested greater amount of total lipids.
Key Words: Oils vegetal, Cardiac tissues, Chronotropic, Post-rest, Rat.
INTRODUCCIÓN
La eliminación de los lípidos de la dieta en ratas produce caída del pelo, descamaciones de la piel, edema, eritema en la cola y lesiones a nivel renal, ocasionando el síndrome por deficiencia de lípidos(1). Turpeinen demostró que el síndrome se curaba al agregar en la dieta ácido linoleico (C18:2,n-6) a partir del cual se forma el ácido araquidónico (C20:4,n-6), que participa en varias vías metabólicas(2). Brown y col, demostraron que los lípidos de la dieta regulan el número de receptores en las membranas de los hepatocitos y existía una correlación positiva entre la ingesta de ácidos grasos saturados y el colesterol plasmático, con mayor riesgo de formación de ateroma(3). En Venezuela, para el año 2002 el consumo de aceites vegetales fue de 17 kg /persona /año con 27% de calorías en forma de lípidos(4). Mediante técnicas electrofisiológicas de patch-clamp se ha determinado que el ácido graso C20:5,n-3 produce inhibición parcial de la corriente de calcio (ICa2+) en el miocito aislado de rata, los ácidos grasos poliinsaturados disminuyen la corriente de sodio (INa+), similar a la acción del C22:6,n-3; C20:4,n-6; C18:3,n-3 y 18:2,n-6; además, la inhibición de la corriente en los canal lentos de calcio I(CaL) son voltaje y tiempo dependiente, por lo cual, estos ácidos grasos son antiarrítmicos(5), los ácidos C18:0 y C18:1,n-9 no produjeron efecto sobre las corrientes I(CaL)(6). El ácido 18:1,n-9 de la fosfatidilcolina (FC) y fosfatidiletanolamina (FE) del sarcolema del ventrículo aumentó al primer día de tratamiento con epinefrina en ratas; al tercer día disminuyó la concentración del C18:1,n-9 y en la FC aumentaron los ácidos C18:2,n-6 y C20:4,n-6. Al séptimo día de tratamiento, el ácido C22:6,n-3 aumentó su contenido en ambos fosfolípidos y disminuyó del número total de receptores β, denominado "Down-regulation". Esto soportó la idea que alteraciones en la composición de los ácidos grasos de las membranas pueden influir en la respuesta de los segundos mensajeros en el músculo cardiaco(7). Leifert y col encontraron que los ácidos grasos n-3 bloquean las INa+ en miocitos ventricular de ratas adultas y se correlacionaba con el aumento de la fluidez de la membrana, mientras que, los ácidos grasos C18:2,n-6; C18:1,n-9 y C18:0 fueron menos efectivos en bloquear las INa+(8). En condiciones fisiológicas, el corazón utiliza como fuente de energía un 70% de los ácidos grasos y un 30% de glucosa; el transporte de estos ácidos grasos a través de las membranas del miocito se realiza mediante proteínas específicas como las proteínas que unen ácidos grasos (FAB) y las proteínas transferidoras (FAT)(9). La distribución celular y subcelular del FAT está presente tanto en la membrana plasmática, como en membrana del retículo endoplásmico, mientras que, el transportador de glucosa Glut-4 en el músculo cardiaco se encuentra predominantemente en la membrana plasmática y su redistribución está asociada a la respuesta a la insulina(10). Una variedad de estímulos extracelulares causan liberación de ácido C20:4,n-6 y diacilglicéridos. Pi y col, encontraron que en miocitos ventriculares aislados, los ácidos grasos y los diacilglicéridos interactúan directamente con la proteína cinasa C, participando en la liberación transitoria de calcio y incrementando paralelamente el efecto inotrópico, sugiriendo que estos compuestos intervienen en la regulación de la homeostasis del calcio(11,12). En el músculo cardiaco existen dos subtipos de receptores β adrenérgicos (β AR): β1 AR que está ubicado en la membrana plasmática y β2 AR está restringida a microdominios en la membrana plasmática (lípidos raft), en la cercanía al canal de calcio tipo L y en la cascada de reacciones del IP3 y no son modulados por receptores muscarínicos (m2) a acetilcolina (m2ACh), como ocurre con los receptores β1 AR. Sato y col, mostraron que cuando hay disfunción β1 AR y β2 AR la respuesta contráctil en los miocitos es mediada por una mayor expresión del intercambiador Na+/Ca2+(13,14). El objetivo de este trabajo fue evaluar los efectos de una dieta enriquecida con 5% de aceites vegetales comestibles, sobre la cinética de la potenciación post-reposo del ventrículo derecho, el cronotropismo en aurículas y los ácidos grasos de estos tejidos aislados de ratas.
MATERIALES Y MÉTODOS
Se utilizaron 18 ratas machos, de la raza Sprague-Dawley, con un peso inicial entre 84 ± 4 g, se dividieron en tres grupos: el primero ingirió ratarina, a base de cereales de fórmula cerrada(15), al segundo grupo la ratarina se enriqueció con 5% de aceite de oleína +soya y al tercero, la ratarina se enriqueció con 5% de aceite de soya y agua ad líbitum, durante 6 semanas. Después del período experimental, las ratas fueron anestesiadas vía intraperitoneal, con pentobarbital sódico (50 mg/Kg de peso). Mediante una esternotomía mediana se extrajo el corazón y se colocó en una solución Tyrode burbujeada con 95% de O2 más 5% CO2, pH 7,4 y se aislaron ambas aurículas y tiras de la pared libre del ventrículo derecho. Las preparaciones se fijaron en un baño de superfusión de órgano con 15 mL de solución Tyrode, la de aurícula contiene 5,0 mM de glucosa y 1,7 mM de CaCl2, mientras que, la solución tyrode del ventrículo contenía 10,0 mM de glucosa y 2,7 mM de CaCl2 a 35°C(16).
Determinación del cronotropismo
Ambas aurículas se dejan estabilizar en el baño de superfusión por 30 min, se registró la frecuencia espontánea del pre-tratamiento sin epinefrina (control), luego se determinó la frecuencia espontánea a concentraciones crecientes de epinefrina de 1x10-7 M hasta 3x10-4 M, y evaluamos la respuesta cronotrópica después de 5 min. Se calculó la diferencia de la frecuencia (lat/min) con respecto al pre-tratamiento, luego se determinó el porcentaje relativo a la respuesta máxima de epinefrina del grupo ratarina.
Determinación de la cinética de la potenciación post-reposo
Las preparaciones de tiras de ventrículo derecho fueron estimuladas bipolarmente(17). Las preparaciones se dejan estabilizar por 30 min. La fuerza de contracción post-reposo se obtuvo luego de detener la estimulación por períodos de 4, 8, 16, 32 ó 64 segundos.
Análisis del porcentaje de ácidos grasos de los lípidos totales
Los lípidos se extrajeron por el método de Folch y col(18). Se transesterifican a ésteres metílicos de ácidos grasos(19), y el porcentaje fue evaluado por cromatografía en fase gas-líquida en un equipo Hewlett Packard, modelo 6890®, con detector de llama, columna capilar de 30 m, 0,25 mm de diámetro y 0,25 µm de espesor de la fase estacionaria INOWAX®, a 200°C en condiciones isotérmicas, con una fase móvil de N2 a 1 mL/min. Se utilizaron patrones Sigma® de los ésteres metílicos de los ácidos grasos.
RESULTADOS Y DISCUSIÓN
El aceite de oleína de palma africana más soya incrementa el contenido de ácido C16:0 y C18:1,n-9 en la dieta, mientras que, el aceite de soya puro aumenta el contenido de ácidos grasos poliinsaturados y aportó un 51,9% de ácido C18:2,n-6, tabla 1, que se incorpora en mayor porcentaje en la posición sn2 de los fosfoacilglicéridos de las membranas, de donde puede ser liberado por acción de la fosfolipasa A2 no dependiente de calcio(20). No se presentaron diferencias en el peso corporal entre los grupos, aunque las dietas aportan diferentes contenidos de calorías en forma de lípidos. La ratarina con 2% de lípidos totales aportó entre 3,5 y 4,5 Kcal de lípidos/día y las dietas con 7% aportaron entre 12,6 y 15,8 Kcal de lípidos/día, esta cantidad no produce un aumento significativo del tejido de reserva(21). No obstante, los triacilglicéridos se pueden presentar como pequeñas gotas de lípidos en el músculo cardiaco en condiciones fisiológicas por el aumento de los lípidos de la dieta(22,23).
Porcentaje de los ácidos grasos en los lípidos totales de las dietas. Media ± E.E.
|
| Ratarina más 5% de aceite | ||
|
| Ratarina | Oleína + Soya | Soya |
| C14:0 C16:0 C16:1,n-9 C18:0 C18:1,n-9ª C18:2,n-6b C18:3,n-3 C20:5,n-3 C22:6,n-3 Lípidos (%) | 2,6 ± 0,4 19,5 ± 0,7 ª 3,1 ± 0,7 5,2 ± 0,7 23,9 ± 1,2 ª 37,2 ± 1,2 b 2,5 ± 0,7 2,8 ± 0,9 2,8 ± 0,9 2 7 | 0,5 ± 0,4 22,1 ± 0,4 ª 0,9 ± 0,4 3,4 ± 1,5 38,0 ± 3,0 a,b 33,6 ± 3,8 ª 2,5 ± 0,8 0,8 ± 0,5 0,8 ± 0,7 7 | 0,4 ± 0,1 13,5 ± 0,4 ª 0,9 ± 0,4 3,9 ± 0,9 24,5 ± 1,0 b 52,7 ± 0,6 a,b 2,5 ± 0,9 0,8 ± 0,8 0,8 ± 0,7 |
Letras iguales indican diferencias significativas entre los % del mismo ácido graso de los grupos, p <0,05. n=2
Las aurículas de todos los grupos mostraron un efecto cronotrópico positivo, a diferentes concentraciones de epinefrina (Gráfico 1), pero en el grupo oleína + soya encontramos un efecto cronotrópico mayor (aumento de la frecuencia) a concentraciones de epinefrina entre 3x10-6 M y 3x10-4 M, y en el grupo soya un efecto cronotrópico menor. No hubo diferencias significativas en el porcentaje de la frecuencia a las concentraciones de epinefrina £ 3x10-6M entre las aurículas de los tres grupos. Pero a la concentración de epinefrina 1x 10-5 M hasta 3x10-4 M se encontró mayor porcentaje de la frecuencia (p<0,05) respecto a la respuesta máxima en las aurículas del grupo oleína + soya en comparación al grupo soya (Gráfico 1). Las aurículas del grupo ratarina, no presentaron diferencias significativas de la frecuencia con respecto al grupo soya; aunque la dieta con aceite de soya posee mayor cantidad de lípidos totales (7%) que la dieta ratarina (2%), por lo tanto, la frecuencia no se altera por la cantidad de lípidos. Las aurículas del grupo oleína + soya presentaron cronotropismo mayor, a concentraciones de epinefrina mayor a 3x10-6 M, mientras que, las aurículas del grupo soya, mostraron un cronotropismo menor a partir de esa concentración. Estudios epidemiológicos han identificado que altas frecuencias cardiacas es un factor de riesgo de muerte. El efecto de las catecolaminas en organismos intactos parece ser indirecto y asociado al tejido adiposo, ya que, las catecolaminas a través de la lipólisis, alteran la composición de los lípidos de las membranas(24). La estimulación beta adrenérgica es un importante modulador fisiológico de la liberación de calcio, del aumento de la frecuencia cardiaca y de la fuerza de contracción; estos efectos están asociados a la activación de la proteína adenilciclasa-Gsa encargada de generar AMPc, PKA y de la activación de canales de calcio o corrientes If(12). La respuesta encontrada en este trabajo parece estar asociada al perfil de los ácidos grasos presentes en los lípidos totales de las membranas de las aurículas y no a la cantidad de lípidos totales en las dieta. En experimentos con conejos sometidos a dietas suplementadas con 13% de aceite de maíz o aceite crudo de palma africana, se encontró que los ácidos grasos de los lípidos totales en el tejido adiposo es un reflejo de los ácidos grasos ingeridos(20). En las aurículas aisladas de los diferentes grupos de dieta, la incorporación del ácido C18.1,n-9 fue similar en las aurículas para los tres tipos de dieta; mientras que, el ácido C18:2,n-6 fue mayor en las aurículas del grupo que ingirió la dieta con aceite de soya (Tabla 2). Las aurículas del grupo ratarina presentó menor porcentaje de ácidos grasos C18:2,n-6 y su disminución fue significativa (p<0,05) con respecto al porcentaje encontrado en las aurículas del grupo soya. Además, se encontró un aumento significativo (p<0,05) de los ácidos grasos poliinsaturados C20:4,n-6 y C22:6,n-3 con respecto al grupo soya. Las aurículas del grupo oleína + soya presentaron un aumento significativo (p<0,05) del C18:0 y una disminución significativa (p<0,05) del C18:2,n-6 con relación a las aurículas del grupo soya. Además, la relación de ácidos grasos Saturados/Poliinsaturados (S/P) presentes en el grupo oleína + soya fue 1,06. Las aurículas del grupo soya presentaron disminución (p<0,05) del porcentaje del C18:0, C20:4,n-6 y C22:6,n-3 con aumento (p<0,05) del C18:2,n-6. El ácido graso C18:2,n-6 se ha asociado a disminución de la frecuencia cardiaca en ratas recién nacidas(25). Grynberg y col, encontraron que en cultivos de células cardiacas aisladas de rata, tanto en presencia de ácido araquidónico o ácidos grasos de la familia n-3, el isoproterenol (1x10-6M) produjo un efecto cronotrópico positivo mayor en presencia de la familia n-3, lo cual no se correlaciona con nuestros experimentos(26). Corazones perfundidos con C22:6,n-3 aumentan la actividad del PKA que fosforila al fosfalambano(27). Existen receptores β adrenérgicos (β AR) en distintos dominios de membrana en los miocitos cardiacos. Los β2 AR están muy cercanos a los canales de calcio y se cree que sus señales son localizadas, activando canales de calcio. Los ácidos grasos de las dietas podrían modular la actividad de los receptores β2 AR en esos nanoambientes lipídicos de las membranas y por tanto, cambiar los efectos cronotrópicos, inotrópicos y lusitrópicos en el músculo cardiaco(28).
Porcentaje de la frecuencia en aurículas aisladas de ratas
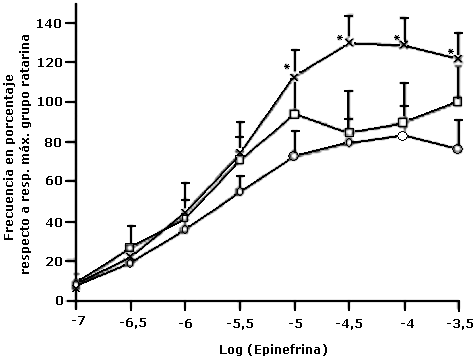
Porcentaje de la frecuencia en aurículas aisladas de ratas, sometidas a ratarina (), ratarina con aceite de oleína-soya ( x ) ó aceite de soya ( o ). El porcentaje es relativo a la respuesta con la dosis máxima de epinefrina del grupo ratarina. Media ± EE. N=6 para cada grupo.
(*) Diferencia significativa (p<0,05) entre los grupos de oleína+soya y soya.
Porcentaje de los ácidos grasos en las auriculas aisladas de rata, en los tres grupos de dieta. Media ± EE.
|
|
| Ratarina más 5% de aceite | |
| Ácidos Grasos | Ratarina (n=5) | Oleína + Soya (n=5) | Soya (n=5) |
| C12:0 C14:0 C15:0 C16:0 C16:1,n-9 C17:0 C18:0 C18:1,n-9 C18:2,n-6 C18:3,n-3 C18:3,n-6 C20:3,n-6 C20:4,n-6 C20:3,n-3 C20:5,n-3 C22:6,n-3 | 0,3± 0,1 1,9 ± 0,2 0,0 ± 0,0 25,5 ± 2,6 8,1 ± 3,3 0,0 ± 0,0 11,5 ± 2,6 22,5 ± 2,8 17,8 ± 4,9ª 0,6 ± 0,1 0,0 ± 0,0 0,5 ± 0,1 7,9 ± 1,1ª 0,0 ± 0,0 0,7 ± 0,3 2,7 ± 0,3ª | 0,4 ± 0,1 1,3 ± 0,1 0,0 ± 0,0 21,7 ± 1,0 2,0 ± 0,5 0,0 ± 0,0 14,6 ± 1,0ª 24,2 ± 1,9 23,2 ± 0,3b 0,6 ± 0,2 0,0 ± 0,0 0,5 ± 0,2 7,6 ± 1,8 0,0 ± 0,0 1,3 ± 0,8 2,6 ± 0,3 | 0,2 ± 0,0 1,3 ± 0,2 0,0 ± 0,0 19,6 ± 1,0 1,9 ± 0,2 0,0 ± 0,0 8,9 ± 1,7ª 24,4 ± 1,1 34,9 ± 0,5a,b 1,3 ± 0,2 0,0 ± 0,0 0,3 ± 0,1 5,0 ± 1,0ª 0,0 ± 0,0 0,5 ± 0,3 1,7 ± 0,3ª |
Letras iguales indican diferencias significativas entre los % del mismo ácido graso de los grupos, p<0,05.
En los ventrículos derechos la potenciación post-reposo está relacionada con la recaptación de calcio por la ATPasa Ca2+ (SERCA) del retículo sarcoplásmico (RS) durante el intervalo de reposo y su liberación por los receptores de rianodina (RYRs), durante la despolarización de la membrana. Con respecto a los resultados de la fuerza de contracción post-reposo (potenciación post-reposo del primer latido) en las tiras de ventrículo derecho, se encontró que la máxima potenciación, después de los distintos intervalos de reposo 4, 8, 16, 32, 64 segundos, se alcanzó a 32 segundos de reposo en todos los grupos estudiados (Gráfico 2), que está relacionado con la mayor cantidad del calcio recirculante y depende del receptor de RYR, SERCA y del RS, lo que indica que este fenómeno es independiente del contenido de lípidos totales en la dieta y el perfil de ácidos grasos en los ventrículos en este protocolo experimental, estos resultados son compatibles con otros publicados, donde se encontró que la máxima potencia post-reposo en los ventrículos fue 32 segundos, el cual disminuyó con 1,5x10-8 M de rianodina, cambiando la cinética de potenciación post-reposo del ventrículo(17).
Estudio cinético de la potenciación post-reposo del primer latido, en tiras de ventrículo derecho aislados de ratas
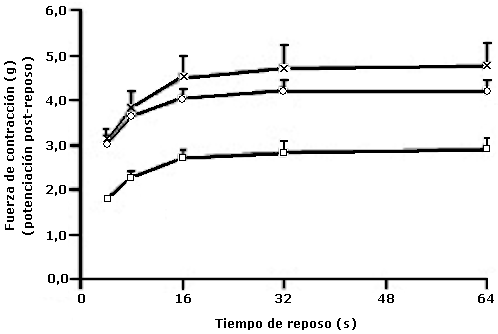
Media ± EE. El grupo ratarina (n=5) presentó diferencia significativa (p<0,05) de potenciación post-reposo, en todos los intervalos de reposo, respecto a los grupos con dieta suplementada con 5 % de aceite oleína+soya (n=5) ó con aceite de soya (n=4).
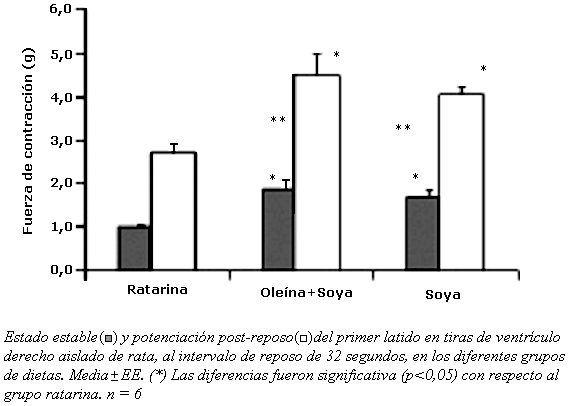
Las tiras de ventrículos del grupo oleína + soya presentaron mayor potenciación post-reposo y mayor fuerza de contracción de estado estable (Gráfico 3). Los ventrículos del grupo ratarina, presentaron menor potenciación post-reposo en todos los intervalos de reposo y menor fuerza de contracción en el estado estable. Se encontró un aumento significativo (p< 0,05) de potenciación post-reposo en los ventrículos del grupos oleína + soya o soya, con relación al grupo ratarina. Esta respuesta se podría explicar, porque la mayor cantidad de lípidos ingeridos puede estar relacionada con mayor beta-oxidación de ácidos grasos y síntesis del ATP requerido para la fuerza de contracción. Los ventrículos del grupo soya presentaron menor porcentaje de ácido C18:1,n-9 (p<0,05). No se encontró diferencias significativas en el porcentaje de los otros ácidos grasos (Tabla 3). El mayor porcentaje de ácidos grasos saturados en los ventrículos se observó en el grupo oleína + soya (42%). Los ventrículos del grupo soya presentaron el mayor porcentaje de ácidos grasos poliinsaturados (56,6%) y menor relación S/P (0,68), sin embargo, el perfil de ácidos grasos de los lípidos totales en los ventrículos no es un reflejo directo del porcentaje de ácidos grasos de la dieta. Pi y col, encontraron que en miocitos ventriculares aislados, el ácido C20:4,n-6, diacilglicéridos y PKC aumentan la liberación transitoria de calcio, sugiriendo que participan en la regulación de la hemostasia del calcio(12). El porcentaje de los ácidos grasos en las aurículas y ventrículos fueron distintos, al parecer cada tejido tiene la capacidad de regular el porcentaje de los ácidos grasos, aunque están influenciados por la dieta.
Porcentaje de los ácidos grasos en tiras de ventrículo aislado de rata, en los tres grupos de dieta. Media ± EE
|
| Ratarina más 5% de aceite | ||
| Ácidos Grasos | Ratarina (n=5) | Oleína + Soya (n=5) | Soya (n=5) |
| C12:0 C14:0 C15:0 C16:0 C16:1,n-9 C17:0 C18:0 C18:1,n-9a C18:2,n-6 C18:3,n-3 C18:3,n-6 C20:3,n-6 C20:4,n-6 C20:3,n-3 C20:5,n-3 C22:6,n-3 | 2,4 ±2,0 0,8 ±0,3 0,0 ±0,0 13,7 ±1,0 0,4 ± 0,1 0,0 ± 0,0 22,0 ± 1,1 6,1 ± 1,1 24,7 ± 1,0 0,4 ± 0,2 3,8 ± 3,8 0,7 ± 0,4 12,7 ± 3,3 0,0 ± 0,0 3,3 ± 2,9 9,0 ± 0,5 | 0,5 ± 0,1 0,4 ± 0,0 0,0 ± 0,0 16,2 ± 0,9 0,4 ± 0,2 0,0 ± 0,0 24,9 ± 1,1 7,8 ± 0,8 22,6 ± 3,0 0,3 ± 0,1 0,0 ± 0,0 0,4 ± 0,1 15,7 ± 0,8 0,0 ± 0,0 0,2 ± 0,0 10,6 ± 0,7 | 0,4 ± 0,2 2,1 ± 1,8 0,0 ± 0,0 13,6 ± 1,1 0,1 ± 0,1 0,0 ± 0,0 22,4 ± 2,0 4,8 ± 0,5a 29,7 ± 2,9 0,3 ± 0,1 0,0 ± 0,0 0,9 ± 0,6 15,2 ± 1,7 0,0 ± 0,0 0,6 ± 0,4 9,9 ± 1,1 |
Letras iguales indican diferencias significativas entre el % del mismo ácido graso de los grupos, p<0,05
REFERENCIAS Bibliográficas
1. Burr G O, Burr M M. A new deficiency disease produced by the rigid exclution of fat from the diet. J Biol Chem. 1929; 85: 354-367. [ Links ]
2. Turpeinnen O. Further studios on the unsaturated fatty acids essential in nutrition. J Nutrition. 1938; 15: 351-366. [ Links ]
3. Brown M S, Goldstein J L. How LDL receptors influence cholesterol and aterosclerosis. Sci Am. 1984; 251: 58-66. [ Links ]
4. INN, Instituto Nacional de Nutrición, Venezuela. 2002. Hojas de Balance de Alimentos. [ Links ]
5. McLennam D, Mano M, Rayner T, Head R. The cardiovascular protective role of docosahexaenoic acid. Eur J Pharmacol. 1996; 300: 83-89. [ Links ]
6. Xiao YF, Gomez AM, Morgan JP, Lederer WJ, Leaf A. Supresión of voltaje-gated L-type Ca2+ currets by polyunsaturated fatty acids in adult and neonatal rat ventricular myocytes. Proc Natl Acad Sci. USA. 1997; 94: 4182-4187. [ Links ]
7. Benediktsdottir VE, Curves J, Sigmundur G. Time of alterations in phospholipid fatty acids and number of β adrenoceptors in the rat Heart during adrenergic stimulation in vivo. J Mol Cell Cardiol. 1999; 31: 1105-1115. [ Links ]
8. Leifert WR, MC Murchie EJ, Saint DA. Inhibition of cardiac sodium currents in adult rat myocytes by n-3 polyunsaturated fatty acids. J Physiol. 1999; 520: 671-679. [ Links ]
9. Muller H, Deckers K, Eckel J. The fatty acid trnslocase (Fat)/CD36 and the glucose transporte GLUT4 are localizad in different cellular compartments in rat cardiac muscle. Biochemical Biophys Res Comun. 2002; 239: 665-669. [ Links ]
10. Roy R, Calvo JH, Hayes H, Rodellar C, Eggen A. Fine mapping of the bovine heart fatty acid-binding protein gene (FABP3 ) to BTA2q45 by fluorescence in situ hybridization and radiation hybrid mapping. Anim Genet. 2003; 34: 466-467. [ Links ]
11. Shinomura T, Asaoka Y, Oka M, Yoshida K, Nishizuka Y. Synergistic action of diacylglycerol and unsaturated fatty acid for protein kinase C activation: its possible implications. Proc Natl Acade Sci USA. 1991; 88: 5149-5153. [ Links ]
12. Pi Y, Jeffery WW. Diacyl glycerol y fatty acids synergistically increase cardiomyocyte contraction via activation of PKC. Am J Physiol Heart Circ Physiol. 2000; 279: H26-H34. [ Links ]
13. Sato M, Gong H, Terraccino CM, Harding SE. Loss of β- adrenoceptor response in myocytes overexpressing the Na+ /C2+ Exchanger. J Mol Cell Cardiol. 2004; 36: 45-48. [ Links ]
14. Steinberg SF. Beta (2)-Adrenergic receptor signaling complexes in cardiomyocyte caveolae/lipid rafts. J Mol Cell Cardiol. 2004; 37: 407-415. [ Links ]
15. Report of the American Institute of Nutrition Ad Hoc Comite on Standards for Nutritrional Studies. J Nutr. 1977; 1340-48. [ Links ]
16. Lamanna V, Alfonso-Pérez C, Rosas Luis, Maritza Padrón, Pérez de Lamanna Margarita. Efectos de las concentraciones crecientes de calcio y de los bloqueantes de canales lentos sobre el automatismo sinusal. Revista de la Facultad de Medicina. 1995; 18: 209-223. [ Links ]
17. Alfonso-Pérez C. Efecto de distintas maniobras inotrópicas sobre la potenciación post-reposo en aurículas izquierda y ventrículo derecho aislado de rata. Trabajo de ascenso. Biblioteca IME-UCV. 2002; 1-87. [ Links ]
18. Folch J, Lees M, Sloane GH. A simple method for isolation and purification of total lipids from tissues. J Biol Chem 1957; 266: 497-509. [ Links ]
19. Ortiz HN, Giacopini MI, Alfonso-Pérez C, Bosch VR. Efecto antioxidante del tomate (Lycopersicum esculentum Mill) en la lipoproteína de alta densidad (HDL) y en los microsomas hepáticos de la rata. Arch Ven Farm Terap. 2003; 22: 1-5. [ Links ]
20. Alfonso Pérez C. Efectos de las dietas enriquecidas con aceite de palma africana, maíz, ajonjolí y mezcla de aceites sobre la composición de los ácidos grasos esterificados a los fosfolípidos de las plaquetas, lípidos plasmáticos tejidos adiposo y la función plaquetaria. Bionotas. 1989; 1: 7-13. [ Links ]
21. Reeves P, Nielsen F y Fahey G. AIN-93 Purified diets for laboratory rodents: Final report of the American Institute of Nutrition Ad Hoc writing committee on the reformulation of the AIN-76A rodent diet. J Nutr. 1993; 123: 1939-51. [ Links ]
22. Oscai LB, Essig DA, Palmer WK. Lipase regulation of muscle triacilglyceride hydrolysis. J Appl Physiol. 1990; 69: 1571-1577. [ Links ]
23. Zhouy T, Grayburn P, Karim A, Shimabuk M, Higa M, Bartens D, Orci L, Unger RH. Lipotoxic heart disease in obese rats: implications for human obesity. Proc Natl Acad Sci USA. 2000; 15: 1784-1789. [ Links ]
24. De Lorgeril M, Salen P. Alpha-linolenic acid and coronary Heart disease. Nutr Metab Cardiovasc Disease. 2004; 14: 162-169. [ Links ]
25. Kang JX, Leaf A. Effects of long-chain polyunsaturated fatty acids on the contraction of neonatal rat cardiac myocytes. Proc Natl Acade Sci USA. 1994; 91: 9886-9890. [ Links ]
26. Grynberg A, Ziegler D, Rupp H. Sympathoadrenergic overactivity and lipid metabolism. Cardiovas Drugs Ther. 1996; 1: 223-230. [ Links ]
27. Negretti S N. Estudios de los efectos antiarrítmicos de los ácidos grasos w-3, en cardiomiocitos aislados de rata. Trabajo de ascenso, Biblioteca IME-UCV. 2003; 52-55. [ Links ]
28. Steinberg SF. β2–Adrenergic receptor signaling complexes in cardiomyocyte caveolae/lipid rafts. J Molec Cell Cardiol. 2004; 37: 407-415. [ Links ]
