










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkArchivos Venezolanos de Farmacología y Terapéutica
versión impresa ISSN 0798-0264
AVFT v.27 n.1 Caracas jun. 2008
Cinética de la potenciación post-reposo en tiras de ventrículo derecho aislado de rata, después de varias maniobras inotrópicas
Candelaria Alfonso-Pérez*, Vito Lamanna**, Holger N. Ortiz***.
* Magíster Science. Sección de Investigaciones Cardio-Renales, Instituto de Medicina Experimental, UCV. Caracas-Venezuela.
** PhD. Sección de Investigaciones Cardio-Renales, Instituto de Medicina Experimental, UCV.
*** Magíster Science. Escuela de Enfermería, Facultad de Medicina, UCV. Caracas-Venezuela.
No hay separatas disponibles. Ayuda parcial del CDCH-UCV, proyecto PI-09-33-4405-1999. Correo Email: alfonsoc@ucv.ve
Resumen
En este trabajo se realizaron maniobras inotrópicas positivas (disminución de [Na+]e, [K+]e y cambios de la frecuencia), y maniobras inotrópicas negativas (disminución de [Ca2+]e y adición de rianodina) en tiras del ventrículo derecho de ratas machos anestesiadas y estimuladas bipolarmente a 1 ó 3 Hz. La máxima potenciación se obtuvo a los 32 s, y disminuyó en las siguientes contracciones monotónicamente hasta alcanzar nuevamente el estado estable. La disminución de [Na+]e produjo un efecto inotrópico positivo ( p<0,05) hasta 16 s de reposo. Con reposo de 64 s y 3 Hz, la fuerza de contracción de los latidos fue menor, quizás por una mayor actividad del intercambiador Na+/Ca2+. La disminución de [Ca2+]e produjo un efecto inotrópico negativo hasta 16 s de reposo. La rianodina fue la única maniobra inotrópica que cambio la cinética de la potenciación post-reposo y del estado estable, lo que indica que la potenciación depende del Ca2+ liberado del retículo sarcoplásmico.
Palabras claves: tejido cardiaco, potenciación post-reposo, rianodina, rata
Abstract
In this work, positive inotropics maneuvers (diminution of [Na+]e, [ K+]e and changes of the frequency), and negative inotropics maneuvers (diminution of [Ca2+]e and adition of ryanodine) were made in right ventricle strip of male anaesthesia rats, stimulated bipolar to 1 or 3 Hz. The maxim potentiation was to the 32 s, with monotonic diminution until the stable state. The diminution of [ Na+]e produced a positive inotropic effect ( p<0,05) up to 16 s rest. With of 64 s rest and 3 Hz, the force of contraction of the beats was smaller, perhaps by a greater activity of Na+/Ca2+ interchanger. The diminution of [ Ca2+]e produced a negative inotropic effect up to 16 s rest. The ryanodine was only inotropic maneuver that change kinetic of potentiation the post-rest and of the stable state, which indicates that the potentiation depend of the Ca2+ released of the sarcoplasmic reticulum.
Key words: cardiac tissues, potentiation post-rest, ryanodine, rat
Recibido: 06/02/2008 Aceptado: 05/03/2008
Introducción
Se han desarrollado varios modelos para explicar el manejo celular del calcio en el músculo cardíaco, con la finalidad de describir la relación entre la potenciación de la fuerza de contracción y el intervalo de reposo en el músculo cardiaco de mamíferos1. El Ca2+ extracelular entra a la células cardiacas como consecuencia del potencial de acción, el cual abre los canales de Ca2+ dependientes de voltaje en la membrana plasmática y este Ca2+ al unirse a los receptores de rianodina (RYR) del retículo sarcoplásmico (RS), induce la liberación de Ca2+ intracelular2. Una vez que el Ca2+ es utilizado para el fenómeno de la contracción, la mayor proporción es nuevamente recaptado por el RS y en menor cantidad es eliminado hacia el medio extracelular por el intercambiador Na+/Ca2+, estos dos mecanismos varían en importancia en diferentes especies de mamíferos. Una pequeña cantidad del Ca2+ es expulsado de la célula por una bomba de calcio del sarcolema. El Ca2+ dentro del RS al ser transportado desde el sitio de recaptación cerca de las miofibrillas, hasta el sitio de liberación en un lugar cercano a los receptores de RYR requiere de un tiempo que varía entre las especie y cuyo mecanismo es desconocido3. Se ha tratado de explicar la potenciación post-reposo en base a un incremento del contenido de Ca2+ del RS, o a la fracción de Ca2+ liberado por el mismo, pero además, en algunas especies la potenciación de la contracción post-reposo está relacionada con el intervalo de reposo4. En este trabajo determinamos el tiempo de reposo en el cual ocurre la máxima potenciación y cuales maniobras inotrópicas cambian la cinética de la potenciación post-reposo en tiras de ventrículo derecho y en el manejo del calcio.
Materiales y métodos
Se utilizaron ratas machos Sprague Dawley, con peso entre 250 y 350 g. Después de anestesiadas con Nesdonal® intraperitoneal, se extrajo el corazón, se aisló una tira de la pared libre del ventrículo derecho y se fijó en un baño de superfusión con 15 mL de solución tyrode, pH 7,4 a 35 °C, burbujeada con 95 % de O2 y 5 % de CO2 con una tensión de basal de 1 g. Las preparaciones fueron estimuladas bipolarmente con pulsos de 3 ms, con una frecuencia basal de 1 Hz y una intensidad 1,5 veces el umbral. La fuerza de contracción expresada en gramos se registró en un polígrafo [5]. Se realizaron varias maniobras inotrópicas positivas: Disminución de [Na+]e de 149 a 75 mM , y de [K+]e de 3,9 a 1,0 mM, además, maniobras inotrópicas negativas: disminución de [Ca2+]e de 2,70 a 1,35 mM, presencia de 1,5x10-8 M de rianodina. También se estudiaron cambios de la frecuencia de 1 a 3 Hz. La fuerza contráctil se midió en el estado estable y luego de intervalos de reposo de 2, 4, 8, 16, 32, 64, 128 s. Una vez colocada las preparaciones en el baño de superfusión, se dejaron estabilizar por 30 min y se efectuó un registro control de la potenciación post-reposo con los distintos intervalos de reposo. Después se cambió la solución del baño para realizar la maniobra inotrópica, se dejó actuar por 10 min, y se hicieron los registros de la potenciación postreposo experimentales.
Resultados
En las tiras de ventrículo derecho, la máxima fuerza de contracción se alcanzó luego de un periodo de reposo de 32 s, con reposos mayores (hasta 128 s) permaneció constante. La potenciación de la fuerza de contracción a la frecuencia de 1Hz disminuyó monotónicamente hasta alcanzar el estado estable. Resultados similares se encontraron en la aurícula izquierda aislada de la rata, datos no mostrados.
Cuando se disminuyó la [Na+]e se encontró un aumento significativo (p< 0,05) en la fuerza de contracción post-reposo después de los intervalos de reposo 2, 4, 8 s. También aumentó la contracción en el estado estable (figura 1). No obstante, la disminución de la [K+]e no produjo cambios significativos en la potenciación, ni en el estado estable en relación al control, para los diferentes intervalo de reposo.
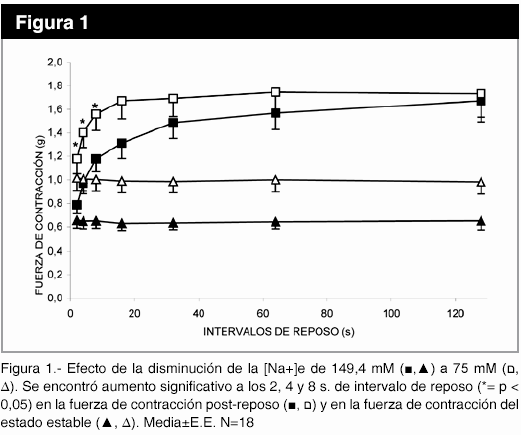
La disminución de la [Ca2+]e produjo una disminución significativa (p<0,05) de la fuerza de contracción post-reposo, entre los intervalos de reposo 2 a 16 s. También disminuyó la fuerza de contracción del estado estable (figura 2), mientras que, el aumento de la [Ca2+]e al doble del control no causó cambios significativos, resultados no mostrados.
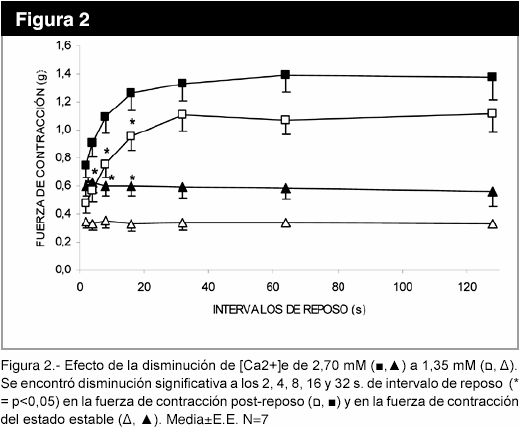
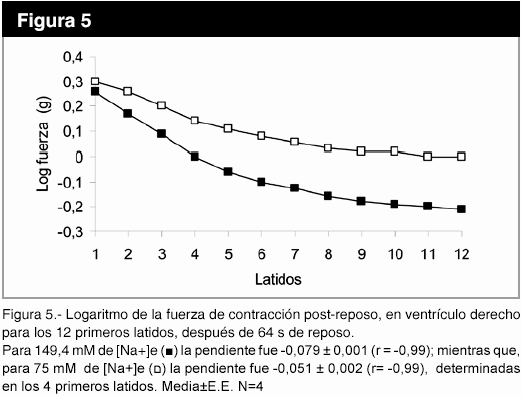
En la figura 3, se observa que el uso de la rianodina, un bloqueante específico de los canales de Ca2+ del RS dependientes de ligando, a la concentración de 1,5 x 10-8 M cambió la cinética de la potenciación post-reposo y disminuyó la fuerza de contracción del estado estable (p<0,05). El aumento de la frecuencia de 1 a 3 Hz no modificó la fuerza de contracción post-reposo (figura 4), pero la pendiente del gráfico logaritmo de la fuerza versus número de latido, con intervalo de reposo de 64 s, fue de -0,067±0,006 (r = -0,99) para 1 Hz; mientras que, para 3 Hz , la pendiente fue -0,09±0,02 (r= -0,94), determinados en los seis primeros latidos. La disminución de la [Na+]e produce una caída más lenta de la fuerza post-reposo latido a latido, con una fuerza de contracción mayor desde el primer latido, en relación a [Na+]e normal (figura 5).
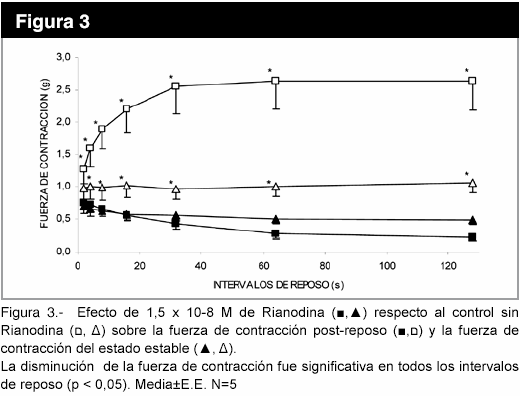
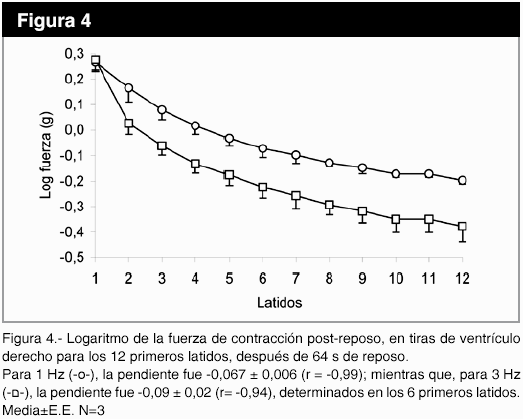
Discusión
En el ventrículo, la disminución de la [Na+]e provocó un efecto inotrópico positivo, posiblemente debido al incremento de la [Ca2+]i, originado por una disminución de la actividad del intercambiador Na+/Ca2+, o por que éste comienza a funcionar de forma inversa6. La [Na+]i en los miocitos es controlada por la actividad de la ATPasa Na+/K+, así como, la [Ca2+]i y [Na+]i regula al intercambiador Na+/Ca2+ 7. La disminución de la [Ca2+]e indujo el efecto inotrópico negativo esperado y por tanto, una disminución de la potenciación post-reposo y del estado estable, posiblemente a la menor cantidad de calcio en el retículo sarcoplásmico y menor [Ca2+]i recirculante8. La rianodina eliminó completamente la potenciación post-reposo,este compuesto, a la concentración utilizada, bloquea la liberación de Ca2+ del retículo sarcoplásmico; por tanto, el fenómeno de la potenciación post-reposo está estrechamente ligado al manejo del Ca2+ en el retículo sarcoplásmico9,10.
Con la frecuencia de 1 ó 3 Hz, la potenciación disminuyó en los latidos 8, 9 y 10 siguientes a la contracción post-reposo, esta disminución es debida posiblemente a la acción normal del intercambiador Na+/Ca2+, ya que, la pendiente de la depotenciación disminuye al bajar la [Na+]e, además, la pendiente aumentó cuando la frecuencia se cambia de 1 a 3 Hz, lo que aumenta rápidamente la concentración de [Ca2+]i.
En los ventrículos derechos de la rata Sprague Dawley, aparentemente se abre el mayor número de canales de calcio en el retículo sarcoplásmico a los 32s, tiempo en que se produce la máxima potenciación post-reposo. De las maniobras inotrópicas evaluadas, la rianodina fue la única que cambió la cinética de la potenciación post-reposo, lo que indica, que el fenómeno de potenciación en el ventrículo derecho está relacionado con el Ca2+ liberado del retículo sarcoplásmico.
Referencias
1. Schouten JV, Van Deen JK, De Tome P, Verveen AA. Force- intervalo relationship in Herat muscle of mammals. Biophys J. 1987; 51: 13-26. [ Links ]
2. Fabiato A. Time and calcium dependence of activation and inactivation of calcium-induced release of calcium from the sarcoplasmic reticulum of the skinned canine cardiac Purkinje cell. J Gen Physiol. 1985; 85: 247-289. [ Links ]
3. Allen DG, Jewell BR, Word EH. Studies of the contractility of mammalian myocardium at low rates of stimulation. J Physiol. 1976; 254:1-17. [ Links ]
4. Bers DM. Ca influx and sarcoplasmic reticulum Ca release in cardiac muscle activation during post-rest recovery. Am J Physiol 1985; 248:H376-381. [ Links ]
5. Alfonso C, Lamanna V, Rodríguez M, Pérez -González M. Efecto de distintas maniobras inotrópicas sobre la potenciación post-reposo en aurículas izquierda aislada de ratas. Revista Facultad de Medicina. 2002; 25: 59-62. [ Links ]
6. Janvier N.C., Boyett M.R. The role of Na-Ca exchage current in the cardiac action potential. Cardiovasc.Res. 1996; 32: 69-84. [ Links ]
7. Barry. W H., Bridge J H. Intracellular calcium homeostasis in cardiac Myocites. Circulation 1993; 57: 1806-1815. [ Links ]
8. Bassani, J W., Yuan W., Bers DM. Fractional S R Ca release is regulated by trigger Ca and R S Ca content in cardiomiocytes. Am J Physiol. 1995; 268: C1313 - C1347. [ Links ]
9. Bers DM. Ryanodine and the calcium content of cardiac RS assessed by caffeine and rapid cooling contractures. Am. J. Physiol 1987; 253:C408 - 415. [ Links ]
10. Zhang L, Kelley J, Schmeisser G, Kobayashi Y M, Jones L. Complex Formation between Junctin, Calsequestrin, and the Ryanodine Receptor. J.Biol.Chem 1997; 272: 23389-23397. [ Links ]
