










Servicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBioagro
versión impresa ISSN 1316-3361
Bioagro v.17 n.2 Barquisimeto abr. 2005
NOTA TÉCNICA
EFECTO DE LAS DIETAS DE ALIMENTACIÓN SOBRE LA BIOLOGÍA Y TABLA DE VIDA DE EUSEIUS CONCORDIS (CHANT) (ACARI: PHYTOSEIDAE)
Carmen Escalona1 y Carlos Vásquez1
1 Dpto de Ciencias Biológicas. Decanato de agronomía. Universidad Centrooccidental "Lisandro Alvarado". Aptdo. 400. Barquisimeto. Venezuela. email: cvasquez@ucla.edu.ve
RESUMEN
Se realizaron estudios de laboratorio para obtener información sobre la biología y tabla de vida del ácaro depredador Euseius concordis (Chant). El ácaro fue colectado en plantas de vid, cultivar Criolla Negra, en Tarabana, estado Lara, Venezuela. En el laboratorio, E. concordis fue alimentado con dos tipos de dietas consistentes de polen de yuquilla, Ruellia tuberosa L. o con individuos de diferente estado de desarrollo de Oligonychus punicae (Hirst). El tiempo promedio de desarrollo de E. concordis (huevo-adulto) no mostró diferencias (5,64 y 5,83 días) por efecto de la alimentación. Sin embargo, los períodos de preoviposición (3,3 y 1,7), oviposición (7,7 y 1,0), postoviposición (0,8 y 0,3 días) y longevidad (11,8 y 4,3) mostraron diferencias significativas entre sí. La fecundidad del ácaro alimentado con polen de yuquilla fue 0,7 huevos/hembra, mientras que solamente una hembra ovipositó durante 1 día, cuando fue alimentada con O. punicae. Los parámetros de la tabla de vida de E. concordis alimentado con polen mostraron valores para la tasa intrínseca de crecimiento (rm) = 0,1625 individuos/hembra/día, la tasa neta de reproducción (R0) = 4,0; el tiempo generacional (T) = 9,17 días y la tasa finita de crecimiento natural (l ) = 1,17 veces/hembra/día. La carencia de oviposición y sobrevivencia en la mayoría de las hembras alimentadas con O. punicae sugieren que el tetraníquido fue una presa inadecuada en la dieta del depredador.
Palabras clave adicionales:
Ácaros, oviposición, vid
Effect of diets on biology and life table of Euseius concordis (Chant) (Acari: Phytoseiidae)
ABSTRACT
Laboratory studies to obtain information about biology and life tables of the predatory mite Euseius concordis (Chant) were carried out. Mites were sampled in grapevines, cv. Criolla Negra, in Tarabana, Lara state, Venezuela. At the laboratory, E. concordis was fed on two sources consisting in minnieroot pollen, Ruellia tuberosa L. or with individuals at different development stages of Oligonychus punicae (Hirst) using leaf discs as rearing units. No differences on mean development time of E. concordis (5.31 and 5.83 days) were found when predatory mite fed on minnieroot pollen or O. punicae, respectively. However, preoviposition (3.3 and 1.7), oviposition (7.7 and 1.0), postoviposition (0.8 y 0.3 days) and longevity (11.8 and 4.3) showed significant differences. Fecundity of the predator fed on minnnieroot pollen was 0.7 eggs/female, whilst just one female oviposited during one day when fed on O. punicae. Life table parameters of E. concordis fed on pollen showed the following values: intrinsic rate of increse (rm) = 0.1625 individuals/female/day, net reproductive rate (R0) = 4.0, generation time (T) = 9.17 days and finite rate of increase (l ) = 1.17 times/female/day. Lack of oviposition and survival in most of the females fed on O. punicae suggests that tetranychid mite was an inappropriate prey.
Additional key words:
Mites, oviposition, grapevineRecibido: Diciembre 14, 2004 Aceptado:Agosto 31, 2005
INTRODUCCIÓN
Los ácaros de la familia Phytoseiidae son ampliamente usados como agentes de control biológico de diferentes especies de Tetranychidae en cultivos de importancia económica (Lofego et al., 2000). La baja tasa de consumo de alimento los favorece para establecerse y mantenerse con mayor eficiencia en los agroecosistemas (Silva, 2000).
McMurtry y Croft (1997) ubican a los ácaros fitoseídos en cuatro grupos funcionales basados en la preferencia de alimentación: tipo 1, depredadores especializados en el consumo de especies de Tetranychus; tipo 2, depredadores selectivos de ácaros Tetraníquidos; tipo 3, depredadores generalistas y tipo 4, depredadores palinívoros especializados-generalistas, representados por el género Euseius, los cuales se alimentan de polen y, ocasionalmente, de algunos estados evolutivos de Tetranychidae.
Existe poca información con relación al efecto de diferentes dietas alimenticias sobre los parámetros biológicos de las especies de Euseius. Broufas y Koveos (2000) encontraron que la tasa intrínseca de crecimiento de Euseius finlandicus (Oudemans) varió notoriamente cuando fue alimentada con polen de Malus silvestris Mill. o de Prunus avium L.
Con relación al potencial biológico de los ácaros fitoseídos, Reis et al. (2000) reportaron que Euseius alatus DeLeon mostró una mayor capacidad de búsqueda que Iphiseius zuluagai Denmark y Muma, aunque este último consumió mayor número de presas. Moraes y Lima (1986) encontraron que el tiempo de desarrollo de los estados inmaduros de Euseius concordis (Chant) fue similar cuando se alimentaron de Aculops lycopersici (Masse) o polen de Ricinus communis L.
En Venezuela, Aponte y McMurtry (1993) reportaron diversos frutales como hospederos de E. concordis entre los que se encuentra Vitis vinifera L. Sin embargo, hasta el presente no se ha investigado el potencial de esta especie para el control de ácaros plagas de la familia Tetranychidae.
Debido a que en el país, E. concordis es generalmente encontrado en asociación con el tetraníquido, Oligonychus punicae (Hirst), en diferentes especies de plantas, el propósito del presente estudio fue determinar la biología (ciclo de vida, preoviposición, oviposición, postoviposición, fecundidad, longevidad) y la tabla de vida de E. concordis alimentado con dos dietas basadas en granos de polen de yuquilla (Ruellia tubersosa) o con individuos de O. punicae.
MATERIALES Y MÉTODOS
Colecta, mantenimiento y determinación de la especie
Adultos del ácaro depredador E. concordis fueron colectados en hojas de vid, variedad Criolla Negra, en el Instituto de la Uva de la Universidad Centroccidental "Lisandro Alvarado" en Tarabana, estado Lara, Venezuela. La colecta consistió en desprender las hojas de la planta, las cuales fueron previamente revisadas con una lupa de mano para verificar la presencia de los ácaros depredadores. El material vegetal seleccionado fue entonces colocado dentro de bolsas plásticas recubiertas internamente con papel absorbente y fue llevado al laboratorio de Investigación de Zoología Agrícola de la Institución. Los ácaros fueron mantenidos en una sala de cría con un ambiente de 28 ± 2 °C, 60% ± 10 % HR y 12: 12 (D: N) horas de fotoperíodo.
Las hojas fueron observadas bajo el microscopio estereoscópico para seleccionar los ácaros fitoseídos, los cuales fueron montados en láminas microscópicas semi-permanentes. La clave taxonómica de Evans et al. (2001) fue utilizada para la identificación del género Euseius, mientras que la identificación de la especie E. concordis fue realizada por Gilberto de Moraes, ESALQ, Universidade de São Paulo, Brasil.
Ciclo de vida de E. concordis
El estudio del ciclo de vida de E. concordis se llevó a cabo en unidades de cría o arenas que consistieron en cápsulas de Petri (8,9 cm de diámetro por 1,8 cm de alto) con una almohadilla circular de poliuretano de 1 cm de espesor en su interior, previamente humedecida para mantener la turgencia de las hojas (Overmeer 1985). Sobre cada arena se colocó un disco de una hoja de vid con el envés hacia arriba rodeado por una banda de algodón humedecido para evitar el escape de los ácaros. Diez hembras y tres machos fueron colocadas en una arena, la cual fue replicada cinco veces. Transcurridas 24 horas, las unidades de cría fueron observadas al estereoscopio para determinar el número de huevos colocados/arena, hasta obtener un total de 50 huevos. Estos fueron entonces observados cada 12 horas para determinar el tiempo de incubación. Una vez emergidas las larvas, éstas fueron separadas en dos grupos. El grupo 1 consistió de 20 larvas, las cuales fueron alimentadas con polen de R. tuberosa, mientras que el grupo 2 consistió de 10 larvas, las fueron alimentadas con individuos en diferente fase de desarrollo de O. punicae. Cada 12 horas, se realizaron observaciones para determinar la duración de los estadios de larva, protoninfa y deutoninfa. Durante su desarrollo, los ácaros fueron transferidos a nuevas arenas en caso de que las hojas perdieran turgencia.
Los períodos de preoviposición, oviposición y postoviposición fueron medidos utilizando las hembras recién emergidas en cada grupo del ensayo del ciclo biológico. Una hembra y un macho fueron colocados en una arena para promover la cópula. El número de arenas fue replicado 10 veces. El período de preoviposición se determinó desde la emergencia de la hembra adulta hasta el momento en que colocó el primer huevo. Para el período de oviposición, se registró diariamente el número de huevos colocados por cada hembra durante su tiempo de vida, mientras que el período de postoviposición se midió desde que la hembra colocó su último huevo hasta su muerte.
Para el estudio de la fecundidad de E. concordis, diariamente el número de huevos puestos por la hembra, en cada una de las arenas, fue contado y anotado. Los datos fueron usados para calcular el promedio de huevos/hembra durante su vida, la tasa de oviposición expresada como huevos/hembra/día.
Para el estudio de la longevidad de E. concordis, el número de días que cada hembra permaneció viva en cada arena fue anotado y usado para calcular los rangos y el tiempo promedio de vida. La longevidad fue entonces graficada con relación al número de huevos puestos diariamente para mostrar la relación existente entre la fecundidad y la edad de la hembra de E. concordis.
Tabla de vida de E. concordis
La tabla de vida de E. concordis fue construida siguiendo la metodología descrita por Birch (1948). La tasa de sobrevivencia (lx) se expresó como el número de individuos vivos a un tiempo x, mientras que la tasa de fecundidad edad-especifica (Mx) se calculó tomando como base el número de descendencia hembras producida a partir de una hembra en un tiempo x. Los parámetros poblacionales estimados fueron 1) tasa neta de reproducción (Ro), 2) tiempo generacional (T), 3) tasa intrínseca de crecimiento (rm) y 4) tasa finita de crecimiento natural (l ), estimados a partir de los datos de fecundidad y sobrevivencia de las hembras. Estos parámetros han sido definidos por Rabinovich (1980) como sigue:
Tasa intrínseca de crecimiento (rm): la capacidad de multiplicación de una población en el lapso de una generación.
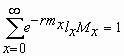
x: = edad de los individuos en días
lx = la proporción de individuos vivos a la edad "x"
Mx = número de descendencia hembra producida por cada hembra en el intervalo de edad "x"
Tiempo generacional (T): el tiempo promedio entre dos generaciones sucesivas.
![]()
Tasa neta de reproducción (Ro): Es el tiempo de reemplazo y refleja el número promedio de progenie hembra que es capaz de producir cada hembra de la población durante su vida.
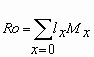
Tasa finita de crecimiento natural (l ): el número de individuos que se agrega a la población por individuo y por unidad de tiempo.
![]()
Análisis estadístico
Los ensayos sobre la biología de E. concordis fueron conducidos en un diseño completamente al azar con medidas repetidas en el tiempo. Los datos obtenidos fueron sometidos a análisis de varianza y pruebas de media según Tukey utilizando el paquete estadístico Statistix version 8.0.
RESULTADOS Y DISCUSIÓN
Duración del ciclo de vida de E. concordis en función de las dietas
El tiempo de desarrollo de E. concordis desde huevo hasta adulto no mostró diferencias significativas (P£ 0,05) por efecto de las dietas de alimentación aunque se detectó cierto efecto en las fases de larva y deutoninfa, sin tendencia definida (Cuadro 1). Cuando el ácaro depredador fue alimentado con granos de polen de yuquilla, el tiempo total de desarrollo fue estadísticamente igual (5,64 días) al de aquellos alimentados con una mezcla de diferentes estados de desarrollo de O. punicae (5,83 días). La duración del ciclo de vida del ácaro alimentado con ambos tipos de dietas fue ligeramente superior a aquellos registrados por Moraes y Lima (1983) para la misma especie alimentada con polen de R. comunis (5,2 días) o con A. lycopersici (4,95 días). Sin embargo, estos autores no observaron adultos cuando fueron alimentados con Tetranychus evansi Baker y Pritchard.
Tiempo de desarrollo en días de E. concordis alimentado con dos tipos de dieta
| Fases de desarrollo | Tipo de dieta | |
| R. tuberosa | O. punicae | |
| Huevo | 1,27a | 1,40a |
| Larva | 1,07b | 1,63a |
| Protoninfa | 1,30a | 1,80a |
| Deutoninfa | 2,00a | 1,00b |
| Total | 5,64a | 5,83a |
Valores de la misma fila seguidos de la misma letra no son diferentes según la prueba de Tukey (P£ 0,05)
Preoviposición:
Las hembras de E. concordis mostraron un período de preoviposición de 3,3 ± 0,55 días cuando fueron alimentadas con polen de yuquilla y de 1,7 ± 0,72 para aquellas alimentadas con una mezcla de diferentes estados de desarrollo de O. punicae (Cuadro 2). Broufas y Koveos (2000) reportaron que las hembras de E. finlandicus mostraron un período de preoviposición de 3 días cuando fueron alimentadas con varios tipos de polen. Sin embargo, cuando se alimentaron con tetraníquidos mostraron un período de preoviposición menor (1-1,5 días) (Moraes y Lima 1983; Badii y McMurtry, 1984).Oviposición:
La oviposición de E. concordis alimentadas con polen se distribuyó en dos picos de una semana cada uno. Durante el primero se obtuvo el mayor número de huevos que tendieron a disminuir en los días subsiguientes. A partir del día 8 se inició el segundo pico, el que mostró un patrón irregular con días de oviposición nula o casi nula de manera intercalada (Figura 1). Estos resultados coinciden con los estudios sobre fitoseídos donde se indica que la mayor oviposición se obtiene durante los primeros días de vida de la hembra adulta y que gradualmente disminuye a medida que la hembra madura (McMurtry et al., 1970). El tiempo promedio de oviposición de las hembras alimentadas con polen fue 7,7±5,23 días con rangos entre 2,8 y 14,8 días mientras que sólo una alimentada con O. punicae logró ovipositar durante un promedio de 0,3 ± 0,57 días (Cuadro 2). Aunque E. concordis ovipositó el 55 % de los huevos durante los primeros 6 días estuvo en capacidad de depositar un importante número de huevos después del día 6 hasta el día 13 (Figura 1). Broufas y Koveos (2000) encontraron que la oviposición de E. finlandicus duró 18 y 56 días cuando se alimentó con polen de pera y nogal, respectivamente.
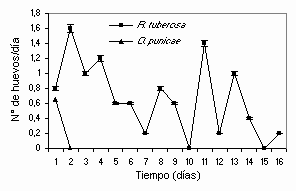
Figura 1. Oviposición diaria de las hembras de E. concordis alimentadas con granos de polen de R. tuberosa y especímenes de O. punicae
Postoviposición: La duración promedio del periodo de postoviposición de los ácaros alimentados con granos de polen fue 0,8 ± 0,35 días, mientras que cuando fue alimentado con O. punicae, el valor fue 0,9 días, sin embargo, este valor provino de una sola hembra que logró alcanzar el periodo de oviposición (Cuadro 2). Moraes y Lima (1983) encontraron que la postoviposición de E. concordis alimentado con polen de A. lycopersici fue de 0,7 días, lo cual esta de acuerdo con nuestros resultados. En contraste, Badii y McMurtry (1984) obtuvieron que cuando Phytoseiulus longipes fue alimentado con el ácaro T. pacificus, el período de postoviposición fue de 12,5 días.
Preoviposición, oviposición, postoviposición, fecundidad y longevidad en días de E. concordis alimentado con dos tipos de dieta
| Período (días) | Tipo de dieta | |
| R. tuberosa | O. punicae | |
| Preoviposición | 3,3 a | 1,7 b |
| Oviposición | 7,7 | 1,0 |
| Postoviposición | 0,8 a | 0,3 a |
| Longevidad | 10,85 a | 4,3 b |
| Fecundidad(huevos/hembra) | 0,7 | 0,33 |
Datos transformados por ![]()
Valores de la misma fila seguidos de la misma letra no son diferentes según la prueba de Tukey (P£ 0,05)
Fecundidad: La fecundidad promedio de E. concordis alimentadas con polen durante su vida fue 0,7 huevos/hembra/día, mientras que la tasa de oviposición fue 0,3 huevos/hembra/día con un máximo de 1,6 y 1,4 huevos puestos los días 2 y 11, respectivamente. Los valores obtenidos mostraron amplia variación, pues mientras un grupo de hembras del depredador (60%) ovipositó 0,96 ± 0,35, el otro grupo (40%) ovipositó 0,17 ± 0,15 huevos/hembra/día. Al contrario, cuando las hembras del ácaro depredador fueron alimentadas con O. punicae la fecundidad estuvo representada por una sola hembra, la cual ovipositó un máximo de 3 huevos inviables (
Cuadro 2). Investigaciones conducidas por Moraes y Lima (1983) reportaron valores para la fecundidad de E. concordis desde 2,1 cuando se alimentó con polen de tártago, mientras que Ferla y Moraes (2003) reportaron 0,9 huevos/día cuando se alimentó con polen de Typha angustifolia L. Adicionalmente, Ferla y Moraes (2003) demostraron que la oviposición diaria de E. concordis disminuyó hasta 10 veces cuando fue ofrecido Oligonychus gossypii (Zacher) como alimento. En general, se considera que las especies de Euseius tienen una tasa reproductiva media y que probablemente las especies de tetraníquidos no son su alimento más apropiado (James, 1989; Abou-setta y Childers, 1989).Longevidad:
La longevidad fue afectada por las dietas de alimentación (P£ 0,05). Aquellas alimentadas con polen duraron un tiempo promedio de 10,85±4,93 días, mientras que cuando se alimentaron con O. punicae el valor fue 4,3±2,33 (Cuadro 2). Broufas y Koveos (2000) reportaron valores de longevidad para E. finlandicus de 18,2 días cuando fue alimentado con polen de pera.Tabla de vida de E. concordis
La tabla de vida sólo pudo ser construida para los ácaros alimentados con polen de yuquilla, debido a que sólo una hembra de E. concordis alimentada con O. punicae logró ovipositar, haciéndolo durante un día. Esto permite concluir que esta última dieta fue inadecuada para la sobrevivencia y fecundidad del depredador. Estos resultados coinciden con la investigaciones conducidas por Moraes y Lima (1983) quienes indicaron que E. concordis no alcanzó su desarrollo cuando se alimentó con T. envansi Baker y Pritchard.
Con relación a los parámetros de vida de E. concordis alimentados con polen de yuquilla se obtuvo que la tasa intrínseca de crecimiento (rm) fue de 0,1625 individuos/hembra/día. La población se multiplicó 4 veces (Ro) en un tiempo generacional (T) de 9,17 días. La tasa finita de crecimiento natural (l ) fue 1,17 veces/hembra/día. Los valores obtenidos en este estudio son comparativos a los registrados por Broufas y Koveos (2000) para E. finlandicus, quienes reportaron un valor de Ro de 3,74, mientras que el valor de de rm fue 0,082 para hembras criadas con polen de pera. Basados en los valores de la tabla de vida de las especies de Euseius indican que estas especies presentan una baja tasa de reemplazo poblacional por lo que se podría inferir sobre su bajo potencial de mantenimiento bajo condiciones de campo.
La curva de sobrevivencia (lx) se mantuvo constante durante los primeros 5 días después del cual tendió a decrecer gradualmente, mientras que la tasa de fecundidad edad-específica (Mx) fluctuó entre 0,2 y 1,6 huevos/hembra/día durante los primeros ocho días de vida de las hembras de E concordis (
Figura 2).
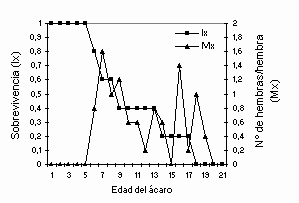
Figura 2. Sobrevivencia (lx) y fecundidad por edad (Mx) de E. concordis alimentado con polen de R. tuberosa
La curva de sobrevivencia de E. concordis mostró un patrón tipo I, en la cual la sobreviencia desde huevo hasta adulto fue cercana al 100% y la mayor mortalidad de las hembras ocurrió gradualmente. Este comportamiento también ha sido registrado para Typhlodromalus peregrinus (Muma) alimentado en diferentes dietas de especies de tetraníquidos y polen de varias especies (Fouly et al., 1995). Similarmente, Badii y McMurtry (1984) encontraron que las curvas lx y Mx de P. longipes también decrecieron rápidamente durante los primeros días. El bajo valor de los parámetros de la tabla de vida de E. concordis obtenidos en este estudio demuestran la baja calidad del alimento ofrecido al depredador lo que afectaría negativamente la dinámica poblacional de esta especie.
CONCLUSIONES
La duración del ciclo biológico de E. concordis no fue afectada por efecto de las dietas evaluadas, sin embargo, se encontró que los parámetros reproductivos fueron superiores en hembras alimentadas con polen de R. tuberosa comparados con las alimentadas con O. punicae. El bajo valor de la tasa intrínseca de crecimiento (rm) obtenido en hembras alimentadas con polen permite inferir que este tipo de alimento no aportó los requerimientos nutricionales del ácaro. Por otra parte, la dieta con tetraníquidos fue inadecuada para la supervivencia y fecundidad del depredador y por lo tanto E. concordis no parece ser un agente de control biológico eficiente para regular las poblaciones de O. punicae en los viñedos.
LITERATURA CITADA
1. Abou-Setta, M.M. y C.C. Childers. 1989. Biology of Euseius mesembrinus (Acari: Phytoseiidae): life tables and feeding behavior on tetranichid mites in citrus. Environ. Entomol. 18: 665-669. [ Links ]
2. Aponte, O. y J. McMurtry. 1993. Phytoseiid mites of Venezuela (Acari: Phytoseiidae). Int. J. Acarol. 19: 149-157. [ Links ] 3. Badii, M. y H. McMurtry. 1984. Life history of and life table parameters for Phytoseiulus longipes with comparative studies on P. persimilis and Typhlodromus occidentalis (Acari: Phytoseiidae). Acarologia 25: 111-123. [ Links ] 4. Birch, L. 1948. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol. 17: 15-26. [ Links ] 5. Broufas, G.D. y D.S. Koveos. 2000. Effect of different pollens on development, survivorship and reproduction of Euseius finlandicus (Acari: Phytoseiidae). Environ. Entomol. 29: 743-749. [ Links ] 6. Evans, D., J. McMurtry y G. Moraes, de. 2001. Key to the genera of Phytoseiidae (Adult females). Agricultural Acarology. 51st Annual Acarology Summer Program. The Ohio State University. [ Links ] 7. Ferla, N. y G. Moraes de. 2003. Oviposição dos ácaros predadores Agistemus floridanus Gonzalez, Euseius concordis (Chant) e Neoseiulus anonymus (Chant y Baker) (Acari) em resposta a diferentes tipos de alimentaçao. Rev. Bras. Zool. 20: 153-155. [ Links ] 8. Fouly, A. H., M. M. Abou-Setta y C. Childers. 1995. Effects of the biology and the life tables of Typhlodromus peregrinus (Acari: Phytoseiidae). Environ. Entomol. 24: 870-874. [ Links ] 9. James, D.G. 1989. Influence of diet on development, survival and oviposition in Asutralian phytoseiid, Amblyseius victoriensis (Acari: Phytoseiidae). Exp. Appl. Acarol. 6: 1-10. [ Links ] 10. Lofego, A., G.J. Moraes y J. McMurtry. 2000. Three new species of Phytoseiid mites (Acari: Phytoseiidae) from Brazil. An. Soc. Entomol. Brasil. 29: 461-553. [ Links ] 11. McMurtry, J.A. y B.A. Croft. 1997. Life-styles of phytoseiid mites and their roles in biological control. Annu. Rev. Entomol. 42: 291-321. [ Links ] 12. McMurtry, J.A., C.B. Huffaker y M. van de Vrie. 1970. Ecology of tetranychid enemies: their biological characters and the impact of spray practices. Hilgardia 40: 331-390. [ Links ] 13. Moraes, G.J. de, y H. Lima. 1983. Biology of Euseius concordis (Chant) (Acarina: Phytoseiidae) a predator of the tomato russet mite. Acarología. 24: 251-255. [ Links ] 14. Overmeer, W.P.J. 1985. Techniques: Rearing and Handling. In: W. Helle y M. W. Sabelis (eds.). Spider Mites. Their Biology, Natural Enemies and Control. Vol. 1B. Elsevier. pp. 161-170. [ Links ] 15. Rabinovich, J. 1980. Introducción a la Ecología de Poblaciones Animales. Compañía Editorial Continental, C. A. México. p. 313. [ Links ] 16. Reis, P., L. Chiavegato, E. Alves y E. Sousa. 2000. Ácaros da familia Phytoseiidae associados aos Citros no municipio de Lavras, Sul de Minas Gerais. An. Soc. Entomol. Brasil. 29: 95-104. [ Links ] 17. Silva, N. A. 2002. Caracterização morfológica e molecular de ácaros predadores do gênero Euseius (Acari, Phytoseidae). Tesis. Universidade de São Paulo, Piracicaba. Brasil. [ Links ]
