










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBioagro
versión impresa ISSN 1316-3361
Bioagro vol.29 no.3 Barquisimeto dic. 2017
Characterization of a wild strain of Saccharomyces cerevisiae for obtaining a biopreparation with probiotic profile
Johanna M. Arias1, Cecilia Lara2, Rodrigo Salgado3, Angélica M. Torregroza3 and Ana C. Torregroza4
1 Institución Educativa Marceliano Polo, Cereté, Colombia. e-mail: johannarias@hotmail.com
2 Universidad de Córdoba, Montería, Colombia
3 Corporación Universitaria del Caribe (CECAR), Facultad de Ciencias Básicas, Ingeniería y Arquitectura. Sincelejo, Colombia. e-mail: rodrigo.salgado@cecar.edu.co (autor de correspondencia); angelica.torregroza@cecar.edu.co
4 Corporación Universidad de la Costa (CUC), Facultad de Ingeniería, Programa de Ingeniería Agroinsdustrial. Barranquilla, Colombia. e-mail: atorregr4@cuc.edu.co
ABSTRACT
Generation of organic wastes is a worldwide environmental issue, although, if properly handled they may be a valuable source of animal nutrients. The objective was to determine the performance of a wild strain Saccharomyces cerevisiae in a biopreparation from ruminal content enriched with peels of fruits. Probiotic properties of the wild strain such as tolerance to bile salts, pH changes, changes in temperature, high concentrations of sodium chloride; additional to antagonism testing and gas production from glucose were verified in vitro. Microbial growth was evaluated in a medium prepared from clarified ruminal content 40 % v/v, enriched with peel wastes from papaya, pineapple and banana at 25, 50, and 75 % w/w concentrations. Results showed that wild strain had a higher growth in the medium obtained with 50 % w/v fruit wastes. Growth of the strain in this medium disclosed the highest biomass production at 40 hours (2.28x108 cfu·mL-1). The viability of the strain showed no important decrease during two months in the log scale of the inoculum. It is concluded that ruminal content enriched with fruit peels provides necessary nutrients for growth of the wild strain S. cerevisiae with probiotic characteristics and the biopreparation represents a supplement which may be useful not only at nutritional level but also to decrease environmental impact caused by these organic wastes.
Additional key words: Ananas comosus, Carica papaya, Musa sapientum, ruminal content
Caracterización de una cepa nativa de Saccharomyces cerevisiae para obtener un biopreparado con perfil probiótico
RESUMEN
La generación de desechos orgánicos es un problema medioambiental mundial, aunque, si se maneja adecuadamente, puede ser una valiosa fuente de nutrientes para animales. El objetivo fue determinar el rendimiento de una cepa silvestre de Saccharomyces cerevisiae en un biopreparado a partir de contenido ruminal enriquecido con cáscaras de frutas. Se verificaron in vitro las propiedades probióticas de la cepa como tolerancia a sales biliares, cambios de pH, cambios de temperatura y altas concentraciones de cloruro de sodio, así como de pruebas de antagonismo y producción de gas a partir de la glucosa. Se evaluó el crecimiento microbiano en el medio elaborado a partir del contenido ruminal al 40% v/v, enriquecido con desechos de cáscaras de papaya, piña y banano en concentraciones de 25, 50 y 75 %. El mayor crecimiento se produjo en el medio con 50 % de desechos de frutas y la cepa reveló la mayor producción de biomasa a las 40 horas (2,28x108 ufc·mL-1). Su viabilidad no presentó disminución importante en la escala logarítmica del inóculo durante dos meses. Se concluye que el contenido ruminal enriquecido con cáscaras de frutas al 50 %, proporciona los nutrientes necesarios para el crecimiento de S. cerevisiae con características probióticas, y el biopreparado representa un suplemento que no solo puede ser benéfico a nivel nutricional, sino que también disminuiría el impacto ambiental ocasionado por estos desechos orgánicos.
Palabras clave adicionales: Ananas comosus, Carica papaya, contenido ruminal, Musa sapientum
Recibido: Enero 24, 2017 Aceptado: Agosto 1, 2017
INTRODUCTioN
Nowadays generation of organic wastes is a worldwide environmental issue, especially those produced from meat industry, because they represent high waste volumes which are also accumulated in inappropriate spaces (Guerrero & Ramirez, 2004). Every year production of worldwide bovine meat increases, it is estimated that growth has been 2.3 million tons during last years, and production of waste from animal sacrifice has grown at an accelerated rate, as a consequence of the productive process of meat processing plants and slaughterhouses. Ruminal content is one of the contaminants with the highest environmental impact because it produces a high organic amount in effluents from meat processor that, due to their settle form, lead to septic tanks, municipal landfills and waste waters (FAO, 2006), thereby promoting contamination. However, ruminal content, instead of being considered contaminant, may be analyzed as a valuable source of animal nutrients because it contains raw protein and energetic materials (Rendueles & Díaz, 2014).
In other hand, wastes or agricultural by-products such as peels, shells, seeds, cores and ends or crowns produced by the consumption of fruits also negatively affect environment (Lopera et al., 2009). When disposed, microorganisms, parasites and enzymes present in food are the main sources of decomposition through vital phenomena, intervening in physical and chemical transformation processes of substances composing them (Pascual & Calderon, 2000; Mossel et al., 2003).
According to the National Agricultural Survey, the department of Córdoba is the main producer of papaya crops (Carica papaya) in Colombia with a total of 69.6 tons equivalent to 65.6 % of national production in the year 2014 (DANE, 2014); also, this department produced 284 tons of pineapple (Ananas comosus), representing 0,17% of total in the country. Data of banana crops (Musa sapientum) are not present in the survey, but this fruit is always available because Córdoba is close to the department of Antioquia, one of the highest production of this fruit in Colombia.
For all the above exposed, by the suitable use of said organic wastes (ruminal content and fruit peels) in the elaboration of a biopreparation, it is intended to give not only added value to the by-products of livestock and fruit growing activities, for having nutritionally interesting elements instead of being considered as contaminants, but also, when providing food for animals, they may increase quality and decrease production costs thereof.
Incorporation of probiotic crops as additives that promote productive response in animals has shown a renewed interest as a solution to the need of reducing the use of antibiotics in the animal food, being also beneficial for the final user (García, 2012). Existen millares de micro-organismos patógenos de animales por lo que uno de los medios más eficaces para combatirlos es la que puede realizar la propia flora intestinal (Drisko et al., 2003). The reason of this is the advantages offered by use thereof, by being from safe origin, generally stable, do not produce accumulative effects, are useful in controlling diseases without causing adverse effects, improve conversion of food, increase live weight and animal growth, as well as improved metabolism of food. It is emphasized that probiotics allow the intestinal microbiota to be kept under equilibrium and therefore, avoid settlement of gut pathogens (Valdovinos, 2013), thus preventing the use of antibiotics and increases of economic expenses (Valdovinos, 2013; Abreu, 2012). Therefore, the objective of this study was to determine conditions necessary to obtain a high concentration of biomass of the wild strain Saccharomyces cerevisiae with probiotic characteristics using a biopreparation from ruminal content as substrate, enriched with peels of three fruits found in the Department of Córdoba (papaya, pineapple and banana), in order to be evaluated in vivo in subsequent research.
MATERIALS Y METhODS
The research was carried out through the following three stages:
A) In vitro probiotic capacity test.
A suspension of a wild strain of S. cerevisiae was provided by Grubiodeq Lab (Biotechnology Group, University of Cordoba), to which testing was carried out to evaluate probiotic capacity thereof. Assays were carried out in triplicate. To carry out in vitro probiotic tests, yeast extract-peptone-dextrose (YPD) culture media were prepared (yeast extract 1 %, peptone 2 %, dextrose 1 %). Inoculums of the wild strain of S. cerevisiae used in each testing had known concentration (logarithmic range 107) for subsequent analysis.
Tolerance to bile salts: Salt concentrations evaluated were 0.05, 0.10, 0.15 and 0.30 % (w/v) of bile salt by adjusting pH 7.0±0.2. One mililiter of the wild strain was inoculated in each YPD media (9 mL contained in glass flasks), incubated at 30 ºC, and counts of viable cell carried out 24 hours later (Rubio, 2008).
Tolerance to pH: Tests were performed at pH 3, 4, 5.6, and 7 ± 0.2 (by adjusting the YPD culture medium with undiluted HCl as of the case), followed by sterilization process. Adding 1 mL of the suspension of the wild strain in glass flasks with 9 mL of YPD broth, incubating at 30 ºC and carrying out viable cell count at 24 hours (Rubio, 2008).
Tolerance to different temperatures: The growth of the wild strain was evaluated at 28 and 43 ± 1 °C. One milliliter of the suspension was added in 9 mL of YPD broth, inoculated during 24 hours and counts of viable cell was carried out (Rubio, 2008).
Tolerances to high NaCl concentrations. The wild strain suspension was inoculated in YPD broth in the presence of 2, 4, 7 and 10 % (w/v) NaCl, and incubated at 37 ºC for 24 hours. At this time, growth was determined by measuring optical density at 600 nm (Rodón et al., 2004).
Antagonism test: A massive bacterial population of Escherichia coli was grown in Petri dishes with agar Mueller Hinton. Then, discs impregnated with the suspension of the wild strain were placed on it. The dishes were refrigerated at 10 ± 2 ºC by half hour, then incubated at 37º C. After 48 hours growth inhibition zone was observed and measured (Leiva et al., 2004).
Gas production from glucose: For this test 0.3 mL of the wild strain suspension were inoculated in inverted vials with YPD media, containing 0.2 % (v/v) bromocresol purple aqueous solution (0.5 %). After inoculation, they were incubated for 48 hours at 37 ºC. Then, the presence or absence of gas burbles was registered (Rodón et al., 2004).
B) Formulating the biopreparation
Preparation of media was carried out by collecting samples of fruit shells (peels) in fruit shops of the city of Montería, and ruminal content was collected from the slaughterhouse in Cereté, both localities in the department of Córdoba.
Peels from papaya, pineapple and banana were selected because these are the fruits found in the department of Córdoba and which are available during the whole year, and therefore, waste production is higher compared to others fruits.
Clarification of the ruminal content started by separating manually solid from liquid part, this last was filtered to obtain a homogeneous liquid, that was 40 % v/v. Subsequently, weighting, cutting and liquefying of peels was carried out and adjusting pH to 6.34 ± 0.03 with sodium bicarbonate solution for liquid and solid media. Concentrations of 25, 50, and 75 % w/v of peels were used to obtain three different biopreparations of the ruminal content-fruit peels media.
To determine the effect of the media on growth and biomass production of the wild strain, 1 mL of the strain suspension was inoculated in 30 mL glass flasks with the media at each of the mentioned three concentrations and incubated for 48 hour at room temperature (±28 °C) with constant stirring in a shaker. Growth was determined by the serial dilutions method and viable cell count was observed in colony-forming units (cfu). A completely randomized design of the treatments was used in this experiment.
Additionally, the following chemical analysis were carried out in triplicate to the best medium (the one that allowed the highest microbial growth):
Carbohydrates were determined with a Perkin Elmer Lambda spectrophotometer. The sample was treated with phenol 5 % and concentrated sulfuric acid in 1:5 ratio and measured at 490 nm wave length. Calibration curve with the equation Abs= 0.0127 Conc + 0.9996 (R2 = 0.99) was used.
Proteins were determined using the Kjeldahl method 955.04 (AOAC, 1990).
Ash percentage was determined using the method 942.05 (AOAC, 1990).
Humidity percentage was determined by the method 930.15 (AOAC, 1990).
For determination of elements potassium, sodium, copper, manganese, iron, and zinc, a Perkin Elmer 3110 atomic absorption equipment was used at 767, 589, 325 280, 249 and 214 nm wave lengths, respectively. Elements sulfur and phosphorous were determined using a Perkin Elmer LX spectrophotometer at 420 and 660 nm wave lengths, respectively. Calcium and magnesium were determined through complexo-metry (APHA, 1998).
C) Evaluation of strain viability over time
To evaluate the wild strain viability, 9 mL of the best medium were used in glass flasks. Every flask was inoculated with 1 mL of the wild strain suspension, stored at ±28 °C with constant stirring in a shaker. At times 0, 15, 30, 45 and 60 days (considering at day 0 the wild strain 48 hours of being incubated) viable cell count was carried out (Marin et al., 2009).
The results for the in vitro tests were analyzed by ANOVA and Tukey test after confirming the assumptions of normality of the data (Shapiro-Wilk test) and homogeneity of variance (Bartlett test). The data from the experiment of the effect of three media on growth and biomass production of the wild strain did not conform normality nor homogeneity of variance, thus a non parametrical test was used (Kruskal-Wallis and Dunn test). All analyses were performed using the R statistical analysis program (R Development Core Team. Auckland University, New Zealand)
The results of the wild strain growth and viability over time were presented as figures with standard errors.
Results AND DISCUSSIoN
Results obtained in each in vitro testing to the wild strain of S. cerevisiae are shown below.
Tolerance to bile salts. Wild strain kept population concentration thereof in the same logarithmic unit with respect to the initial population (107 cfu·mL-1), therefore, it is capable of tolerating and maintaining the range of concentration of bile salts tested as observed in the counts (Table 1).
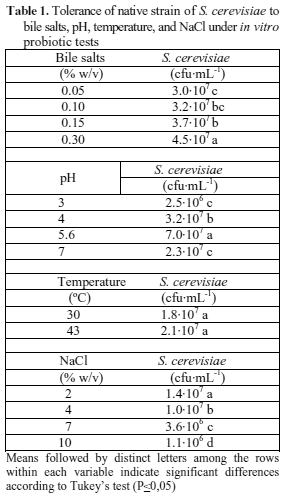
The highest average cell growth was obtained at a concentration of 0.2 % of bile salts, and the lowest at a concentration of 0.05 %. The test showed that there was no significant difference between the concentrations 0.15 and 0.1, and between 0.1 and 0.05.
According to Ortiz et al. (1997), S. cerevisiae has integral ATP-binding membrane proteins (ABC proteins), responsible for the translocation of bile salts and may efficiently transport conjugated bile acids. Another mechanism through which yeast is resistant to high concentration of bile salts is based on the accumulation of polyols and glycerol, as mechanism to regulate osmotic pressure in the cell (Moser & Savage, 2001), suggesting that yeast species which tolerate bile salts contribute to the function of microorganisms in the gastrointestinal tract (Duncan et al., 2013). Maintenance of probiotic microorganism population composing a commercial biopreparation should be in the range of 106-108 cfu·mL-1, when reaching target cells, to assure they may act on the host (Cavazzoni et al., 1998). It suggests that it is necessary for an in vivo evaluation to start from an inoculum with higher biomass concentration, in order to assure that yeast facing bile salts, having antimicrobial activity, decrease their cell viability, but still maintaining the necessary concentration to affect positively the host (Salmines et al., 1999).
Wild strain of S. cerevisiae showed to be a microorganism with capacity to tolerate bile salt concentrations from 0.05 to 0.3 % (w/v), keeping a constant number of cfu and being able to develop metabolic activities thereof, without being inhibited or altered, characteristic which is important for formulating a probiotic preparation.
pH Tolerance. There were significant differences in average cell growth at different pH levels and a growth decrease occurred as the pH moved away from a mean value (Table 1). The highest cell growth was obtained with pH 5.6 and the lowest was obtained with pH 3. When exposed to this latter pH, there was a large decrease of the wild strain population with respect to the initial one as observed by the net decrease in one logarithmical unit (107 to 106), whereas for the others, the order of magnitude of the population was maintained. A probiotic microorganism population should be in 108 cfu·mL-1 when administered and when reaching enterocytes, and it should be viable, with a maximum viability loss of two logarithmic units (Cavazzoni et al., 1998). Hence, it is required to have an inoculum with higher microorganism concentration as mentioned before.
Tolerance to pH may rely on two types of Na+/H+ anti-transporters in the yeast: Nha1p, found in the plasmatic membrane and Nhx1p, both located in the prevacuolar endosomal compartment (Membré et al., 1999). These proteins catalyze monovalent cation exchange (Na+ or K+) and H+ through membranes, then they regulate cation concentrations and pH at cytoplasmic level and organelles (Mitsui et al., 2005; Ohgaki et al., 2005). Another possible regulation mechanisms is represented by ATPase located in the cytoplasmic membrane, which may create a proton electrochemical gradient leading to the secondary solute transport which is implied for maintaining pH close to neutral (Viegas et al., 1998; Sychrovae et al., 1999).
Tolerance to change in temperature. The response of the wild strain under two different temperatures showed that the increase from 30 to 43 ºC did not affect the microorganism growth, maintaining the logarithmic population regarding the initial one (Table 1). Thereby the wild strain met another criterion to be considered as a suitable probiotic for the elaboration of a biopreparation.
Temperature is a relevant factor because from the microbiological point of view, microorganisms have optimal temperatures for growth thereof obtaining the highest yield. This temperature is often close to the maximum tolerated and varies according to the type of microorganism. In yeasts, temperature affects their capacity to split sugars, reproduction and cell growth (Manovacía et al., 2008). In this test, the temperature used was in the appropriate range for S. cerevisiae growth.
Tolerance to high NaCl concentrations. There were highly significant differences in the average absorbance at each of the NaCl concentrations (Table 1). The wild strain proved tolerance for concentrations of 2 and 4 % w/v NaCl (Table 1), but it showed a decrease in tolerance and decrease in the growth rate for the higher salt concentrations of 7 and 10 % w/v.
According to Martinez (2016), when S. cerevisiae faces a high osmolarity condition, suffers an immediate change in cell volume due to the water loss in the cytosol. Dehydration is a short process, which takes very short time, and it is partially compensated by the influx of water from the vacuole as it accumulates ions for the benefit of the cytoplasm and organelles (Serrano, 1996). This influence is shown as an internal response to the differences in osmotic concentrations between the cytoplasm and the interior of the vacuole that tries to maintain the hydration of the cell, also helped by the contact surface with the cytoplasm that allows the passive movement of solutes and water to survive and maintain growth (Blomberg, 2000). Dehydrated cells may recover stiffness provided stress severity is physiologically acceptable. Cell proliferation may resume after a conditioning period (Tao et al., 1999). At the same time, mechanisms involved in stress resistance are induced, such as increase in the glycerol intracellular concentration and induction of products that participate in production of protecting proteins and readjustment in levels of carbohydrate, lipids and amino acids (Rep et al., 2000). For all the above mentioned, it can be stated that wild strain has the capacity to condition and stabilize itself before high bile acid concentration, allowing the maintenance of population when it is introduced as a beneficial microorganism resisting different salinity conditions.
Antagonism test. Antagonism test with respect to E. coli showed that, in vitro, yeast does not produce antimicrobial substances which might spread to the medium and being capable of counteracting growth of the pathogen evaluated.
It was observed, however, that the wild strain has some advantage for the competitive exclusion, which suggests that they could be more resistant to gastrointestinal environment being colonization and multiplication thereof more productive compared to pathogens. This assumption may be valid if we consider that it is difficult to evaluate antagonism in vitro because the difficulty to promote gastrointestinal conditions.
Production of gas from glucose. Wild strain showed poor gas production; this property is very important because diet of hosts is commonly based on glucose and if a gas producing probiotic is used, digestive problems will appear and it may be considered as a loss of energy for the host, because it is potential energy not used for metabolic processes of the microorganisms.
Microbial growth in the media from ruminal content and fruit peels. Mean values of the results obtained for growth of the wild strain of S. cerevisiae, under previously established conditions, in the culture medium from ruminal content enriched with fruit wastes in concentrations of 25, 50, and 75 % are summarized in Table 2.
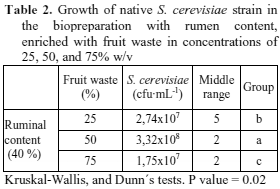
It is observed that the wild strain has better population growth in the biopreparation with 50 % w/v fruit waste content than concentrations of 25 and 75 % of wastes.
Based on the above, the ruminal content medium enriched with fruit wastes 50 % w/v was selected for chemical characterization because this medium was the one that allowed the greatest growth of the wild strain S. cerevisiae.
Chemical analysis of the best medium from ruminal content and fruit peels. The results of the chemical characterization made to the medium with 50 % w/v fruit waste concentration shows that it counts on the nutritional requirements necessary for the good growth of the wild strain of S. cerevisiae in the elaboration of a biopreparation (Table 3).
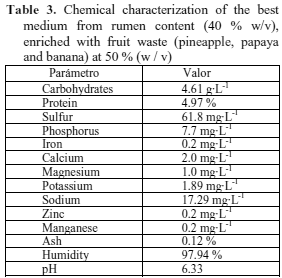
The carbohydrate (4.61 g·L-1), protein (4.97 %) and phosphorus (7.7 mg·L-1) contents were higher than those found in different media such as Psidium araca (Lara et al., 2008), Ipomoea batatas (Lobaina Rodríguez et al., 2007), or a mix of vegetable residues (Lara et al., 2010).
On the other hand, mineral microelements such as sulfur and sodium that are fundamental for microorganism growth (Bridson, 1994) were found in important amounts in the medium. The sulfur and sodium concentrations (61.8 and 17 mg·L-1) surpassed the values of 6. mg·L-1 and 14.93 reported by Lara et al. (2008; 2010) for those nutrients, respectively. These comparisons demonstrate good nutritional contents that can favour the growth of S. cerevisiae.
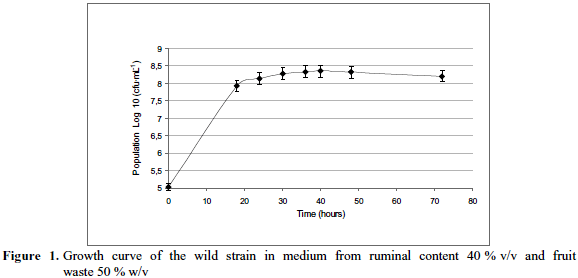
Evaluation of the strain viability over time. The Figure 1 represents the population of viable cells existing in the medium from ruminal content (40% v/v) with residues of the mixtures fruits (papaya, pineapple and banana) at a concentration of 50% w/v. According to plotted biomass production logarithm data, it is observed that the strain S. cerevisiae adapted quickly to the culture medium evaluated and remained in logarithmic phase increasing initial population by 60 % en 18 hours, thereby achieving maximum growth at 40 hours, to subsequently decrease; decrease which is probably produced by the exhaustion of nutrients present in de medium.
In Figure 2, the trend of population viability of the wild strain of S. cerevisiae can be observed every 15 days during two months, in the best medium, i.e. the medium from ruminal content (40% v/v), with fruit wastes (papaya, pineapple and banana) at 50% w/v concentration.
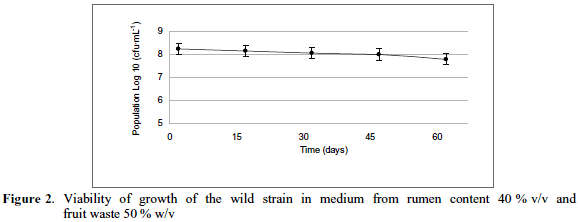
In this research, the three results obtained after 45 days of storage maintained the log scale of 108 cfu·mL-1, with a decrease in counts made in last 15 days to the log scale 107; it i supposed that refrigeration may have had consequences in the decrease, as well as the decrease of nutrients available in the medium. According to nutritional viability parameters of probiotics in food (FAO, 2016), S. cerevisiae met the requirement of viability, which should be higher than106. These results may confirm that the biopreparation selected is a good vehicle for probiotic administration. Organic waste used in this study have been subject of several individual research, in order to take advantage of nutritional value thereof, finding the production of flours, nutritional blocks and compost (Yato & Orihuela, 2015; Seema, 2015), in the case of ruminal content; and for the peels of different fruits, elaboration of jams, flours (Gil et al., 2011), dietary fiber (Gutiérrez et al., 2002) and fermentation products (Saval, 2012).
Conclusions
Elaboration of a biopreparation from clarified ruminal content 40% v/v, enriched with peels from three fruits (pineapple, papaya and banana) 50 % w/v, produced in slaughterhouses (Cereté, Colombia) and fruit shops (Montería, Colombia) respectively, with the strain Saccharomyces cerevisiae constitute a viable byproduct, which may contribute to the improvement of environment, by decreasing by-products caused in the livestock and fruit production.
Wild strain of S. cerevisiae showed favorable results for tests of tolerance to bile salts, changes in temperature, high salt concentrations, additionally to showing a competitive exclusion competence. Wild strain showed poor gas production, which a very important property because the consumption of glucose by the yeast causes decreasing bioavailability of this carbohydrate for the host organism.
Best growth and maintenance of the wild strain of species S. cerevisiae was observed in the clarified ruminal content medium (40 % w/v) and fruit peels (pineapple, papaya and banana) 50 % w/v. And chemical analysis showed that the medium has the necessary nutrients for the yeast in the elaboration of a biopreparation.
Acknowledgement
The authors thank Universidad de Córdoba (Montería, Colombia) and Corporación Universitaria del Caribe (CECAR) (Sincelejo, Colombia).
LITERATURA CITADA
1. AOAC. 1990. Methods of Analysis. Association of Official Analytical Chemist. Washington, D.C. [ Links ]
2. Abreu-Abreu, A. 2012. Prebióticos, probióticos y simbióticos. Rev Gastroenterol Mex. 77: 26-28. [ Links ]
3. APHA. 1998. Standard methods for the examination of water and wastewater. American Public Health Association. Washington, D.C. [ Links ]
4. Blomberg, A. 2000. Metabolic surprises in Saccharomyces cerevisiae during adaptation to saline conditions: questions, some answers and a model. FEMS Microbiol Lett. 182(1): 1-8. [ Links ]
5. Bridson, E. 1994. The Development, Manufacture and Control of Microbiological Culture Media. Unipath. Bedford, UK. [ Links ]
6. Cavazzoni, V., A. Adami and G. Castrovilli. 1998. Performance of broiler chickens supplemented with Bacillus coagulans as probiotic. British Poultry Science, 30: 526-529. [ Links ]
7. DANE (Dirección Nacional de Estadística). 2014. Censo Nacional Agropuecuario. http://www.dane.gov.co/index.php/estadisticas-por-tema/agropecuario/ (visited 07/28/2017). [ Links ]
8. Drisko, J., C. Giles and B. Bischoff. 2003. Probiotics in health maintenance and disease prevention. Altern. Med. Rev. 8(2): 143-155. [ Links ]
9. Duncan, S., K. Scott, A. Ramsay, H. Harmsen, W. Gjalt and S. Colin. 2013. Effects of alternative dietary substrates on competition between human colonic bacteria in an anaerobic fermentor system. Appl. Env. Microbiol. 69: 1136-1142. [ Links ]
10. FAO. 2006. Livestock's long shadow. Environmental issues and options. Food and Agriculture Organization of the United Nations. Rome. 290 p. [ Links ]
11. FAO. 2016. Probiotics in animal nutrition: production, impact and regulation. Animal Production and Health. Paper 179. Rome. [ Links ]
12. García, M., Y. López and A. Carcassés. 2012. Empleo de probióticos en los animales. Sitio Argentino de Producción Animal. 8 p. http://produccion-animal.com.ar (visited 07/28/2017). [ Links ]
13. Gil, M., L. Vélez, L.d. Millán, M. Acosta, A. Díez, N. Cardona and G. Villa. 2011. Desarrollo de un producto de panadería con alto valor nutricional a partir de la harina obtenida del banano verde con cáscara: una nueva opción para el aprovechamiento de residuos de la industria de exportación. Producción+Limpia 6(1): 96-107. [ Links ]
14. Guerrero, E. and F. Ramírez. 2004. Manejo ambiental de residuos en mataderos de pequeños municipios. Scientia et Technica 10(26): 199-204. [ Links ]
15. Gutiérrez, E., G. Medina, M. Roman, O. Florez and O. Martínez. 2002. Obtención y cuantificación de fibra dietaria a partir de residuos de algunas frutas comunes en Colombia. Vitae 9(1): 5-14. [ Links ]
16. Lara, C. 2008. Chemical composition of a culture medium from bitter guava (Psidium araca) and its relation to the ruminals microorganisms nutrition. Revista Colombiana de Biotecnología, 2: 44-49. [ Links ]
17. Lara, C., L. García and L. Oviedo. 2010. Medio de cultivo utilizando residuos-sólidos para el crecimiento de una bacteria nativa con potencial biofertilizante. Revista Colombiana de Biotecnología, 12 (1): 103-112. [ Links ]
18. Leiva, S., M. Yáñez, L. Zaror, H. Rodríguez and H. García. 2004. Actividad antimicrobiana de actinomycetes aislados desde ambientes acuáticos del sur de Chile. Revista Médica de Chile, 132: 151-159. [ Links ]
19. Lopera, M., J. Homez, M. Ordoñez and H. Pabon. 2009. Guía Ambiental Hortifruticula de Colombia. Edit. Nuevas Ediciones. Bogotá. [ Links ]
20. Lobaina, T., C. Rodríguez and R. Zhurbenko. 2007. Caracterización de un extracto de Ipomoea batatas para ser utilizado en calidad de base nutritiva en medios de cultivo. Rev. Cubana Med. Trop. 59(3): 218-26. [ Links ]
21. Manovacía, N., A. Moreno, O. Mayorga and R. Barahona. 2008. Evaluation of the nutrient content and biomass production in Colombian and commercial yeast strains. Revista Facultad Nacional de Agronomía (Medellín) 61(2): 4542-4553. [ Links ]
22. Marín, Z., M. Cortes and O. Montoya. 2009. Evaluación de la viabilidad de crecimiento de la cepa nativa Lactobacillus plantarum lpbm10 y la cepa comercial Lactobacillus casei atcc 393 en pulpa de uchuva y en solución isotónica de glucosa. Vitae 16(2): 210-217. [ Links ]
23. Martínez, E., A. Torregroza, Ana. Torregroza and D. Mogollón. 2016. Efecto de la deshidratación osmótica-microondas sobre propiedades fisicoquímicas del mango (Mangifera indica L.) variedad Corazón. Agronomía Colombiana, 34: 1236-1239. [ Links ]
24. Membré, J., M. Kubaczka and C. Chene. 1999. Combined effects of pH and sugar on growth rate of Zygosaccharomyces rouxii, a bakery product spoilage yeast. Appl. Env. Microbiol. 69(2): 1136-1142. [ Links ]
25. Mitsui, K., H. Yasui, N. Nakamura and H. Kanazawa. 2005. Oligomerization of the Saccharomyces cerevisiae Na+/H+ antiporter NHa1p: Implications for its antiporter activity. Biochim. Biophys. 1720: 125-136. [ Links ]
26. Moser, S. and D. Savage. 2001. Bile salt hydrolase activity and resistance to toxicity of conjugated bile salts are unrelated properties in Lactobacilli. Appl. Env. Microbiol. 67(8): 3476-3480. [ Links ]
27. Mossel, D., B. Moreno and C. Struijk. 2003. Microbiología de Los Alimentos. Edit. Acribia. Zaragoza, Spain. [ Links ]
28. Ohgaki, R., N. Nakamura, K. Mitsui and H. Kanazawa. 2005. Characterization of the ion transport activity of the budding yeast Na+/H+ antiporter, NHA1P, using isolated secretory vesicles. Bioch. Biophys. Acta. 1712: 185-196. [ Links ]
29. Ortiz, D., M. St.Pierre, A. Abdulmessih and I. Arias. 1997. A yeast ATP-binding cassettetype protein medianting ATP-dependent bile acid transport. J. Biol. Chem. 272: 15358-15365. [ Links ]
30. Pascual, M. and V. Calderón. 2000. Microbiología Alimentaria. Metodología Analítica para alimentos y bebidas. Edit. Díaz de Santos. Madrid. [ Links ]
31. Rendueles, M. and M. Díaz. 2014. Biotecnología industrial. Arbor CPC 190-768(a155): 13 p. [ Links ]
31. Rep, M., M. Krantz, J. Thevelein and S. Hohmann. 2000. The transcriptional response of Saccharomyces cerevisiae to osmotic shock. Hot1p and Msn2p/Msn4p are required for the induction of subsets of high osmolarity glycerol pathway-dependent genes. J. Biol Chem. 275(12): 8290-300. [ Links ]
33. Rodón, A., L. Samaniego, R. Bocourt, S. Rodríguez, G. Milián, M. Ramilla and M. Pérez. 2004. Ciencia y Tecnología Alimentaria, 6(1): 56-63. [ Links ]
34. Rubio, A. 2008. Identificación preliminar in vitro de propiedades probióticas en cepas de S. cerevisiae. MVZ 13(1): 1157-1169. [ Links ]
35. Salmines, S., A. Ouwenhand and Y. Lee. 1999. Probiotics: how should they be defined. Trends of Food Science and Technology, 10: 107-110. [ Links ]
36. Saval, S. 2012. Aprovechamiento de residuos agroindustriales: pasado, presente y futuro. BioTecnología 16(2): 14-46. [ Links ]
37. Seema, R. 2015. Probiotics in valorization of innate immunity across various animal models. Journal of Functional Foods, 14: 549-561. [ Links ]
38. Serrano, R. 1996. Salt tolerance in plants and microorganisms: toxicity targets and defense responses. Int. Rev. of Cyt. 165: 1-52. [ Links ]
39. Sychrovae, H., J. Ramírez and A. Peña. 1999. Involvement of Nha1 antiporter in regulation of intracellular pH in Saccharomyces cerevisiae. Microbiol. Let. 171(2): 167-172. [ Links ]
40. Tao, W., J. Deschenes and J. Fassler. 1999. Intracellular glycerol levels modulate the activity of Sln1p, a Saccharomyces cerevisiae two-component regulator. J Biol Chem. 274(1): 360-367. [ Links ]
41. Uicab-Brito, L. A. and C.A. Sandoval. 2003. Uso del contenido ruminal y algunos residuos de la industria cárnica en la elaboración de composta. Tropical and Subtropical Agroecosystems 2: 45-63. [ Links ]
42. Valdovinos, M. 2013. Microbiota intestinal en los trastornos digestivos. Probióticos, prebióticos y simbióticos. Revista de Gastroenterología de México 78: 25-27. [ Links ]
43. Viegas, C., P. Almeida, M. Cavaco and I. Sá-Correia. 1998. The H-ATPase in the plasma membrane of Saccharomyces cerevisiae is activated during growth latency in octanoic acid-supplemented medium accompanying the decrease in intracellular pH and cell viability. Appl. Environ. Microbiol 64(2): 779-783. [ Links ]
44. Yato, G. and J. Orihuela. 2015. Uso de fuentes no convencionales de nitrógeno en la fertilización del maíz (Zea mays L.) en Cañete (Perú). I: Rendimiento y extracción de N, P y K. Ecología Aplicada 14(2): 157-162. [ Links ]
