Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkBoletín de Malariología y Salud Ambiental
versión impresa ISSN 1690-4648
Bol Mal Salud Amb v.47 n.1 Maracay ene. 2007
Wyeomyia trujilloi Pulido y Sutil, 1981, nuevo sinónimo de Wyeomyia bicornis (Root, 1928) (Culicidae: Sabethini), con redescripción de la pupa parte de la larva
Juan-Carlos Navarro1* & Jonathan Liria2
1 Lab. Biología de Vectores, Instituto de Zoología Tropical, Universidad Central de Venezuela, Apdo. 47058, Caracas 1041-A.
2 Departamento de Biología, Facultad Experimental de Ciencias y Tecnología, Universidad de Carabobo. Campus Bárbula, Naguanagua. Estado Carabobo.
*Autor de correspondencia: jnavarro@ciens.ucv.ve
Wyeomyia (Wyeomyia) trujilloi Pulido & Sutil, 1981 es reconocido como sinonimia de Wyeomyia (Nunezia) bicornis (Root, 1928), con base en la morfología de larva, piezas bucales, pupa y genitales del macho en comparación con el holotipo de Wy. bicornis, topotipos y conforme con un análisis del holotipo de Wy. trujilloi. Se provee una explicación e ilustración sobre las características a nivel subgenérico de Nunezia. Para Wy. bicornis la morfología cefálica de la larva es redescrita. La morfología y quetotaxia de la pupa y las piezas bucales de la larva son descritas e ilustradas por primera vez.
Palabras claves: Fitotelmata, morfología, mosquitos, Nunezia, Sabethini, Wyeomyia.
Wyeomyia trujilloi Pulido & Sutil, 1981, new synonym of Wyeomyia bicornis (Root, 1928) (Culicidae: Sabethini), with redescription of the pupa and part of the larva
SUMMARY
Wyeomyia (Wyeomyia) trujilloi Pulido & Sutil, 1981, is recognized as the synonym of Wyeomyia (Nunezia) bicornis (Root 1928), based on larval chaeototaxy and mouthparts morphology, pupae and male genitals in comparison and analysis with Wy. bicornis holotype, topotypes and Wy. trujilloi holotype. Discussion and characteristics illustration about subgeneric level are provided. The larval mouthparts morphology and pupae chaetotaxy are described and illustrated by first time, and head of larvae chaetotaxy are re-described.
Key words: morphology, mosquitoes, Nunezia, Phytothelmata, Sabethini, Wyeomyia.
Recibido el 08/11/2005 Aceptado el 15/12/2006
INTRODUCCIÓN
La Tribu Sabethini está integrada por 360 especies (204 y 160 en el Nuevo Mundo y Viejo Mundo, respectivamente) de acuerdo con Knight & Stone (1977) y Guimarães (1997). No obstante, esta cifra pudiera estar fuertemente subestimada de acuerdo a revisiones realizadas a la colección del Museo de Historia Natural, Smithsonian Institution (Judd, Com. pers.).
Igualmente, el poco conocimiento y la confusión existente en la sistemática de Sabethini es claramente observada en intentos recientes de un ordenamiento y clasificación natural del grupo (Zavortink, 1985; Harbach, 1991a,b, 1994, 1995; Harbach & Peyton, 1990, 1991, 1993; Harbach & Petersen, 1992; Judd, 1996; Harbach & Peyton, 2000).
Judd (1996, 1998) sugiere que esta clasificación, en algunos casos no natural, es consecuencia de la escasa comparación entre fases inmaduras en las descripciones originales, del pobre tratamiento sistemático y la carencia de claves adecuadas para una correcta y eficiente identificación de diferentes especies tratadas en Lane y Cerqueira (1942) y Lane (1953).
Particularmente, el género Wyeomyia Theobald no escapa a este caos sistemático. Este género incluye más de 100 especies, 37 de ellas en Venezuela (Sutil, 1980) que son difíciles de reconocer en su mayoría. Infragenéricamente, está representado por 15 subgéneros (Judd, 1998; Porter & Wolf, 2005; Motta & Lourenço-de-Oliveira, 2005) luego que recientemente, miembros de este género han sido transferidos a otros géneros, se han creado tres subgéneros nuevos, e inclusive especies del género Sabethes Robineau-Desvoidy han sido trasladados hacia Wyeomyia (Zavortink, 1985; Harbach & Peyton, 1990, 1991; Judd, 1998).
El género Wyeomyia es uno de los taxa de Sabethini más numeroso y quizás menos estudiado con respecto a su clasificación natural debido al desconocimiento de las fases inmaduras. A nivel subgenérico, Nunezia Dyar es muy poco conocido, siendo hasta el presente un taxón con sólo dos especies, Wy. lateralis Petrocchi, 1927 y Wy. bicornis (Root, 1928), esta última, la especie tipo del subgénero y el único representante conocido en fase larval.
El presente trabajo es parte y consecuencia de un estudio en gran escala acerca de la fauna de mosquitos en Parques Nacionales de Venezuela, cuyas fases inmaduras se desarrollan en plantas Fitotelmata, las cuales poseen estructuras morfológicas (brácteas, espatas, axilas, etc.) que pueden almacenar agua de lluvia en donde se desarrollan una variedad de insectos acuáticos (Varga, 1928; Fish, 1983). Los Sabetinos representan cerca del 50% de la fauna de Culicidae de las fitotelmata en Venezuela (Navarro et al., 2007) y especies del género Wyeomyia son importantes cohabitantes en diferentes especies y tipos de estas plantas.
En el inventario (identificación, revisión) de especies nos hallamos la curiosa descripción de Wyeomyia (Wyo.) trujilloi por Pulido & Sutil (1981) como una especie nueva, cuya morfología no concordaba con las características del subgénero al cual fue asignado por estos autores, y con el tipo de criadero en el cual estaba señalado en la descripción bionómica. Luego de un análisis meticuloso determinamos que Wy. trujilloi debe ser sinonimizada con Wy. (Nunezia) bicornis, lo cual es el propósito de este trabajo.
Igualmente, se realizó la descripción para Wy. bicornis de la pupa y de las piezas bucales de la larva, las cuales no estaban descritas, así como parte de la cabeza y sifón respiratorio las cuales presentaban un pobre tratamiento taxonómico. Finalmente, se proporcionan explicaciones morfológicas, bionómicas y biogeográficas en las cuales se basó la sinonimia con el objetivo de realizar un aporte al conocimiento de la historia natural y sistemática de los sabetinos en Venezuela.
MATERIALES Y MÉTODOS
Material examinado
Se analizó el holotipo (macho) de Wy. trujilloi Pulido & Sutil con los números, 21-40-1 y 21-40-2 depositado en la colección del Laboratorio de Morfología de Insectos, División de Endemias Rurales, Dirección de Malariología y Saneamiento Ambiental, Maracay, Estado Aragua, Venezuela, (DERM sensu Knight & Stone, 1977; Guimarães, 1997) colectada en Altamira, Municipio Altamira, Estado Barinas. Este holotipo consiste de los genitales del macho montados en lamina (21-40-2), una exuvia de pupa (Pe, 21-40-2) y una exuvia de larva (Le, 21-40-1).
También fue revisado el holotipo (macho) de Wy. bicornis (Root) depositado en el Museo de Historia Natural (NMNH sensu Knight & Stone 1977; Guimarães 1997), en el Walter Reed Biosystematic Unit, Museum Support Center, Washington DC, Estado Unidos. El holotipo (# 44162) consiste de los genitales masculinos montados en lámina, con etiqueta: Venezuela / Ocumare / July 6 1927 / No 108-1.
Para el análisis comparativo se utilizaron los holotipos de Wy. trujilloi y Wy. bicornis, así como ejemplares de Wy. bicornis de la colección del Laboratorio de Biología de Vectores, Museo de Biología, Universidad Central de Venezuela (LBVMB- UCV-IX sensu Guimarães, 1997) colectadas en: 1. Localidad tipo (Topotipos) Parque Nacional (PN) Henri Pittier (Estación Biológica Rancho Grande); vía Ocumare de La Costa, con los números 500-Le, 508-Le, Pe, 509a,b-L4. Fitotelmata hospedadora: Aechmea sp. (Bromeliaceae). Colectores: JC Navarro y H Piñango. Fecha: 13 / V / 1994. Determinación: J. C. Navarro y J. Liria.
2. Sierra de San Luis, PN Juan Crisóstomo Falcón, Edo. Falcón, 2.000 m snm, con los números 722, L- 023.1. Fitotelmata hospedadora: Vriesea platynema (Bromeliaceae). Colectores: Equipo LBV. Fecha: IV / 1994. Determinación: J. C. Navarro, J. Ingunza y J. Liria.
3. Páramos Batallón y La Negra, PN Juan Pablo Peñaloza, Edo. Táchira 1.445-1.475 msnm, con los números 1048-L4, 1106-Le,P. Fitotelmata hospedadora: Aechmea sp. (Bromeliaceae). Colectores: Equipo LBV. Fecha: X / 1995. Determinación: J. C. Navarro y J. Liria.
4. Monte Zerpa, Mérida, Edo. Mérida, PN Sierra Nevada, 2.020 msnm, con el número 906-L4. Fitotelmata hospedadora: Guzmania mitis. (Bromeliaceae). Colectores: Equipo LBV. Fecha: 09 / III / 1995. Determinación: J. C. Navarro y J. Liria. Especies de Nunezia no descritas Wy. (Nunezia ) sp. 3
5. Guanay Tepui, Edo. Amazonas, 1.250 m, bajo los números 691-Le, Pe; 705-Le; L-023.3-L4; L-023.4-L4. Fitotelmata hospedadora: Brocchinia tatei (Bromeliaceae). Colectores: J. C. Navarro y J. Ingunza. Fecha: 16 / II / 1995.
6. Yutaje Tepui. Edo. Amazonas, 1.750 m bajo los números 676-Le,Pe; 780-Le,Pe; 1704-L4. Fitotelmata hospedadora: Brocchinia tatei y Vriesea rubra. (Bromeliaceae). Colectores: J. C. Navarro y J. Ingunza. Fecha: II / 1995. Wy. (Nunezia) sp. 2
7. Auyantepui, PN Canaima, Edo. Bolívar, 1.730 m., con los números 433-L4. (Harbach y Navarro 1996). Fitotelmata hospedadora: Brocchinia tatei (Bromeliaceae). Colectores: J. C. Navarro y J. Liria. Fecha: 03 / II / 1994. Wy. (Nunezia) sp. 1
8. La Gran Sabana (La Escalera), PN Canaima, Edo. Bolívar, 1.040 m, con los números 400 y 405 (L4). Fitotelmata hospedadora: Brocchinia sp. (Bromeliaceae). Colectores: Equipo LBV. Fecha: IX / 1993.
Preparación de muestras
Las fases inmaduras de estos ejemplares se preservaron en etanol al 80%, se transparentaron en KOH al 10% (Harbach, 1977; Harbach & Knight, 1977) y se transfirieron a una batería de deshidratación alcohólica (Belkin, 1962).
Las piezas bucales se disecaron en una gota de Hoyer o una mezcla de bálsamo de Canadá y fenol (Wirth, 1961), bajo el microscopio estereoscópico. Las láminas (conteniendo larvas, sifón respiratorio y piezas bucales) se incubaron en estufa a 45-50°C durante dos a tres semanas (Harbach & Peyton, 1993; Pérez & Navarro, 1996; Liria & Navarro, 1999) Para la denominación de las estructuras se siguió la nomenclatura y el sistema de abreviaturas propuesto por Harbach & Knight (1980). Las abreviaturas genéricas y subgenéricas siguen el sistema de Reinert (2001).
Las estructuras de interés se fotografiaron por medio de un microscopio óptico Leitz Wetziar laborlux con cámara automática de 0.32X, modelo MPS51-Wild y controlador MPS45-Wild.
Análisis cuantitativo de datos
Ordenamiento: Se realizó un Análisis de Componentes Principales (ACP) con el programa de computación PAST 1.3 (Hammer & Harper, 2004), como técnica de ordenamiento de especies para verificar estadística y gráficamente la diferenciación morfológica a nivel de subgnéneros y comprobar además, la exclusión de Wy. trujilloi del subgénero Wyeomyia. Se utilizaron 31 caracteres (Apéndice I) de la larva (28 de la quetotaxia y tres morfométricos). En el análisis se incluyeron especies como Wy. (Wyo.) arthrostigma (Lutz 1905) (130.1-1.4, Torondoy, Edo. Mérida) y Wy. (Dec.) felicia (Dyar & Nuñez-Tovar 1927) (288-L4, Sierra de San Luis, Edo. Falcón), los topotipos de Wy. bicornis y el holotipo de Wy. trujilloi como especies problema, y tres especies no descritas del subgénero Nunezia pertenecientes a las cinco localidades antes señaladas.
Filogenético: Se realizó un análisis cladístico con los programas de computación NONA + WINCLADA (Goloboff, 1996; Nixon, 1999) con base a los caracteres larvales anteriormente señalados y previa codificación (Apéndice II) mediante el método de Gap-Weighting (Thiele, 1993; Kitching et al. 1998; Rae, 1998). Se emplearon las especies de Nunezia, Wy. trujilloi, Wy. arthrostigma, Wy. felicia y Wy. (Phoniomyia) deanei (Lourenço-de-Oliveira 1983), y como grupo externo a Onirion imparis Peyton & Harbach, 2000.
RESULTADOS Y DISCUSIÓN
Luego del análisis comparativo entre los holotipos de Wy. trujilloi Pulido & Sutil (1981) con Wy. bicornis (Root, 1928), llegamos a la conclusión de sinonimizar la primera con la última, señalada en conformidad con evidentes características presentes en la larva y sus partes bucales, así como en los genitales del macho de Wy. trujilloi que corresponden con la descripción de Wy. bicornis.
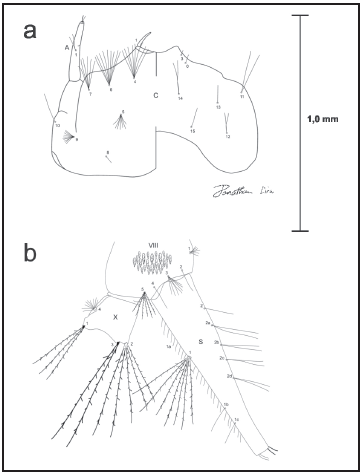
Extrañamente, Pulido & Sutil (1981) en su descripción, incluyeron a Wy. trujilloi como perteneciente al subgénero Wyeomyia, sin señalar las características utilizadas para llegar a esa errónea conclusión: sic. Wy. trujilloi esta agrupada en el subgénero Wyeomyia porque la proboscis es casi tan larga como el fémur anterior. Además, las características de la genitalia del macho son inconfundibles (págs. 222 y 226). Esta aseveración, fue probablemente la que llevó a estos autores a no verificar la clara asociación de la larva (particularmente en el sifón, seta 1-S) (Fig. 1b) con la única especie de Nunezia presente en el país y cuya localidad tipo precisamente está en Venezuela (Edo. Aragua). Una posible explicación a esto se relaciona con las figuras mostradas en Lane (1953), texto ampliamente utilizado para la identificación de mosquitos Suramericanos, las cuales no corresponden con la descripción original en Dyar (1928: 51) para Wy. bicornis, obviando la presencia del mechón de setas en 1-S: sic. a paired tuft toward base and two single hairs toward apex característica no descrita por Pulido & Sutil (aunque si dibujada) y que claramente lo presenta Wy. trujilloi.
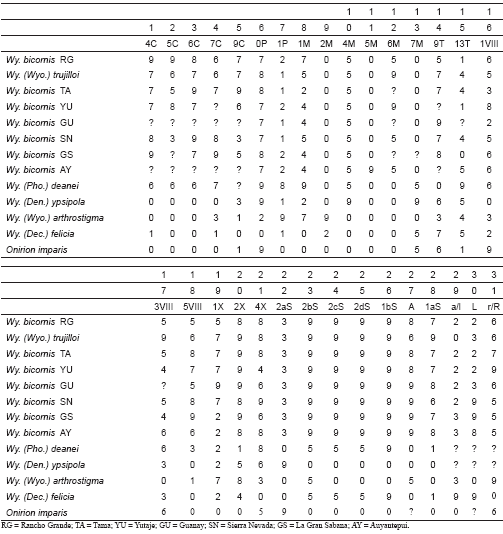
Tabla I. Quetotaxia para la cabeza y segmento abdominales VIII y X para la larva de IV estadio de Wyeomyia (Nunezia) bicornis, (n=4).
| Seta No. | Cabeza | Segmentos Abdominales | |
| VIII | X | ||
| 0 | 1 | 1 | - |
| 1 | 1 | 8,7(8) | 3,4(4) |
| 2 | 1 | 1 | 7,8(8) |
| 3 | 1 | 6,7,9(7) | 2 |
| 4 | 10,8(9) | 1 | 7,8(8) |
| 5 | 9,6(8) | 6,7(7) | - |
| 6 | 9,8(9) | - | - |
| 7 | 7 | 1-S | 7 |
| 8 | ?,1 | 1a-S | 17,19,20,32(22) |
| 9 | 10 | 1b-c-S | 1 |
| 10 | 1 | 2-S | 1 |
| 11 | 2,1(2) | 2a-d-S | 2 |
| 12 | 2,1(2) | 6-8-S | 1 |
| 13 | ?,1 | - | - |
| 14 | 2,? | 1 | - |
| 15 | 1 | - | - |
Leyenda: El número de setas entre paréntesis indica la media; ? significa que el carácter no pudo ser apreciado.
En la misma descripción, Pulido & Sutil (1981) indican: dorsalmente con 5 pares de pelos lisos, el primer simple y más pequeño... concuerda exactamente con Dyar (1928) y Lane (1953): on dorsal aspect five split hair, the basal one single y dorsal aspect five split hairs, the basal one simple..., respectivamente.
Estas características están presentes en Wy. bicornis de Rancho Grande; vía Ocumare (Edo. Aragua), Sierra de San Luis (Edo. Falcón) y del Páramo Batallón y La Negra (Edo. Táchira) esta última localidad en la misma vertiente -suroriental de la cordillera andina- de la localidad tipo de Wy. trujilloi.
Igualmente, en la larva se puede observar la ramificación presente en las setas 4,5,6,7-C las cuales son múltiples (7-9) en Nunezia (Wy. bicornis, Wy. trujilloi, Wy. (Nuz.) sp. 1, 2, y 3 (Fig. 1a), y por otro lado simples o en pares para los subgéneros Wyeomyia, Exallomyia Harbach & Peyton, Caenomyiella Harbach & Peyton, Zinzala Zavortink y Decamyia Dyar, de acuerdo con especimenes observados en la colección LBV-MB-UCV-IX y a las descripciones de estos subgéneros por Harbach & Peyton (1990,1991).
Fig. 1. Cabeza (a) y segmento VIII (b) de la larva de 4to estadio de Wy. bicornis.
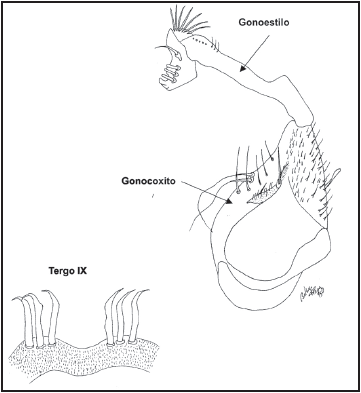
Adicionalmente, la descripción y dibujo de Pulido & Sutil (1981) para los genitales concuerda paso a paso con la descripción proporcionada por Dyar (1928) y Lane (1953) para Wy. bicornis, lo cual puede ser verificado en los holotipos de Wy. bicornis y en Wy. trujillloi (Fig. 2 y 3, respectivamente). La hembra de Wy. trujillloi no fue colectada ni obtenida en cría asociada por Sutil & Pulido (1981), por lo que esta fase no pudo ser comparada con los topotipos, ni con la descripción de Wy. bicornis. No obstante, esta fase es la de menor importancia en la diagnosis específica en comparación con la larva y los genitales del macho.
Fig. 2. Genitales masculinos del holotipo de Wy. bicornis: a) Gonoestilo (200x), b) gonocoxito (100x), c) tergo IX (400x) y d) proctiger (400x).
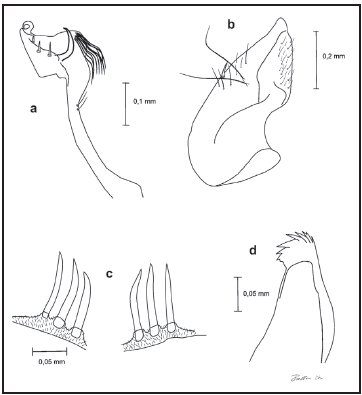
Fig. 3. Genitales masculinos de Wy. trujilloi (tomado de Pulido & Sutil, 1981).
La importante utilización de la morfología de las piezas bucales larvales, en el diagnóstico genérico de la Tribu, igualmente proporciona características diagnósticas para el subgénero Nunezia. El cuerpo maxilar (MxBo), presenta una relación largo/ancho de aproximadamente 1.3x para Nunezia, mientrasque en Wyeomyia (Wyeomyia) grayii Theobald 1901 esta relación es igual a 1; la seta 4-Mx es tan larga como el MxB y ramificada apicalmente, mientras que en Wyeomyia es claramente sencilla. La seta 6-Mx es delgada y corta en Nunezia y cerca de 0.5x el largo del palpo maxilar (MpLp), por el contrario Wy. grayii es gruesa y 0.7x el largo de MpLp (Fig. 4c).
Fig. 4. Maxila de Wy. bicornis (a), detalle de la seta Mx-4 (b), y maxila de Wy. grayii (c) tomado de Harbach & Peyton (1993).
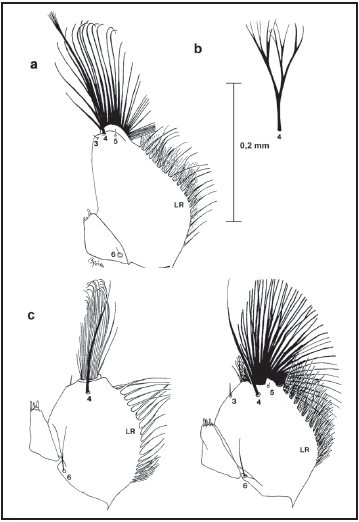
Debido a que la muda de la larva del holotipo de Wy. trujilloi presenta la cápsula cefálica completa (sin disección de las piezas bucales) fue necesario observar estas características en el ejemplar completo. En la Fig. 5 se observa la forma de la maxila en el holotipo de Wy. trujilloi, la cual al ser comparada con la Fig. 4a de Wy. bicornis y la Fig. 4c de Wy. grayii, tipo del subgénero Wyeomyia (mostrada por Harbach & Peyton 1993), se comprueba que la morfología corresponde con la del subgénero Nunezia.
Fig. 5. Maxila de la larva de 4to estadio de Wy. trujilloi (=Wy. bicornis).
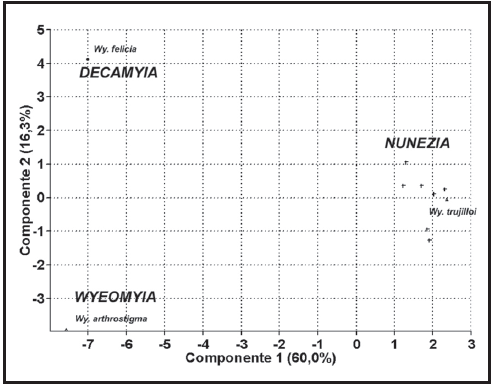
Análisis de Ordenamiento
El ACP (Fig. 6) arrojó una diferenciación entre las especies de los subgéneros Wyeomyia y Decamyia de las especies de Nunezia, incluyendo en este grupo a Wy. trujilloi. El componente principal número uno representado primordialmente por las setas 4-7-C, la 1-S y la relación ancho/largo del sifón explicó el 60% del total de la varianza. Este resultado indica claramente la ubicación subgenérica de Wy. trujilloi en Nunezia y no en Wyeomyia.
Fig. 6. Análisis de Componentes principales en tres subgéneros de Wyeomyia.
Análisis Filogenético
Los análisis realizados con Parsimonia Máxima (Fig. 7) muestran dos topologías o soluciones más parsimoniosas (7a y 7b), de las once encontradas. En las cuales se observa, que el clado X incluye dos subclados (X1 y X2). En el primero (X1) se ubican los individuos de Wy. (Nuz.) sp de Auyantepui y Gran Sabana, pudiendo representar ambos una entidad taxonómica no descrita y el clado X2 con dos subramas que incluyen a Wy. bicornis.
Fig. 7. (a,b) Dos de los 11 árboles más parsimoniosos, y (c) cladograma de consenso para las muestras de Wy. (Nuz.) sp. (círculos negros indican apomorfías y rectángulos blancos homoplasias).
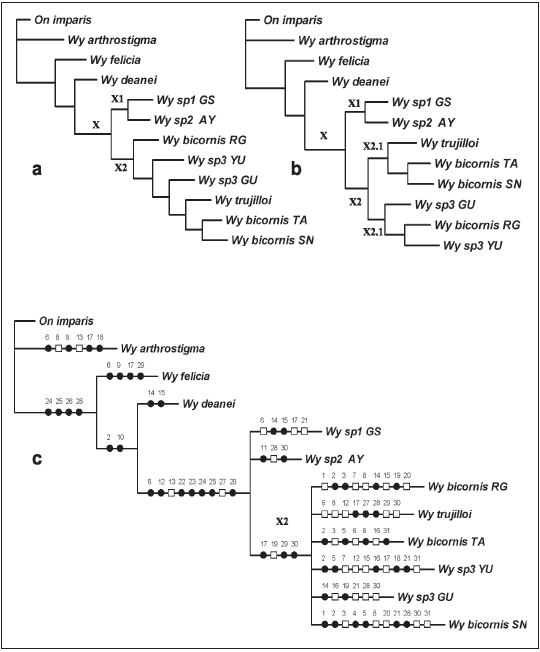
En la Fig. 7b, la subrama X2.1 incluye los individuos pertenecientes Wy. bicornis de los Andes (Tama y Sierra Nevada) incluyendo a Wy. trujilloi, mientras que la otra subrama X2.2 incluye a los individuos de Guayana (Guanay y Yutaje) y la localidad tipo (Wy. bicornis s.s., Rancho Grande).
El cladograma que muestra el consenso estricto (7c), colapsa ambas subramas en una politomía, sugiriendo así que el clado X2 muestra los individuos de Wy. bicornis y sus variedades geográficas, mientras que el clado X1 a otra especie de Nunezia no descrita hasta ahora.
También se aprecian las relaciones entre el subgénero Nunezia, y Wy. (Wyeomyia) arthrostigma, Wy. (Decamyia) felicia, Wy. (Phoniomyia) deanei y Onirion imparis. El género Onirion fue utilizado como taxa de enraizamiento, debido al arreglo mostrado por Harbach & Peyton (2000) en donde este género es grupo hermano del clado (Isostomyia+Shanoniana) + (Phoniomyia (Wyeomyia (Limatus+Sabethes)))). Luego Wy. (Phoniomyia) deanei es grupo hermano de Nunezia, tal como se aprecia en uno de los cladogramas más parsimoniosos presentados por Judd (1996; 1998).
Bionomía
Similarmente, la data bionómica suministrada por Pulido & Sutil (1981), no corresponde con las características ecológicas de las especies de Nunezia ni de Wyeomyia. (Lane & Cerqueira, 1942; Lane, 1943; Lane, 1945; Lane, 1953).
Estos autores señalan haber colectado esta especie en plantas del género Heliconia sp., y asociada a otras especies de Wyeomyia como Wy. felicia y Wy. ulocoma (ambas del subgénero Decamyia). De acuerdo a estudios realizados en la taxonomía y bioecología de mosquitos en Parques Nacionales de Venezuela (Navarro et al., 1995; 2007), el subgénero Nunezia se ha obtenido asociada estrictamente en especies de bromeliáceas, mientras que las especies del subgénero Wyeomyia se han colectado en bromeliáceas, aráceas e internudos de bambú, pero nunca en Heliconiaceas en donde se encuentran exclusivamente especies de Decamyia. Esto podría evidenciar que los autores, mezclaron muestras de diferente origen (bromelias y heliconias) y confundieron ejemplares de las tres especies de Wyeomyia.
Sobre la base de los diferentes análisis descritos, el nombre válido para Wy. (Wy.) trujilloi es Wy. (Nunezia) bicornis. Por consiguiente, realizamos a continuación el tratamiento taxonómico de Wy. bicornis y por último se muestra una descripción parcial detallada de la larva (cabeza y piezas bucales) así como la primera descripción de la pupa.
TRATAMIENTO TAXONÓMICO
Género Wyeomyia Theobald Subgénero Nunezia Dyar, Ortotipo: Dendromyia (Eunicemyia) bicornis Root. Especie tipo: Wyeomyia bicornis (Root, 1928). Venezuela, Brasil, Bolivia y Perú, Incluye especies: Wy. lateralis Petrocchi 1927, Argentina.
Wyeomyia (Nunezia) bicornis (Root, 1928)
bicornis Root, 1928. En: Dyar, 1928: 50 (Dendromyia), holotipo: macho y larva. Localidad tipo: Ocumare, Aragua, Venezuela (NMNH).
Wyeomyia (Nunezia) bicornis de Lane, 1953: 931 (Macho, hembra y larva) Figs. 912 y 913.
Wyeomyia (Nunezia) bicornis de Knight y Stone, 1977: 332 (información del holotipo y distribución geográfica en América).
Wyeomyia (Nunezia) bicornis de Sutil, 1980: 19 (distribución geográfica en Venezuela).
Wyeomyia (Wyeomyia) trujilloi Pulido & Sutil 1981: 219. NUEVA SINONIMIA, localidad tipo: Altamira, Municipio Altamira, Barinas, Venezuela. Holotipo: GM, Le, Pe, (DERM)
Larva (Fig. 1a,b y 5): Se presenta parte de los caracteres y posiciones; el número de ramas se describen en la Tabla I. Maxila (Fig. 5): 1-Mx ausente, setas 2-6-Mx simples (4-Mx larga, gruesa y ramificada en el ápice), LR1 con 18 setas conspicuas, MxB con 22 filamentos. Mandíbula con cuatro dientes ventrales el primero de mayor tamaño que el resto, MnS1 y MnS2 múltiples, MnR con cuatro filamentos, MSA hacia el borde distal. Cabeza: (Fig. 1a) Setas 1,8,10,13,15-C simples, 11-12-C simples o dobles, 14-C doble, 4-C de ocho o diez ramas, 5-C con seis o nueve ramas, 6-C con ocho o nueve ramas, 7-C con siete ramas. Abdomen. 1-VIII con siete u ocho ramas, 2,4-VIII simples, 3-VIII con siete o nueve ramas, 5- VIII con seis o siete ramas, CS con tres hileras de escamas. Sifón: Pseudopeine variable, con 20 a 32 setas. 1-S con siete ramas, 1b,c,d-S simples, 2-S simple, 2a,b,c,d-S dobles, 7-9-S simples. Segmento X: 1-X triple o cuádruple; 2-X con siete u ocho ramas, 3-X doble; 4-X con siete u ocho ramas.
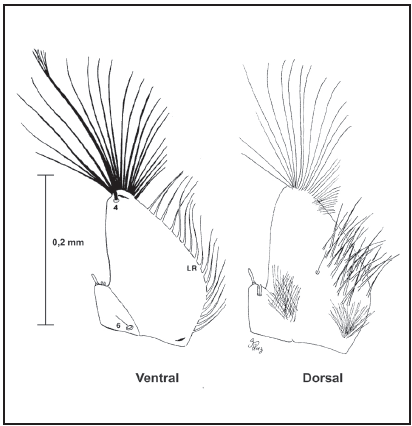
Pupa, redescripción (Fig. 8): Se muestran las posiciones y números de las setas (Tabla II). Cefalotórax: Seta 1-CT doble y larga; 2-7-CT y 9- CT con aproximadamente 1/6 del largo de 1-CT; 2- 3,9-CT dobles, 5,7-CT triples, 6-CT cuádruple; 8-CT doble y la mitad de larga que 1-CT; 10-12-CT simples y largas. Trompeta: Larga y cilíndrica. Abdomen: Seta 1-I con diez ramas tipo dendrítico, 1-II doble o con seis ramas, 1-III con seis ramas, 1-IV-V con ocho ramas, 1-VI-VIII dobles; 2-I-VI simples; 3-I-III,IVVIII simples, 3-IV con cuádruple, 3-V simple o doble; 4-1 con cinco o seis ramas, 4-II cuádruples, 4-III-IV simples, 4-VI triple, 4-VII-VIII doble; 5-I-II simples, 5-III doble, 5-IV-VI triples y largas, 5-VIII simple o triple y larga; 6-I-II simple y larga, 6-III simples y corta, 6-IV doble o triple, 6-V cuádruple o quíntupla, 6-VI simple o doble, 6-VII doble; 7-I con cuatro a seis ramas, 7-III doble o cuádruple, 7-IV-V simples o dobles, 7-VI doble, 7-VII simple; 8-II-III triples o cuádruples, 8-IV-V cuádruples, 8-VI con tres o cuatro ramas, 8-VII cuádruple o quíntupla; 9-I-VI simples, 9-VII con quince ramas tipo dendrítico, 9-VIII de veintisiete o treinta ramas tipo dendrítico; 10-III,V dobles, 10-IV,VI simples o dobles 10-VIII simple; 11- II triple y larga, 11-III simple o triple, 11-IV doble o triple, 11-V simple o doble, 11-VI doble o cuádruple, 11-VII simple o doble; 14-VIII simple. Paleta: Con una hilera de espículas en el borde interior o distal.
Fig. 8. Quetotaxia de la pupa de Wy. bicornis.
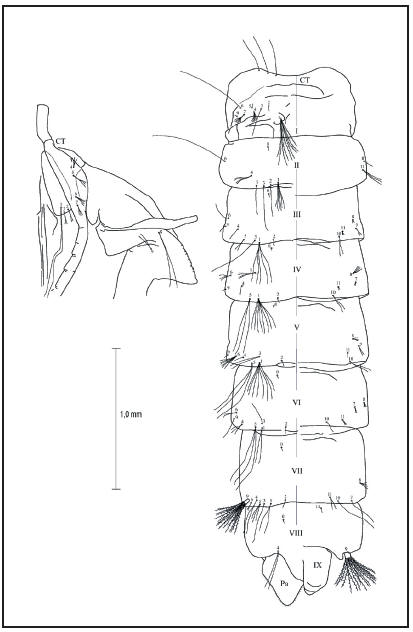
Tabla II. Quetotaxia de la pupa de Wyeomyia (Nunezia) bicornis, (n=2).
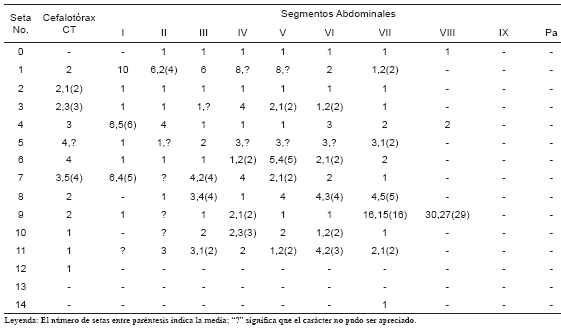
Leyenda: El número de setas entre paréntesis indica la media; ? significa que el carácter no pudo ser apreciado.
AGRADECIMIENTOS
Los autores desean agradecer su valiosa colaboración a las siguientes personas e instituciones: Insp. Jesús Amarista () e Ing. Carmen Escalante de Ugueto por facilitarnos el holotipo de Wy. trujilloi de la colección DERM; al Dr. Richard Wilkerson y Sr. Thomas Gaffigan, por el préstamo del holotipo de Wy. bicornis de la colección NMNH, MSc. Belkys Pérez por el trabajo de disección, montaje en laboratorio (partes bucales) y dibujos de piezas bucales; a Adriana Zorrilla por la búsqueda de los datos de colección; Lic. Norberto Rebolledo, Director del Centro de Ecología de las Tierras Altas, El Cobre, Edo. Táchira (CETA-PN Juan Pablo Peñaloza); los Doctores Armando Michelangeli y Ricardo Guerrero, Fundación Terramar (Expediciones Yutaje-Guanay y Auyantepui) e INPARQUES; al Dr. Luis Daniel Otero y Lic. Jorge Avila (ULA-Mérida). Al CDCH – UC y la Facyt (Decana Dra. Yaqueline Loyo de Sardi), por facilitar los viáticos y pasaje (JL) para la visita a NMNH. Este trabajo fue financiado por CONICIT-RP-VII-240060 (JCN) y FONACIT LAB-2000001593, Grupo CNRA (J Clavijo y JCN).
REFERENCIAS
1. Belkin J. N. (1962). The Mosquitoes of the South Pacific (Diptera: Culicidae). Vol. 1 y 2. University of California Press, Los Angeles, USA. [ Links ]
2. Dyar H. G. (1928). The Mosquitoes of the Americas. Carnegie Institution of Washington, USA. [ Links ]
3. Fish D. (1983). Phytotelmata: Flora and Fauna. pp 1-25. En: Phytotelmata: Terrestrial plants as host aquatic insect communities. Eds. Frank J. H. & Lounibos L. P. Plexus publishing, New Jersey, USA. [ Links ]
4. Guimarães J. H. (1997). Systematic Database of Diptera of the Americas South of the United States, Family Culicidae. Ed. Pleide. Sâo Paulo: Fundação de Amparo a Pesquisa do Estado de Sâo Paulo, Brasil. [ Links ]
5. Goloboff P. A. (1996). Nona. Noname ver. 1,50 (versión 32-bit). Programa y manual distribuido por J. M. Carpenter. Dept. Entomology, American Museum of Natural History, New York, USA. [ Links ] 6. Hammer O. & Harper D. A. T. (2004). PAST: Palaeontological Statistics, version 1.30. Disponible en URL: http://folk.uio.no/ohammer/past. [ Links ]
7. Harbach R. E. (1977). Comparative and functional morphology of the mandibles of some fourth stage mosquito larvae (Diptera: Culicidae). Zoomorphologie. 87: 217-236. [ Links ]
8. Harbach R. E. (1991a). A new Subgenus of the Genus Sabethes (Diptera: Culicidae). Mosq. Syst. 23: 1-9. [ Links ]
9. Harbach R. E. (1991b). Neotype designation, Generic realignment and description of Dendromyia shnusei Martini (Diptera: Culicidae). Mosq. Syst. 23: 175-181. [ Links ]
10. Harbach R. E. (1994). The Subgenus Sabethinus of Sabethes (Diptera: Culicidae). Syst. Entomol. 39: 207-234. [ Links ]
11. Harbach R. E. (1995). A new Sabethes of the Subgenus Peytonulus (Diptera: Culicidae) with an unusual fourth-instar larva. Ent. Scand. 26: 87-96. [ Links ]
12. Harbach R. E. & Knight K. L. (1977). A Mosquito Taxonomy Glossary X. The Larval Mandible. Mosq. Syst. 9:25-27 [ Links ]
13. Harbach R. E. & Knight K. L. (1980). Taxonomists Glossary of Mosquito Anatomy. Plexus Publishing, Inc., Marlton, New Jersey, USA. [ Links ] 14. Harbach R. E. & Navarro J. C. (1996). A new species of Anopheles subgenus Kerteszia (Diptera: Culicidae) from Venezuela. Ent. Scand. 27: 207-216. [ Links ]
15. Harbach R. E. & Petersen J. L. (1992). Two species previously confused under the concept of Sabethes tarsopus in Central America (Diptera: Culicidae). Mosq. Syst. 24: 102-124. [ Links ]
16. Harbach R. E. & Peyton E. L. (1990). A new Subgenus in Wyeomyia (Diptera: Culicidae) with the reclassification and redescription of the type species, Sabethes fernandezyepezi. Mosq. Svst. 22: 15-23. [ Links ]
17. Harbach R. E. & Peyton E. L. (1991). A new Subgenus in Wyeomyia (Diptera: Culicidae), with the reclassification and redescription of Wyeomyia (Davismyia) arborea, Wyeomyia (Dendromyia) tarsata and Sabethes (Sabethes) carrilloi. Mosq. Svst. 23: 92-109. [ Links ]
18. Harbach R. E. & Peyton E. L. (1993). Morphology and evolution of the larval maxila and its importance in classification of the Sabethini (Diptera: Culicidae). Mosq. Syst. 25: 1-16. [ Links ]
19. Harbach R. E. & Peyton E. L. (2000). Systematics of Onirion, a new genus of Sabethini (Diptera: Culicidae) from the Neotropical Region. Bull. Mus. Nat. Hist. Lond (ent). 69: 115-169. [ Links ]
20. Judd D. (1996). Review of the systematics and phylogenetic relationships of the Sabethini (Diptera: Culicidae). Syst. Entomol. 21: 129-150. [ Links ]
21. Judd D. (1998). Review of a bromeliad-ovipositing lineage in Wyeomyia and the resurrection of Hystatomyia (Diptera: Culicidae). Ann. Entomol. Soc. Am. 91: 572-589. [ Links ]
22. Kitching I., Forey P., Humphries C. & Williams D. (1998). Cladistics: The Theory and Practice of the Parsimony Analysis. Oxford University Press. [ Links ]
23. Knight K. L. & Stone A. (1977). A Catalog of the Mosquitoes of the World (Diptera: Culicidae). San Thomas Foundation, Entomological Society of America, Vol. VI. USA. [ Links ]
24. Lane J. (1943). The Geographical distribution of Sabethini. Rev. Ent. 14: 409-429. [ Links ]
25. Lane J. (1945). Os sabetíneos da América (Addenda e Corrigenda). Rev. Ent. 16: 132-157. [ Links ]
26. Lane J. (1953). Neotropical Culicidae. Vol. II. Published by the University of São Paulo, Brasil. [ Links ]
27. Lane, J. & Cerqueira N. L. (1942). Os sabetíneos da América (Díptera: Culicidae). Arq. Zool. São Paulo. 3: 473-849. [ Links ]
28. Liria J. & Navarro J. C. (1999). Morfología bucal larval de Deinocerites melanophylum Dyar & Knab y D. cancer Theobald (Diptera: Culicidae). Bol. Entomol. Venezolana. 14: 53-62. [ Links ]
29. Motta M. I. & Lourenço de Oliveira R. (2005). Spilonympha, a new subgenus of Wyeomyia (Diptera: Culicidae) and description of a new species Wyeomyia aninge. Ann. Entomol. Soc. Am. 98: 832-852. [ Links ]
30. Navarro J. C., Ingunza J., Fernández Z. & Barrera R. (1995). Mosquitoes and bromeliads: speciesspecific selectivity patterns on the northern coast and southern Guayana Shield in Venezuela. J. Am. Mosq. Control Assoc. 11: 345-346. [ Links ]
31. Navarro J. C., Liria J., Piñango H. & Barrera R. (2007). Biogeographic area relationships in Venezuela: A parsimony analysis of Culicidae- Phytotelmata relationships distributions in national Parks. Zootaxa. 500: 1500 (en prensa). [ Links ]
32. Nixon K. C. (1999). Winclada (BETA) ver. 0.9.99m24 publicado por el autor, Ithaca, NY. Disponible en URL: http://www.cladistics.com. [ Links ]
33. Pérez B. Y. & Navarro J. C. (1996). Morfología bucal de larvas de mosquitos: herramienta taxonómica a nivel subgenérico Anopheles (Diptera: Culicidae). Acta Zool. Mex. 68: 13-26. [ Links ]
34. Porter, C. H. & Wolf M. I. (2004). A new species of Wyeomyia (Hystatomyia) (Diptera: Culicidae) from Colombia and a redescription of Wy. intonica Dyar & Knab. Zootaxa. 477: 1-31. [ Links ]
35. Pulido J. & Sutil E. O. (1981). Wyeomyia (Wyeomyia) trujilloi (Diptera, Culicidae) nueva especie de Venezuela. Bol. Direcc. Marariol. San. Amb. 21: 219-226. [ Links ]
36. Rae T. C. (1998). The Logical Basis for the use of Continuous Characters in Phylogenetic Systematics. Cladistics. 14: 221-228 [ Links ]
37. Reinert J. F. (2001). Revised list of abbreviations for genera and subgenera of Culicidae (Diptera) and notes on generic and subgeneric changes. J. Am. Mosq. Control Assoc. 17: 51-55. [ Links ]
38. Sutil O. E. (1980). Enumeración histórica y geográfica de las especies de Culicidae de Venezuela ordenadas según su taxonomía. Bol. Dir. Malariol. San. Amb. 20: 1-32. [ Links ]
39. Thiele K. (1993). The Holy Grail of the perfect character: the cladistic treatment of morphometric data. Cladistics. 9: 275–304. [ Links ]
40. Varga L. (1928). Ein interenssanter biotop der bioconosse von Wasser-organismen. Biologisches Zentralbtatt. 48: 143-162. [ Links ]
41. Wirth W. W. (1961). Instructions for preparing studies of Ceratopogonidae and Chironomidae. Studia Ent. 4: 553-554. [ Links ]
42. Zavortink T. J. (1985). Zinzala, a new Subgenus of Wyeomyia with two new species from, pitcherplants in Venezuela (Diptera, Culicidae, Sabethini). The Wasman Jourmal of Biology. 43: 46-59. [ Links ]
Apendice I.- Matriz de datos merísticos y morfométricos para quetotaxia larval.
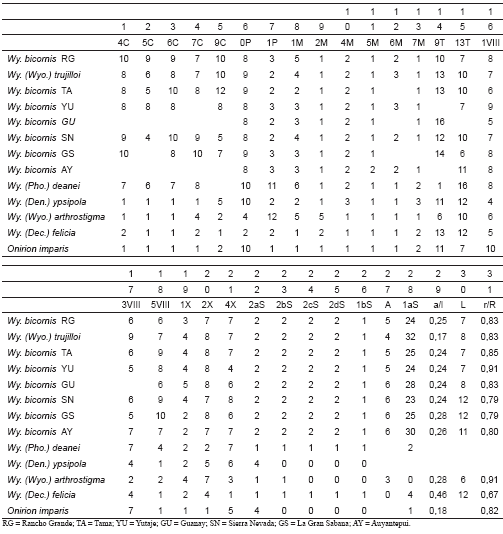
Apendice II. Matriz de datos merísticos y morfométricos para quetotaxia larval, con codificación
Gap-Weighting.